
Abstract
Infections caused by metallo-β-lactamase (MBL)-producing bacteria are emerging and carry a significant impact on patients' outcome. MBL producers are spread worldwide, both in community and hospital setting, with increasingly reported epidemic clusters and the search for MBL inhibitors is an important topic for public health. MBLs are zinc-dependent enzymes whose functioning can be hampered by zinc chelators. We evaluated the potential of six zinc chelators (disulfiram, nitroxoline, 5-amino-8-hydroxyquinoline, 1,4,7,10-tetraazacyclododecane-1,4,7,10-tetraacetic acid [DOTA], cyclam, and N,N,N′,N′-tetrakis (2-pyridymethyl) ethylenediamine [TPEN]) in restoring carbapenem activity against MBL producers. Zinc chelators alone or in combination with meropenem against MBL-producing Klebsiella pneumoniae, Chryseobacterium indologenes, Elizabethkingia meningoseptica, and Stenotrophomonas maltophilia isolates were tested in vitro and in vivo (Galleria mellonella). In vitro experiments showed a synergistic activity between TPEN and meropenem toward all the strains. Nitroxoline alone retained activity against S. maltophilia, C. indologenes, and E. meningoseptica. In vivo experiments showed that TPEN or nitroxoline in combination with meropenem increased survival in larvae infected with E. meningoseptica, S. maltophilia, and K. pneumoniae. Based on our data, zinc chelators are potential carbapenem adjuvants molecules (restoring carbapenem activity) against MBL-sustained infections and could represent an interesting option for infections induced by these microorganisms.
Introduction
Infections caused by metallo-β-lactamase (MBL)-producing Gram-negative bacteria have limited therapeutic options, and an unfavorable impact on patients' outcome. 1
MBLs are diffused worldwide in several bacterial species and MBL producers are responsible for both community- and hospital-acquired infections, and in epidemic forms. The increase in population exchange at global level and enhanced medical tourism could play a significant role in spreading uncontrolled MBL-related resistance worldwide. Moreover, the prevalent plasmid-mediated nature and intestinal carriage of MBL producers strongly promote their spread. 2
The main MBLs are Verona integron-encoded MBL (VIM), imipenemase (IMP), and New Delhi MBL (NDM) types. The Indian subcontinent, the Balkan region, and the Middle East are the main endemic areas for NDM enzymes, 3 but this determinant is also emerging in Africa, representing an alarming cause of concern for European countries, due to considerable migration flows. 4 Notably, a large outbreak of NDM-producing Enterobacteriaceae involving seven hospitals and 350 patients in Tuscany region has been recently described in Italy. 5
MBL producers are usually nonsusceptible to all currently available β-lactams (including carbapenems and the recently approved β-lactam/β-lactamase inhibitor combinations ceftolozane/tazobactam, ceftazidime/avibactam, and meropenem/vaborbactam), with the exception of monobactams. 6 Owing to the frequent co-presence of other resistance mechanisms, acquired-MBL producers are usually also resistant to aminoglycosides, fluoroquinolones, and sulfonamides, thereby considerably hampering their effective treatment due to the paucity of dependable therapeutic options. 7
Acquired MBLs are commonly encountered in Enterobacteriaceae, Pseudomonas aeruginosa, and Acinetobacter baumannii. 6 Other bacterial species can intrinsically produce MBLs. Of note, some nonfermentative Gram-negative bacilli naturally producing MBLs (Chryseobacterium indologenes, Elizabethkingia meningoseptica, and Stenotrophomonas maltophilia) are also intrinsically resistant or less susceptible to several antibiotics, including colistin.8,9 There has been an increased incidence of infections caused by these opportunistic bacteria in critical care settings during the last 5 years.10–12
In recent years, interest in the use of zinc chelators as MBL inhibitors has been growing.13,14 In fact, since MBL enzymes require one or two zinc ions for their activity, zinc chelators could act as zinc “strippers” from MBL enzymes, depriving them of anti-β-lactam activity, or as metal-binding competitive MBL inhibitor. 15 In 2014, an MBL inhibitor extracted from Aspergillus versicolor, called aspergillomarasmine A was identified. This compound acts with a metal depletion mechanism and has a selectivity toward NDM and VIM MBLs, and was clearly demonstrated to restore meropenem activity against NDM-producing Enterobacteriaceae both in vitro and in vivo. 13 Similarly, in 2018, compounds based on the selective zinc chelator tris-picolylamine were developed and demonstrated the ability to restore the bactericidal activity of meropenem against VIM-producing P. aeruginosa and NDM-producing K. pmeumoniae. 14
Consistent with this hypothesis, in vitro synergistic activity of 1,4,7,10-tetraazacyclododecane-1,4,7,10-tetraacetic acid (DOTA), dithiocarbamates, N,N,N′,N′-tetrakis (2-pyridymethyl) ethylenediamine (TPEN), and dipicolinic acid with meropenem was demonstrated against NDM and VIM producers.16–18 However, there are only a few studies in animal models that investigate the potential therapeutic applications of zinc chelators in infectious diseases.18,19
The larvae of the greater wax moth Galleria mellonella are a reproducible and advantageous alternative in vivo system to mammalian models (e.g., mice). The immune system of insects shares many similarities with the innate immune response of mammals and as a consequence, insects may be used to assess the virulence of microbial pathogens and produce results comparable to those obtained using mammalian systems.20,21 Larvae have been used to assess the virulence and pathogenesis of multidrug-resistant bacterial strains and have also been employed to test the toxicity of novel antimicrobial agents (e.g., phenanthroline chelators),22–24 the efficacy of novel antimicrobial agents, 25 the repurposing of antimicrobial agents, 26 and also in the development of new drug combination therapies. 27
The search for MBL inhibitors is now an important topic for public health.27,28 Although chelators were largely used in microbiology laboratories for diagnostic purposes and in chelation therapy,27–29 they have never been included in anti-MBL commercially available pharmacologic formulations. It is likely that two aspects have contributed to the fact that zinc chelators have not been used so far: (1) MBL producer infections are a relatively recent clinical problem and (2) the development of a single inhibitor to neutralize MBLs has been deemed too technically challenging, and overcoming in vivo toxicity associated with cross reactivity with human metalloenzymes has been a concern. 13 Against this background, provided their activity and efficacy (plus tolerability) are confirmed in preclinical and clinical studies, therapies including MBL inhibitors may become a reasonable alternative to classical therapeutic approaches to infections due to MBL producers, especially when robust safety data will become available.
In this study, we assessed the in vitro and in vivo activity of six zinc chelators against MBL producers, either alone or in combination with meropenem, to evaluate their potential in restoring meropenem activity against difficult-to-treat MBL-producing pathogens. In particular, we tested six known ligands (L): disulfiram 30 (anti-alcohol drug); nitroxoline (HQ5NO2) 31 (antibiotic); 5-amino-8-hydroxyquinoline (HQ5NH2) 31 ; DOTA 32 ; cyclam (1,4,8,11-tetraazacyclotetradecane), 33 and TPEN 34 (a membrane-permeable chelating agent widely used in zinc biology) (Fig. 1). The ligands form Zn complexes with different stability (Table 1).
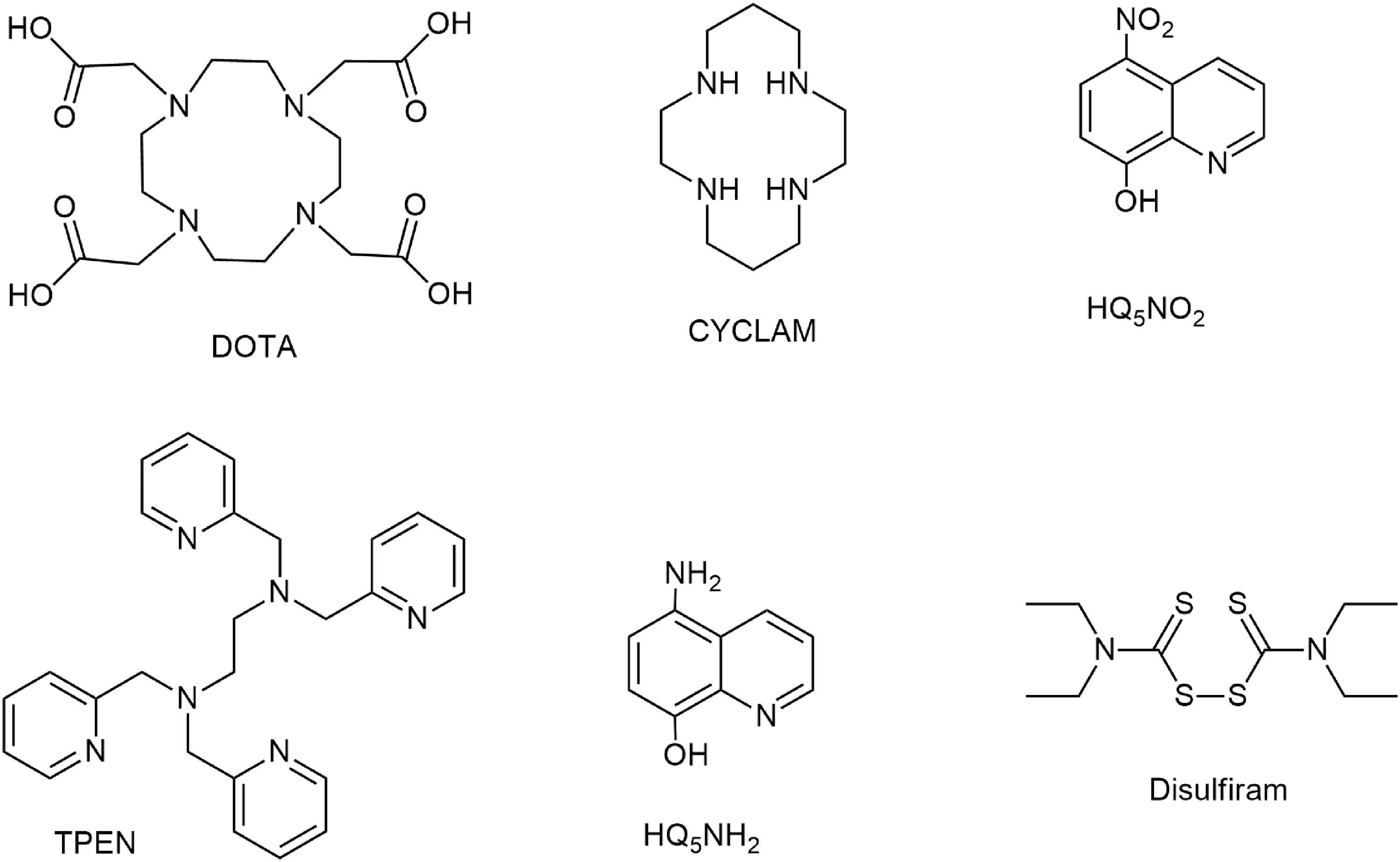
Chemical structure of Zn chelators used in this study.
Stability Constants of Zinc(II) Complexes with L
Zn2+ + Ln ⇆ ZnL K1; ZnL(2−n) + Ln ⇆ ZnL2(2−2n) K2
DOTA, 1,4,7,10-tetraazacyclododecane-1,4,7,10-tetraacetic acid; TPEN, N,N,N′,N′-tetrakis (2-pyridymethyl) ethylenediamine.
Materials and Methods
Chemicals
All chemicals obtained from commercial sources were used without further purification. Nitroxoline (HQ5NO2, MW 190.16 g/mol) and 5-amino-8-hydroxyquinoline (HQ5NH2, MW 160.17 g/mol) were purchased from Sigma Aldrich (Milan, Italy). TPEN (MW 424.54 g/mol), DOTA (MW 404.2 g/mol), disulfiram (MW 296.54 g/mol), and cyclam (MW 200.33 g/mol) were purchased from TCI (Tokyo, Japan). The purity of the ligands was ≥98%. NMR spectra were recorded for all the ligands used.
Stock solutions of chelators (5 g/L) (HQ5NO2 26.29 mM; HQ5NH2 31.21 mM; TPEN 11.78 mM; DOTA 13.37 mM; disulfiram 16.89 mM; and cyclam 24.96 mM) were prepared in water (DOTA, cyclam, TPEN, and disulfiram) or in dimethyl sulfoxide (DMSO) (nitroxoline and 5-amino-8-hydroxyquinoline) for in vitro experiments.
Bacterial isolates
We studied four MBL-producing clinical isolates responsible for invasive infections in patients from intensive care unit (ICU) of “Alessandro Manzoni” Hospital (Lecco, Italy). Bacterial isolates were selected on the basis of their origin (invasive, ICU patients), resistance to carbapenems, typology of carbapenemases (chromosomally encoded or acquired MBLs), and availability of very few therapeutic alternatives. Bacterial isolates were as follows: Klebsiella pneumoniae (LC954/14), S. maltophilia (LC669/17), C. indologenes (LC650/17), and E. meningoseptica (LC596/11). Bacterial identification was performed by MALDI-TOF (Vitek MS, bioMérieux, Marcy l'Etoile, France). K. pneumoniae isolate (LC954/14) has been previously described. 4 It was isolated from blood in 2014, produced an acquired NDM-1 determinant, belonged to ST11, and represented the first described NDM-producing K. pneumoniae imported from Africa to Italy. Remaining isolates produced chromosomally encoded carbapenemases. In particular, S. maltophilia (LC669/17, this study) was isolated from blood in 2017, C. indologenes (LC650/17, this study) was isolated from blood in 2017, and E. meningoseptica (LC696/11, this study) was isolated from bronchoalveolar fluid in 2011.
Separated evaluation of antimicrobial activity of meropenem and zinc chelators
Antibiotic minimum inhibitory concentration (MIC) values were measured by broth microdilution method using a dedicated TREK panel (DKMGN; Thermo Fisher Diagnostics, Milan, Italy). Antimicrobial activity of the six zinc chelators against bacterial isolates was investigated using the Kirby-Bauer method, plating a standard 0.5 McF inoculum for each isolate Mueller Hinton II agar (MHA; bioMérieux). Ten microliter of concentrated solution of each chelator (5 g/L) was placed on blank disks and plated on inoculated MHA. The presence of inhibition ring was further investigated by standard broth microdilution (CLSI) using Cation-Adjusted Mueller-Hinton (CAMH; Thermo Fisher Diagnostics) broth to asses MIC values.
Evaluation of interactions between meropenem and zinc chelators
Interactions were determined by preliminary qualitative double-disk diffusion test, followed by quantitative checkerboard and time-kill assays. All tests were performed in triplicate.
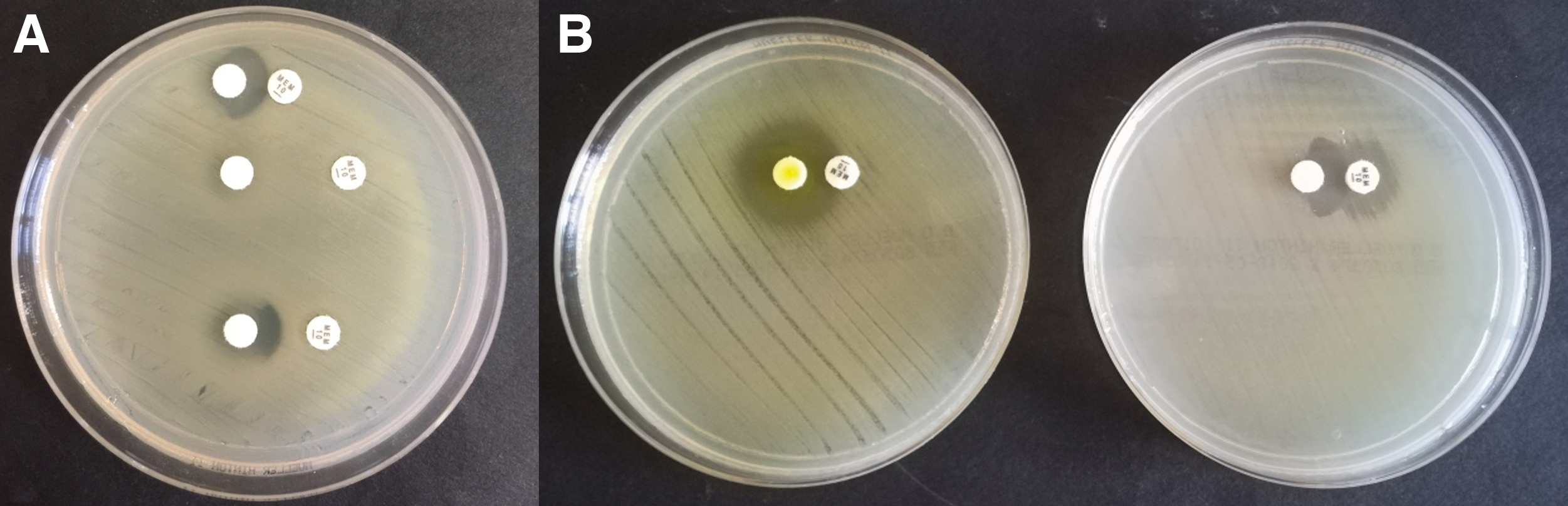
Double-disk diffusion tests were performed with disks placed at 10, 15, and 20 mm center-to-center for each combination chelator-meropenem (Fig. 2A). Standard 0.5 McF inoculums for each isolate were plated on MHA. Ten microliters of concentrated solution of each chelator (5 g/L) was placed on blank disks on MHA. Disks of meropenem (10 μg; bioMérieux) have been used. Synergy was defined as the presence of alterations in disk's inhibition rings.
Checkerboard assays
Positive interactions in double-disk diffusion tests were further investigated by the checkerboard assay as previously described. 35 MIC values of chelator and meropenem were measured by standard broth microdilution method (CLSI) using CAMH broth. The range of drug concentration used in the checkerboard analysis was such that the dilution range encompassed the MIC for each drug used in the analysis. Broth microdilution plates containing CAMH broth were inoculated with each bacterial isolate to yield ∼106 CFU/mL in a 100-μL final volume and incubated for 18 hours at 37°C. Synergy was defined as a fourfold reduction in the MIC of both antibiotics in combination, compared with each used alone, measuring the fractional inhibitory concentration index (FICI). The FICI was calculated for each combination using the following formula: FICI = FICA + FICB, where FICA = MIC of drug A in combination/MIC of drug A alone, and FICB = MIC of drug B in combination/MIC of drug B alone. The FICI was interpreted as follows: synergy = FICI ≤0.5; indifference = 0.5 < FICI ≤4; antagonism = FICI >4. 35
Time-kill assays
Time-kill assays were performed in CAMH broth using 0.5 × MIC of antibiotic and chelator, which demonstrated synergistic interactions in checkerboard assays. Bacteria were inoculated in 2.5 mL of broth containing meropenem, chelator, and a combination of them to reach a final concentration of ∼1 × 107 CFU/mL and incubated at 37°C. Viable cell counts were performed after 30 minutes, 1, 2, 4, and 8 hours of exposure (detection limit, 100 CFU/mL), streaking 10-fold dilutions of broth cultures in MHA. Synergy was defined as a >2 log10 decrease of CFU/mL between combination and its most active component. 36
Larval culture and inoculation
Sixth instar larvae of the greater wax-moth G. mellonella (Livefoods Direct Ltd., Sheffield, England), were stored in the dark at 15°C and maintained on wood chippings.21,37 Larvae weighing 0.22 ± 0.03 g were selected and used within 2 weeks of receipt. Ten healthy larvae per treatment and controls were placed in sterile 9-cm Petri dishes lined with Whatman filter paper and containing some wood shavings. Larvae were inoculated with drug and/or bacterial pathogen through the last left pro-leg into the hemocoel with a Myjector U-100 insulin syringe (Terumo Europe N.V., Belgium). Larvae were acclimatized to 37°C for 1 hour before all experiments and incubated at 37°C for all studies. All experiments were performed independently on three separate occasions.
Determination of toxicity of test compounds
Larvae (n ≥ 10) were injected with 20 μL of TPEN (phosphate-buffered saline [PBS]), nitroxoline (10% DMSO/PBS), and meropenem (PBS) at a range of concentrations (128, 64, 8, and 2 mg/L) with the addition of 10% DMSO/PBS control and PBS only (20 μL). For assessment of larval viability, the larvae were gently probed with a needle and if no response was observed, the larvae were considered to be dead. Experiments were performed on three independent occasions.
Determination of hemocyte density following injection with test compounds
The larval hemocyte density was determined by injection of TPEN, nitroxoline, and meropenem at varying concentrations (128, 64, 8, and 2 mg/L). Changes in hemocyte density were assessed by bleeding 40 μL each from G. mellonella larvae (n = 5) into a microcentrifuge tube on ice to prevent melanization. Hemolymph was diluted in 0.37% (v/v) mercaptoethanol-supplemented PBS and cell density was determined using a hemocytometer. Hemocyte density was expressed in terms of hemocyte density/larva. Experiments were performed on three independent occasions and the mean ± standard error was determined.
Efficacy of chelators on antibiotic activity in G. mellonella larvae
Bacterial species (E. meningoseptica, S. maltophilia, K. pneumoniae, and C. indologenes) were grown to stationary phase in nutrient broth, centrifuged and washed in PBS thrice, resuspended in PBS, and OD600 adjusted to 0.1. In total, 20 μL of each bacterial suspension was injected into the last left pro-leg and larvae incubated at 37°C for 1 hour. Injection of larvae with C. indologenes (20 μL of OD600 = 1 suspension) did not decrease larval survival due to a lack of virulence; therefore, the in vivo efficacy of chelator/antibiotic combinations was not assessed using C. indologenes. Following this incubation, larvae were also inoculated with 20 μL of a suspension containing meropenem (64 mg/L) alone, TPEN or nitroxoline alone (128, 64, and 8 mg/L), or meropenem with the addition of test compound (TPEN or nitroxoline) at a range of concentrations (128, 64, and 8 mg/L) and viability was assessed over 120 hours. Experiments were performed on three independent occasions.
Results
In vitro results
Evaluation of antibiotic MIC values and specific antimicrobial activity of zinc chelators
Bacterial isolates showed high MIC values for the majority of tested antibiotics (including meropenem) and low MIC values for tigecycline (Table 2). Low MIC values were also observed for trimethoprim/sulfamethoxazole in S. maltophilia, C. indologenes, and E. meningoseptica, for colistin in NDM-producing K. pneumoniae, and for ciprofloxacin in C. indologenes and E. meningoseptica (Table 2).
Minimum Inhibitory Concentration Values of New Delhi Metallo-β-Lactamase-Producing Klebsiella pneumoniae, Stenotrophomonas maltophilia, Chryseobacterium indologenes, and Elizabethkingia meningoseptica, as Assessed by Broth Microdilution Method
MIC values reported for trimethoprim/sulfamethoxazole concern trimethoprim component.
MIC, minimum inhibitory concentration.
Regarding chelators, relevant antimicrobial activity was demonstrated only for nitroxoline against S. maltophilia, C. indologenes, and E. meningoseptica, as testified by the presence of inhibition ring around disks using the Kirby-Bauer method (Table 3 and Fig. 2B). No inhibition zone was observed for NDM-producing K. pneumoniae. TPEN showed weak antimicrobial activity against C. indologenes and E. meningoseptica. Standard broth microdilution showed a high MIC value (256 mg/L) of nitroxoline for NDM-producing K. pneumoniae, confirming the absence of antimicrobial activity against this strain. MIC values for nitroxoline ranging from 1 to 4 mg/L were observed for other strains. An MIC value of 16 mg/L was observed for TPEN against C. indologenes and E. meningoseptica (Table 3).
Chelator-Related Antimicrobial Activity
MIC values are in mg/L.
Synergy tests
Double-disk diffusion tests showed alterations in disk's inhibition rings for combinations of TPEN and meropenem against all tested isolates (Fig. 2B), and a weak alteration for the combination of HQ5NH2 and meropenem against S. maltophilia. No other alteration was observed.
The checkerboard analysis confirmed the presence of synergistic interactions between TPEN and meropenem against all tested strains (Table 4), while it showed indifference between HQ5NH2 and meropenem against S. maltophilia.
Synergistic Concentrations of N,N,N′,N′-Tetrakis (2-Pyridylmethyl) Ethylenediamine and Meropenem Obtained by Checkerboard, Time-Kill, and In Vivo Assays
N/A, not available.
Time-kill assays further confirmed the presence of synergistic interactions between TPEN and meropenem detected in checkerboard assays. The combination of TPEN and meropenem (0.5 × MIC) was synergistic against all tested strains (Table 4). The greatest reduction in cell viability was observed for E. meningoseptica, against which the combination reduced 5 log10 the CFU/mL compared to antibiotic and chelator alone (Fig. 3). No antagonism was detected by all methods.

Time-kill assay.
In vivo study
Determination of toxicity of test compounds on G. mellonella larvae
Inoculation of test compounds resulted in no decrease in G. mellonella larval survival over 72 hours. There was no melanization of the cuticle in response to drug injection and some larvae formed pupae after 2 weeks, indicating test compounds had no adverse effect on larval development. Some antimicrobial agents with no inherent antibacterial activity possess immune stimulatory effects by increasing the number of circulating hemocytes in G. mellonella larvae, which protects against a subsequent bacterial infection. To exclude these effects, changes in hemocyte density were determined after inoculation of test compound at 24 hours. Injection of larvae with meropenem at 8 and 128 mg/L resulted in a slight increase in circulating hemocyte density to 55 ± 1.41 × 105/mL and 51 ± 1.41 × 105/mL, respectively, compared to PBS-injected larvae (36 ± 2.82 × 105/mL); however, this was not statistically significant. The highest concentration of TPEN and nitroxoline resulted in a nonsignificant increase in hemocyte density to 58.33 ± 6.02 × 105/mL and 60.66 ± 7.02 × 105/mL relative to the control larvae (36 ± 2.82 × 105/mL). There was no increase in hemocyte density relative to the control after inoculation with DMSO (10% v/v PBS).
Analysis of in vivo efficacy of chelators alone and of meropenem supplemented with chelators
To determine if TPEN and nitroxoline displayed intrinsic antimicrobial activity in vivo, larvae were infected with E. meningoseptica, S. maltophilia, and K. pneumoniae, and followed up with TPEN and nitroxoline as described. Larval survival over 120 hours was assessed. Treatment of infected (E. meningoseptica, S. maltophilia, and K. pneumoniae) larvae with TPEN (128, 64, and 8 mg/L) displayed no enhanced survival relative to the PBS control. Interestingly, nitroxoline only at a concentration of 128 mg/L displayed in vivo efficacy at enhancing G. mellonella larval survival at 48 hours (40% ± 6.33%) and 72 hours (28% ± 9%) with E. meningoseptica, 96 hours (48% ± 6.33%) with S. maltophilia, and 24 hours (30% ± 9%) with K. pneumoniae relative to the PBS-injected controls (Fig. 4).
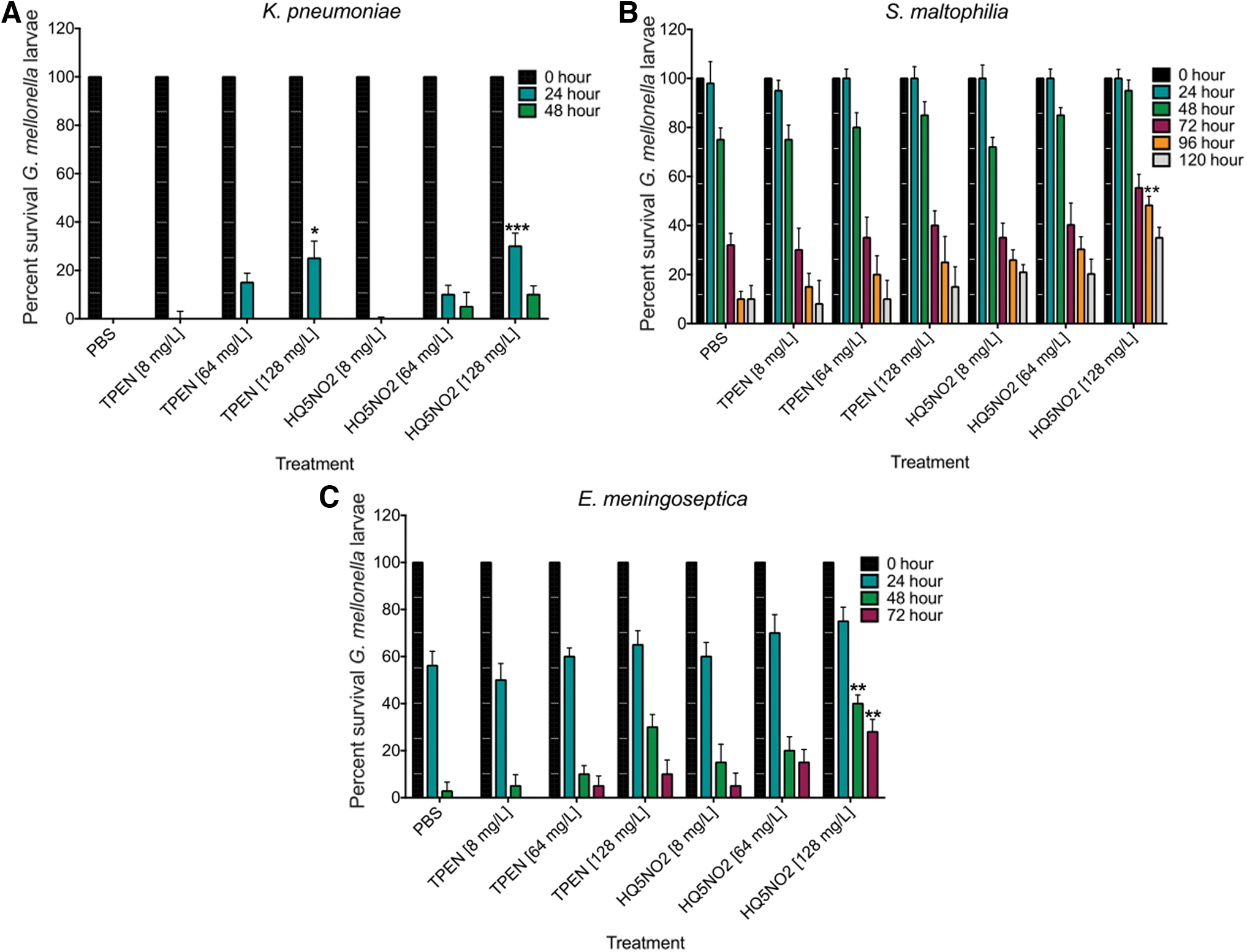
Effect of TPEN and nitroxoline on the survival of Galleria mellonella larvae. Effect of TPEN and nitroxoline alone on the survival of G. mellonella larvae (n ≥ 10). Larvae infected with 20 μL of Elizabethkingia meningoseptica
To assess the synergistic activity of chelators with meropenem, larvae were infected with 20 μL of K. pneumoniae, S. maltophilia, or E. meningoseptica for 1 hour and then injected with 20 μL of a suspension of meropenem or meropenem plus TPEN/nitroxoline (128, 64, and 8 mg/L), and viability assessed over 120 hours. Results obtained showed that both TPEN and nitroxoline work in synergy with meropenem against all tested strains. TPEN was more active than nitroxoline in increasing larval survival relative to meropenem alone. The effects of chelators in combination with meropenem were most active against K. pneumoniae, (Fig. 5). In this case, meropenem resulted in a 10% survival after 48 hours, while combination of meropenem with TPEN (128 mg/L, 0.3 mM) and nitroxoline (128 mg/L, 0.67 mM) resulted in 66% (p < 0.0001) and 59.9% (p < 0.0001) survival 48 hours postinfection, respectively. After 48 hours, no larvae survived for the meropenem alone treatment, while for meropenem plus TPEN 128 and 64 mg/L treatment, survival was 24.12% (p < 0.0001) and 17.63% (p < 0.0001), respectively. Meropenem plus nitroxoline at concentrations of 128 and 64 mg/L resulted in 16.33% (p < 0.0001) and 4.56% (p < 0.001) survival. TPEN, but not nitroxoline, was very active against S. maltophilia in combination with meropenem. After 96 hours, meropenem alone resulted in a 10.23% survival, which was lower than that observed for meropenem plus TPEN at 128 (56.51%, p < 0.05), 64 (34.91%, p < 0.05), and 8 (37.81%, p < 0.01) mg/L. The use of nitroxoline/meropenem combinations was not associated with a statistically significant increase in larval survival compared to meropenem treatment alone (Fig. 5). TPEN 128 (52.38%, p < 0.001) and 64 (32.52%, p < 0.05) mg/L combined with meropenem (64 mg/L) resulted in a significant increase in larval viability after 72 hours of E. meningoseptica infection compared to meropenem alone (8.82%), (Fig. 5).
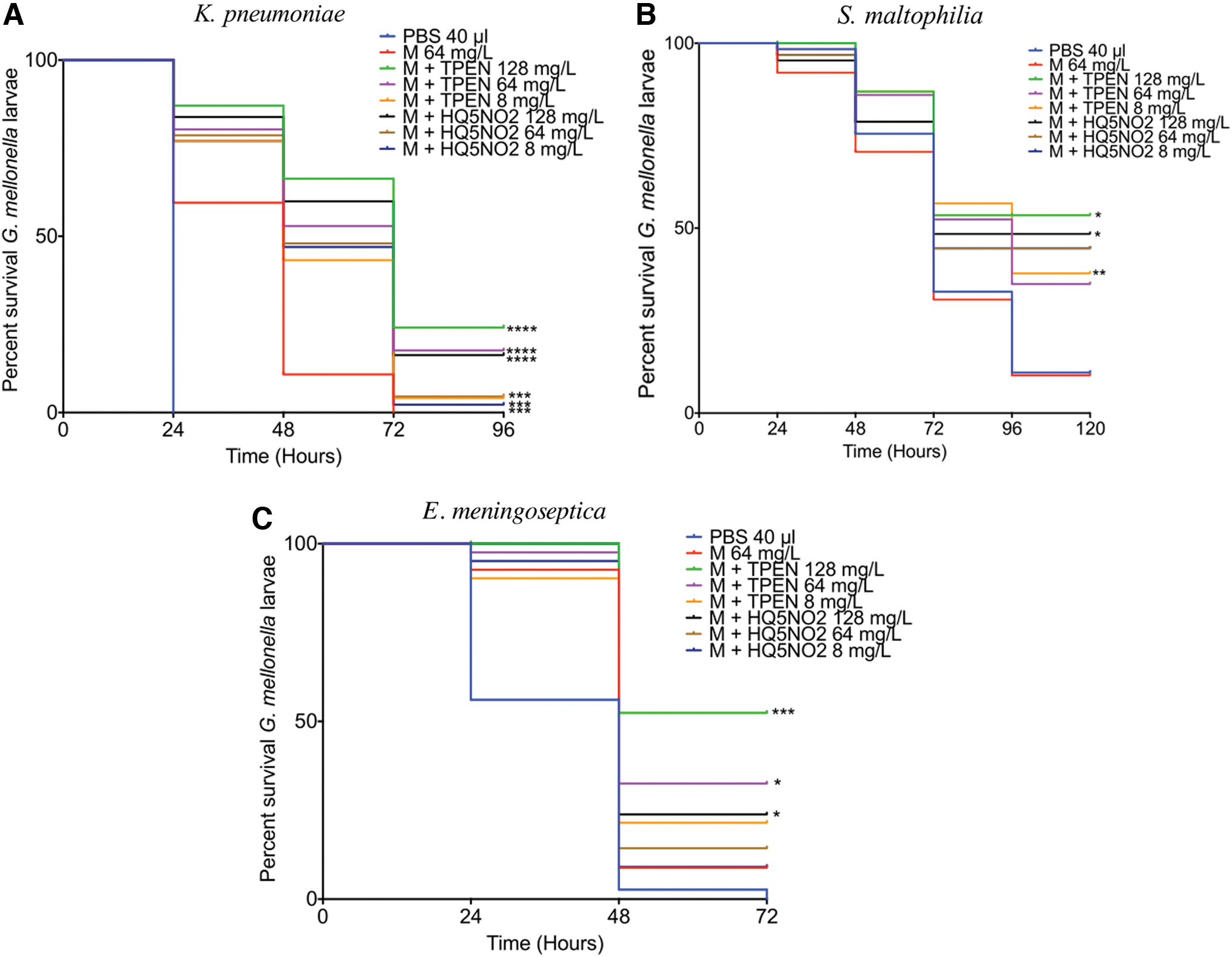
Survival curve of Galleria mellonella larvae following Klebsiella pneumoniae
Discussion
Very limited therapeutic options are currently available for the treatment of infections caused by MBL producers. Furthermore, last-resort agents such as polymyxins are impaired by the risk of either suboptimal or toxic concentrations, at least in the absence of therapeutic drug monitoring, which is unfortunately very often unavailable outside research laboratories.34,38 Therefore, antimicrobial alternatives against MBL producers are urgently needed. Fortunately, some promising agents active against MBL producers are in late phase of clinical development.39–41 However, as demonstrated by the recent experiences with some much awaited and important novel agents active against KPC-producing K. pneumoniae, resistance to new drugs may emerge rapidly after (or even before) their introduction in the market,42,43 and this could be also reasonably expected for novel agents active against MBL producers. Moreover, other mechanisms of resistance, like efflux or impermeability, could have a role in resistance development, also against new agents, potentially influencing their antimicrobial activity alone or in combination with other drugs. Certainly, the dissemination of resistance to novel antibiotics could (and should) be controlled through adequate implementation of antimicrobial stewardship and infection-control interventions, but it remains also imperative not to be unprepared when facing those infections by MBL producers (hopefully a few) that will become resistant to novel agents.
Zinc chelators have been used in human therapeutics, especially for neurological disorders, that is, ischemic stroke and Alzheimer's disease.44,45 In infectious diseases, their use is mostly limited to the administration of nitroxoline in patients with urinary tract infections, where it demonstrated variable results, but an invariable good safety profile.46,47
In this preclinical in vitro and in vivo study, we explored the activity of zinc chelators in combination with meropenem against MBL-producing Gram-negative bacteria. The study was based on the hypothesis that chelation of zinc ions could impair the activity of MBL enzymes. Several zinc chelators were evaluated, but only two (nitroxoline or TPEN) demonstrated possible activity in vitro (alone or in combination with meropenem, with different patterns being observed for the two chelators), and were subsequently tested in vivo. Eventually, the most marked in vivo activity was observed for meropenem/TPEN combinations.
The effect of meropenem, TPEN, and nitroxoline on larval viability was assessed and no drug displayed toxic effects on larval survival over 72 hours. Furthermore, no cuticular melanization in response of the drug was observed and some larvae formed pupa 2 weeks postinjection, indicating the drugs had no adverse effect on larval development. There was also no significant increase in larval hemocyte density following injection with a range of concentrations (8–128 mg/L) of the test compounds, which is a common effect observed with a range of antimicrobial drugs. Therefore, any antimicrobial effect observed should be as a result of the primary antimicrobial activity of the drug and not as a result of increased cellular immune responses. 20
The intrinsic antimicrobial activity of TPEN and nitroxoline was assessed in larvae against infection by E. meningoseptica, S. maltophilia, and K. pneumoniae. Only nitroxoline displayed antimicrobial activity against the bacterium at a concentration of 128 mg/L. With this in mind, the in vivo activity of TPEN and nitroxoline, on reversing the MBL-mediated resistance to meropenem, was assessed in G. mellonella larvae. TPEN was more active than nitroxoline and the effect of the chelators with meropenem was strongest against K. pneumoniae. TPEN was also active in combination with meropenem against S. maltophilia and E. meningoseptica.
In particular, we provided the first in vitro and in vivo description of synergistic activity between TPEN and meropenem against S. maltophilia and E. meningoseptica. Furthermore, although in vitro synergistic activity between TPEN and meropenem against NDM producers had been already demonstrated by other authors, 17 we confirmed this synergistic interaction also in an in vivo model of NDM-producing K. pneumoniae infection. The synergism is expressed as a significant reduction in larvae mortality when TPEN was added to meropenem for each of the abovementioned (carbapenem resistant) bacteria. In vitro synergistic activity was also demonstrated between TPEN and meropenem in C. indologenes, but the bacterium was not able to kill larvae. Furthermore, this study demonstrates the utility of using G. mellonella larvae to test novel therapeutic approaches against clinically relevant, emerging pathogenic and antibiotic-resistant bacteria. Results can be obtained from larvae within 6 and 72 hours, larvae are cost effective and in vivo antibiotic sensitivity may be assessed before identification of the isolate.
Interestingly, a range of spiro-indoline-thiadiazole chelators were found to exhibit synergy with carbapenems against NDM-1-harboring K. pneumoniae. This finding was also demonstrated in a mouse model of infection where mice that received combination treatment displayed a significant decrease in bacterial burden compared to animals that received antibiotic treatment alone. 18 Bacterial zincophore [S,S]-ethylenediamine-N,N′-disuccinic acid also improved survival of K. pneumoniae-infected G. mellonella larvae that were also administrated imipenem, relative to monotherapy. 48 Zinc chelators (1,10-phenanthroline and TPEN) improve survival of A. fumigatus-infected mice alone and in combination with caspofungin. 19 This illustrates the importance of zinc both in bacteria and fungi. These studies highlight the impact zinc chelators may have in tackling antibiotic resistance and also the potential antifungal activity. Our findings also demonstrate the utility of G. mellonella larvae to rapidly assess a range of zinc chelators with an antibiotic of choice, to assess efficacy and synergy, while overcoming the legal, ethical, and economic considerations of using mammals (e.g., mice).
In conclusion, provided they are sufficiently effective and safe in the different phases of clinical development, zinc chelators combined with beta-lactams could become an important last-resort (or maybe even frontline) therapeutic option for infections caused by MBL producers. Nonetheless, further preclinical investigation is needed to precisely identify the most active and promising combinations to be possibly subjected to clinical development, as well as their safety and achievable concentrations in humans.
Footnotes
Acknowledgments
G.S. is the recipient of a Maynooth University Doctoral studentship.
Disclosure Statement
No competing financial interests exist.
Funding Information
This publication has emanated from research supported, in part, by a research grant from Science Foundation Ireland (SFI), and is co-funded under the European Regional Development Fund under Grant No. 12/RC/2275_P2.