
Abstract
Emergence of colistin-resistant bacteria harboring mobile colistin resistance genes (mcr genes) pose a threat for food-producing animals and humans. In this article, we aim to highlight the emergence of Escherichia fergusonii as an important new reservoir to mcr-1-harboring plasmid in poultry production. Three strains closely related were isolated from cloacal swabs. Their genome contains four plasmids, including a 182,869 bp IncHI2 plasmid harboring the colistin resistance gene mcr-1. These results will contribute to our understanding of plasmid-mediated mcr-1 gene presence and transmission in E. fergusonii.
Antimicrobial resistance is becoming a great challenge to public health worldwide. The global spread of the mcr-1 gene is now evident and extensively documented especially in food-producing animals.1–3 Farm animals can serve as a reservoir of antibiotic-resistant bacteria and antibiotic resistance determinants that may be transferred to humans. 4 The systematic use of colistin as growth promotor in livestock production has been increasing the selection pressure for antibiotic-resistant bacteria, especially in Enterobacterales such as Escherichia fergusonii. Furthermore, the emergence of carbapenem-resistant bacteria worldwide and the increased use of polymyxins as “last line” antibiotics to treat human infections may have accelerated the spread of polymyxin resistance. 5
E. fergusonii has now been recognized as an important emerging opportunistic pathogen to both animals and humans.6–9 In animals, E. fergusonii has been isolated from poultry, porcine, and cattle displaying symptoms of salmonellosis-like infections, including diarrhea.7,10,11 In this study, we report the first identification of E. fergusonii harboring IncHI2 plasmid containing mcr-1 mobile colistin-resistant gene from poultry farms in Brazil to the best of our knowledge.
As part of a continued project that aims to monitor colistin resistance and to detect the presence of mcr-harboring bacteria from broilers in Rio de Janeiro, southeastern Brazil, 75 cloacal swabs were collected from small poultry farms between July and October 2019.
Cloacal swab samples were streaked onto Hektoen enteric agar (Difco Laboratories, Detroit, Mich) supplemented with 4 μg/mL colistin (Sigma-Aldrich, St. Louis, MO) and incubated at 35 ± 2°C for 18–20 hr. Antimicrobial susceptibility profiles were determined by automated MicroScan Walkaway system (Siemens Healthcare Diagnostics Limited, Camberley, United Kingdom) and colistin MICs by the broth microdilution method (www.eucast.org). The mcr-1 and extended-spectrum β-lactamases (ESBLs) genes were screened by PCR and sequencing. 12
From 12 colistin-resistant strains (16%) grown onto selective medium, 3 were identified by MicroScan Walkaway system as E. fergusonii. All isolates were positive for mcr-1 gene and negative for ESBL profile by PCR. Species identification was confirmed by 16S rRNA gene sequencing method and compared with other 16S rRNA genes in the GenBank, using BLAST-n (https://blast.ncbi.nlm.nih.gov); and by PCR using primers targeting the yliE and EFER_1569 genes. 13
All mcr-1 positive E. fergusonii strains were only resistant to colistin (CST), tetracycline (TET), nalidixic acid (NAL), and trimethoprim-sulfamethoxazole (SMZ/TMP) (Table 1).
Characterization of the Three mcr-1-Positive Escherichia fergusonii Isolates and the Transconjugant Analyzed in This Study
Resistance genes for isolates and transconjugant were detected by whole-genome sequencing (WGS) and PCR.
Antimicrobial susceptibility tests were interpreted according to the European/Brazilian Committee on Antimicrobial Susceptibility Testing guidelines.
MIC for colistin was obtained using the broth microdilution test, according to the European/Brazilian Committee on Antimicrobial Susceptibility Testing guidelines.
Antimicrobial susceptibility for tetracycline was performed by disk diffusion test.
MIC, minimum inhibitory concentration; NA, not applicable; ND, not determined.
These three isolates were to be closely related by pulsed-field gel electrophoresis (PFGE) (Fig. 1a). The S1-nuclease PFGE was performed to determine the plasmid profiles and four different plasmids (182,869, 90,790, 69,760, and 34,730 bp) were detected in all strains (Fig. 1b). Mating assay was performed by using one of E. fergusonii isolates (isolate A12) as donor and Escherichia coli J53 as recipient. 14 Transconjugants selected onto Hektoen enteric agar supplemented with colistin (4 μg/mL) were found to be mcr-1 positive by PCR and had a colistin resistance (16 μg/mL) 32 times higher than that observed in E. coli J53. In addition, transconjugants showed no resistance to any other antibiotic tested (Table 1). After conjugation, the plasmid profiles were again determined by S1-nuclease PFGE for the transconjugants recovered confirming only one transmissible high-molecular-weight plasmid (pA12_mcr-1; 182,869 bp) (Fig. 1b).
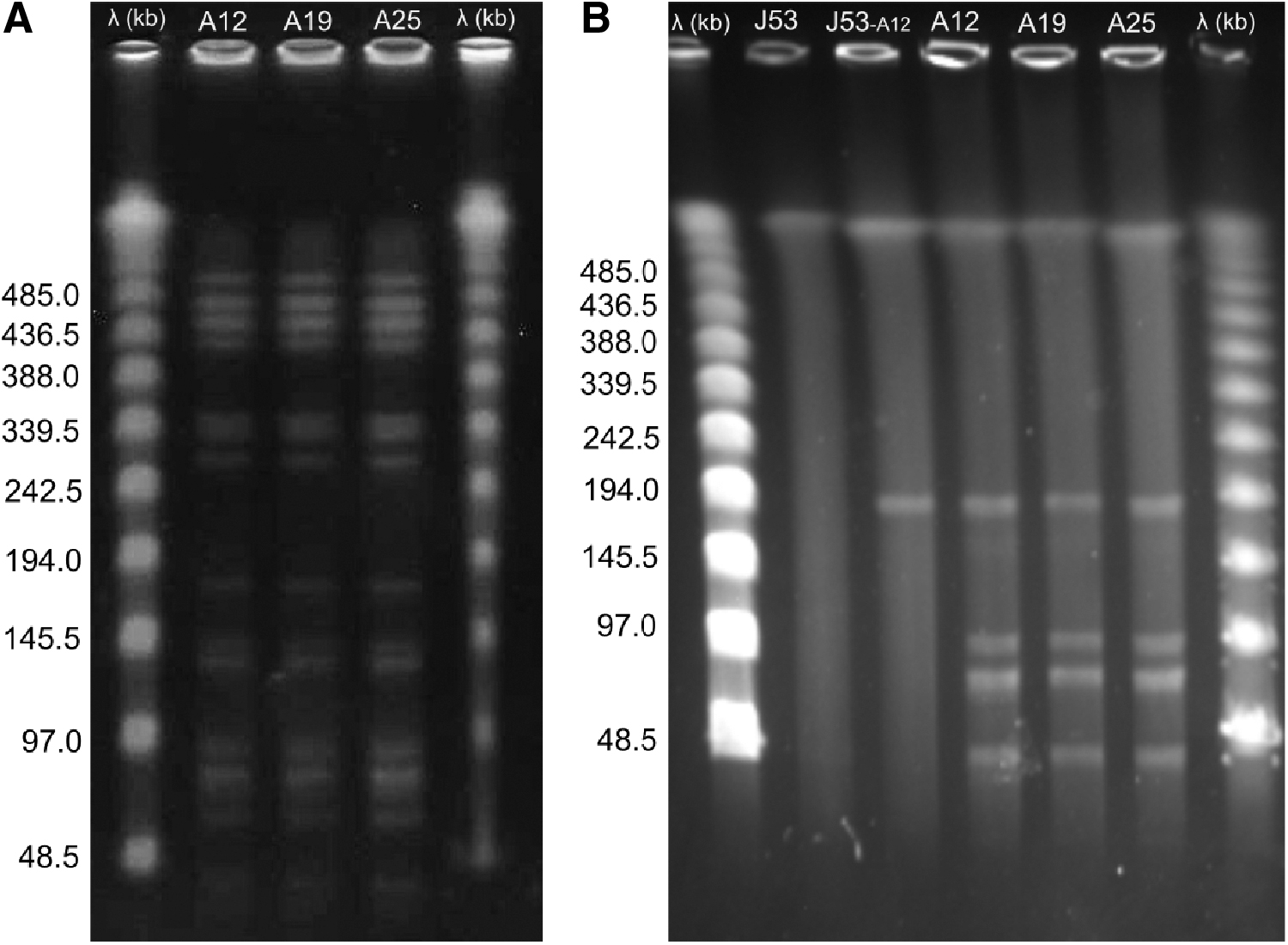
Pulsed-field gel electrophoresis of mcr-1 positive Escherichia fergusonii.
The whole-genome sequencing (WGS) of transconjugant and E. fergusonii A12 were performed using an Illumina MiSeq platform (Illumina, Inc., San Diego, CA). A genomic library was constructed by transposon tagmentation with the Nextera XT DNA Sample Prep kit (Illumina, Inc., USA). Raw reads were trimmed using Trimmomatic 0.36 to average quality score of 20 and 250 bp minimal length. 15 Paired reads were assembled using SPAdes and plasmidSPAdes version 3.10. 16 The assemblies were subjected to ResFinder (https://cge.cbs.dtu.dk/services/ResFinder) and plasmid sequence type (pMLST) (https://pubmlst.org/plasmid) analyses. We adopted the annotations from NCBI Prokaryotic Genome Annotation Pipeline.
For transconjugant, the reads were mapped to E. coli J53 (CP028702.1). Unmapped reads assembly resulted in two main plasmid's contigs. According to BLAST alignments, these contigs have great synteny with plasmid p180-PT54 (CP015833.1). So, the contigs were extended using AlignGraph 17 and the p180-PT54, resulting on a plasmid with 182,869 bp (WYDM02000005.1). Plasmid replicon type IncHI2 was detected for this plasmid (identity 100%, template coverage 100%). The antimicrobial resistance gene mcr-1.1 was the only resistant gene found. Annotation revealed that the pA12_mcr-1 carries genes coding for conjugal transfer proteins (traB, traD, traN, traG, traF, and traN), tellurium resistance (terA, terB, terC, terD, terF, and terZ), type II toxin-antitoxin system HipA family toxin, tyrosine-type recombinase/integrase, pilus assembly protein, toxin-plasmid maintenance system killer protein and nucleases. Insertion sequences annotation using ISFinder 18 revealed the presence of IS1R and an IS66-like element ISEc23 family transposase, this one 14,218 bp downstream mcr-1.1. The pMLST revealed type 4.
The plasmid described in our study carrying mcr-1 was closely related to p180-PT54 (CP015833.1) from E. coli O157, from the same incompatibility group and pMLST, which does not carry ISEc23, mcr-1.1, neither other genes common on IncHI2 plasmids 19 (Fig. 2) (see Supplementary Data). We believe that the insertion of the mcr-1 gene in A12 plasmid is related to ISEc23 and is a different event from those responsible for the insertion of this gene into others IncHI2 plasmids such as pHNSHP45-2 (KU341381) from E. coli and pEF01 from E. fergusonii (NZ_CP040806.1). These two plasmid carry IS5-like and ISApl1 near to mcr-1, respectively, besides numerous resistance determinants.20,21
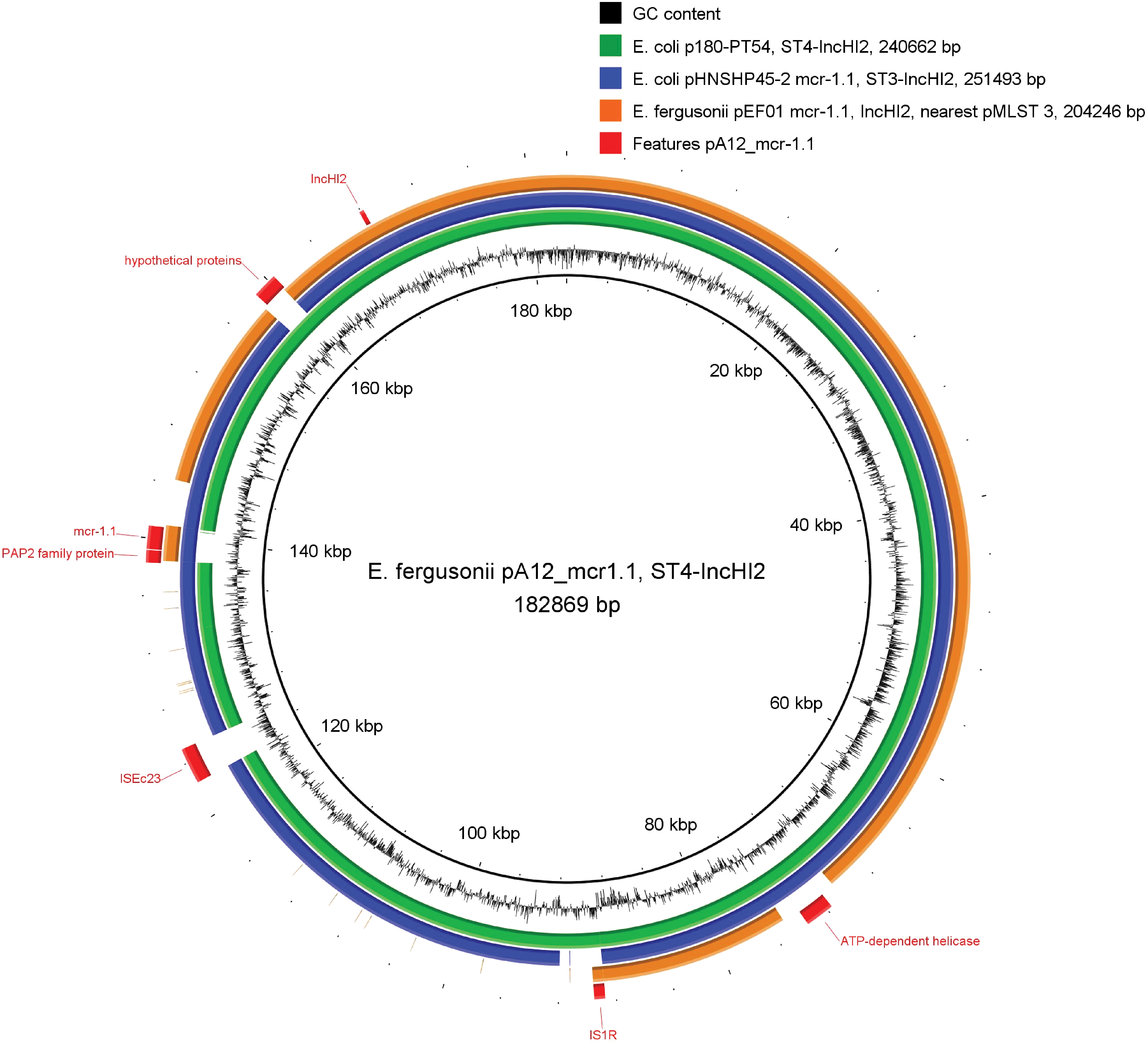
Genomic comparison between IncHI2 plasmids in relation to pA12_mcr-1 from Escherichia fergusonii A12. Inside out, the first ring is plasmid pA12_mcr-1 from E. fergusonii strain A12. The second ring is the pA12_mcr01 GC content. The outer rings represent the nucleotide sequence of the corresponding DNA region of the following IncHI2 plasmids in different colors against the reference genome (pA12_mcr-1): p180-PT54 from E. coli O157 (ST4-IncHI2) (green), pHNSHP45-2 from Escherichia coli strain SHP45 (ST3-IncHI2, mcr-1) (blue), and pHLJ179-85 from E. fergusonii (IncI2, mcr-1) (orange). The last one is the CDS from pEF01 (red). Color images are available online.
Paired reads from E. fergusonii A12 WGS were mapped to the mcr-1 plasmid. The unmapped reads were assembled with SPAdes and scaffolded using 4 E. fergusonii reference chromosome (NC_011740.1, NZ_CP042945.1, NZ_CP042942.1, and CP040805.1). Four chromosomal contigs, a total of 4.938293 bp were obtained. Twenty-seven plasmid's contigs >500 bp resulted from assemble with plasmidSPAdes using A12 unmapped reads. Among them, WYDM02000006.1 (50,063 bp), WYDM02000026.1 (2,800 bp), and WYDM02000007.1 (36,913 bp) have replicon types IncB/O (identity 100% to reference B/O plasmid [M28718]), IncX1 (identity 100%, template coverage 100%) and p0111 (identity 99.1%, template coverage 100%), respectively. The antimicrobial resistance genes dfrA1, aadA1, tet(B), sul2, and sat-2 were found in these contigs. No resistance gene was found on chromosomal contigs.
Using BLAST-n, we search for virulence genes aggR, astA, cnf2, cvi, eae, hlyF, ibeA, ipaH, iroN, irp2, iut, ompT, stx1, tsh, and usp. No gene was detected in mcr-1's plasmid nor in chromosome. Only iut, responsible for siderophore uptake, was found in plasmid contig WYDM02000010.1.
The mcr genes have been found on diverse plasmid backbones, including IncHI2 (IncI2, IncX1, IncX4, IncFIA, IncFIB, IncFIC, and IncHI1) with high in vitro transfer rates. They are often harbored together with other resistance determinants, such as β-lactamases genes.22–24 Although ISApl1-mediated complex transposition has been proposed to play a critical role in the spread of mcr-1 gene, 25 our results showed no ISApl1 elements flanking the mcr-1 gene from IncHI2 plasmid nor β-lactamases genes. Data are consistent with previous reports showing the lack of ISApl1 in conjugative plasmid of E. coli, E. fergusonii, and Klebsiella pneumoniae strains isolated from poultry and human. 11 Thus, multiple mcr-1 gene-bearing plasmids with different Inc-types and ISs could be actively involved in colistin resistance.
Another aspect that might contribute to colistin resistance is the co-selection of colistin-resistant bacteria and genes by other antibiotics.22,24 In this study, colistin-resistant E. fergusonii isolates were also resistant to TET, NAL, and SMZ/TMP. These antibiotics are widely used in food-producing animals and could potentially enhance mcr-1 gene transfer promoting the emergence of several colistin-resistant bacteria in poultry farm. 26
Considering colistin resistance a serious situation, in November 2016, the Brazilian Ministry of Agriculture, Livestock, and Supply (MAPA) banned the use of colistin as a feed additive for animal growth promotion purpose (regulatory instruction no. 45 [www.agricultura.gov.br]), following the international recommendations of the World Health Organization (WHO). Despite government action, our group recently detected a high prevalence of mcr-1 gene as the only resistance gene in E. coli strains isolated from broilers in several poultry farms in Rio de Janeiro (unpublished data). Furthermore, previous surveillance studies showed evidence that mcr-1 gene has been circulating in food-producing animals in Brazil since 2012. 27 The mcr-1 gene was also detected in commercial chicken meat sold in markets in São Paulo, southeastern Brazil, contributing to worsen the scenario. 28 Together, data suggest that E. coli and now E. fergusonii are important reservoirs to colistin resistance mcr-1 gene in poultry farms in Brazil.
The poultry meat has become an inexpensive source of protein and its impact of transferring resistance cannot be neglected. The overuse of antibiotics will promote the unrestricted expansion and circulation of drug-resistant strains, such as IncHI2-harboring E. fergusonii, among human–animals environment. Therefore, great concern should be given to continuous surveillance, improving prevalence data in both human and veterinary settings in our country.
Accession Number
IncHI2 plasmid (pA12_mcr1) and E. fergusonii A1 sequences were deposited in the GenBank database under accession number WYDM00000000.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This project was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (Grant number: 402524/2018-7), Fundação Carlos Chagas de Amparo à Pesquisa, and Instituto Oswaldo Cruz (FIOCRUZ).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.