
Abstract
The study investigates the prevalence of antimicrobial resistance in Campylobacter jejuni and Campylobacter coli in gastroenteritis patients in the eight most populous regions in Australia and compares the prevalence of antimicrobial resistance in Europe and North America. A total of 164 Campylobacter isolates were collected from patients with campylobacteriosis and tested for susceptibility to six antimicrobials using ETEST® strips and compared with reports from Europe and the United States. Genomes were sequenced on Illumina NextSeq to identify genetic determinants of resistance. Phenotypically, 1.8%, 14.0%, 14.6%, and 20.1% of isolates were resistant to erythromycin (ERY), ampicillin, tetracycline (TET), and ciprofloxacin (CIP), respectively. Comparing published phenotypic results of antimicrobial resistance in several European countries and the United States with these Australian isolates reveals that rates observed in Australia are among the lowest observed for ERY, CIP, and TET for both C. coli and C. jejuni. For each antimicrobial tested, concordance between resistance phenotype and genotype ranged from 66.6% to 100.0%. This study highlights that, among industrialized countries, Portugal and Spain have very high levels of antimicrobial resistance in C. jejuni and C. coli, especially when compared with the United Kingdom, United States, and Australia.
Introduction
C
Macrolides such as erythromycin (ERY), clarithromycin, or azithromycin are typically prescribed to treat such cases. 3 Alternatively, in cases of undiagnosed diarrheal illness, fluoroquinolones such as ciprofloxacin (CIP) are commonly prescribed when necessary, as they are additionally effective against Salmonella spp.. 4 Tetracycline (TET) and β-lactam antibiotics have also been used in treating gastrointestinal illnesses. 5 In addition, in vitro trials have shown that gentamicin (GEN), clindamycin, meropenem, and telithromycin are promising alternative campylobacteriosis therapies.4–6
Campylobacter resistant to commonly used antimicrobials such as macrolides and fluoroquinolones has been recognized as a public health threat by the US Centers for Disease Control and Prevention (CDC) because of their potential to cause more severe outcomes, including longer duration illness and higher risk of invasive infection or death. 7 Many of the antimicrobials used to treat campylobacteriosis, including aminoglycosides, macrolides, penicillins, and quinolones, have been classified by the World Health Organization as critically important for human medicine 8 ; thus, monitoring resistance to these antimicrobials is imperative.
The prevalence of antimicrobial resistance varies significantly across the globe and is likely influenced by local practices relating to the use of antimicrobials. 5 In China, antimicrobials including fluoroquinolones are used extensively as they are available without prescription and the prevalence of resistance in Campylobacter from diarrheal patients to CIP (89.7%), TET (74.6%), and nalidixic acid (69.0%) is very high. 9 In contrast, the prevalence of CIP resistance in clinical Campylobacter isolates in the United States is significantly lower (25%).5,10 Previous reports suggest the prevalence of antimicrobial resistance in Campylobacter species in Australia is markedly lower than most high-income countries, but these reports have only examined isolates representing a small geographic area of Australia.3,11,12
Although less prevalent than C. jejuni, symptoms of C. coli infection are clinically indistinguishable from C. jejuni. 5 Many studies have examined the susceptibility of C. jejuni to common antimicrobials,3,11–14 but only a limited number of reports have characterized antimicrobial resistance in C. coli. In Australia, Campylobacter isolated from human diarrheal samples are only identified to genus level,15,16 thus the findings presented here provide novel insights into the distribution of species responsible for campylobacteriosis in Australia.
The aims of this study were to (1) investigate the prevalence of resistance in clinical isolates of C. jejuni and C. coli to commonly used antimicrobials across Australia, (2) identify Campylobacter antimicrobial resistance genotypes using whole-genome sequencing (WGS) and compare these with in vitro antimicrobial susceptibility testing (AST) resistance phenotypes and (3) compare Australian isolates prevalence of resistance to CIP, ERY, and TET to those from Europe and the United States.
Materials and Methods
Collection of Campylobacter isolates
Campylobacter isolates (C. jejuni n = 137; C. coli n = 27) were collected from fecal samples obtained from humans in the Australian Capital Territory (ACT), New South Wales (NSW), Northern Territory (NT), Queensland (Qld), South Australia (SA), Tasmania (Tas), Victoria (Vic), or Western Australia (WA), in 2018 or 2019. Samples collected from individuals in the ACT, NSW, and Qld were from cases of domestically acquired infections and were collected as part of the CampySource study. 17 Travel history was not considered as part of the selection of samples in the remaining states and territories. Fecal samples were stored at 2–8°C and processed within 48 hours of collection.
Arbitrarily, 20 isolates, except WA, where 30 isolates were used owing to geographical size of this state relative to the other states and territories, were obtained from clinical laboratories in each of the states and territories (Fig. 1). Samples were collected from October 2018 to December 2018. In the NT, only two samples were obtained during this time period; thus, additional samples were subsequently collected from December 2018 to February 2019 from NT (n = 6) and northern WA (n = 8; similar climate to the northern part of NT). Northern WA samples were collected from Karratha, Cable Beach, Carnarvon, Derby, Broome, and Wickham. Figure 1 summarizes the number and location of isolates tested from each state or territory.

Geographic location of the clinical Campylobacter isolates, based on patient location, collected in the Australian Capital Territory, New South Wales, Northern Territory, Queensland, South Australia, Tasmania, and Victoria from October to December 2018 and in Northern Territory and Western Australia from October 2018 to February 2019. Note “n” represents the number of isolates.
Human diarrheal fecal samples were cultured on CampyFood® (bioMerieux, Marcy-l'Étoile, France) or Campylobacter agar (Oxoid, Hampshire, United Kingdom) and incubated microaerobically (85% N2, 10% CO2, 5% O2) at 42°C for 48 hours. Presumptive Campylobacter spp. were confirmed as C. jejuni or C. coli using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry or polymerase chain reaction 18 and were transferred to a charcoal swab (BD, Franklin Lakes, NJ) and sent to the Microbiological Diagnostic Unit Public Health Laboratory (MDU PHL) at the Doherty Institute for further characterization.
In vitro AST
The use of AST hereafter refers solely to phenotypic testing. The activity of amoxicillin–clavulanic acid (AMO), ampicillin (AMP), CIP, ERY, GEN, and TET were tested against 137 C. jejuni and 27 C. coli isolates. Minimum inhibitory concentrations (MICs) were determined by ETEST® (Epsilometer test; bioMérieux). 13 Epidemiological cutoff (ECOFF) values, determined as per the European Committee on Antimicrobial Susceptibility Testing (EUCAST), were used to determine the microbiological resistance to AMO (C. coli > 8 mg/L; C. jejuni > 16 mg/L), AMP (>8 mg/L), CIP (>0.5 mg/L), ERY (C. coli > 8 mg/L; C. jejuni > 4 mg/L), GEN (>2 mg/L), and TET (C. coli > 2 mg/L; C. jejuni > 1 mg/L). 19
For some antimicrobials (AMO, ERY, and TET), the MIC breakpoint or ECOFF value was species dependent. C. jejuni NCTC11351 (ATCC 33560) was used as the quality control strain as described in the ETEST Application Guide. 20 Isolates with a nonwild-type interpretation were classified as “resistant,” whereas isolates with a wild-type interpretation were classified as “susceptible.” Isolates resistant to three or more antimicrobials were classified as multidrug resistant (MDR). Because of the low number of C. coli isolates (n = 27), C. coli and C. jejuni were analyzed together.
Genomic DNA extraction and WGS
Genomic DNA was extracted from the 164 Campylobacter isolates using the QIASymphony® DSP DNA Mini kit (Qiagen) with an automated DNA extraction robot (QIAsymphony; Qiagen) following manufacturer's instructions. Sequencing libraries were prepared using the Nextera® XT DNA sample preparation kit (Illumina, San Diego, CA) and sequenced on the Illumina NextSeq500® (150 base, paired-end read sets).
Bioinformatic analyses
The paired-end read set for each isolate was analyzed using the Nullarbor pipeline v2. The quality of each of the read sets was evaluated to ensure sufficient read coverage depth (min. 50 × ) and isolate purity (lack of contaminating reads using Kraken and total assembled bases within 20% of the average genome size in the analysis group). Read sets for each isolate were aligned to a reference genome; either NC_003912 (for C. jejuni isolate analysis) or NC_022132 (for C. coli isolate analysis) using Snippy v4.3.6/BWA-MEM v0.7.17-r1188. The read sets for each isolate were assembled to produce a draft genome sequence using SPADes v3.13.0 and the sequence annotated using Prokka v1.13.321 as part of the Nullarbor pipeline.
Identification of genetic determinants of resistance
The ResFinder v3.1 resistance gene database was used for identifying antibiotic resistance genes in each draft genome sequence using Abricate v.0.8.12; hits were filtered using a cutoff set at 50% sequence coverage and 95% nucleic acid sequence identity. The presence of a resistance enhancing variant (RE-CmeABC) of the predominant efflux pump in Campylobacter, CmeABC, was also examined as described previously. 22 Chromosomal mutations associated with antimicrobial resistance genotypes were investigated in the gyrA (quinolone resistance determining region), 23S rRNA, and blaOXA-61 genes for each isolate.
The nucleotide sequences of the 23S rRNA and blaOXA-61 genes were extracted from the draft genome sequence of each isolate using BLAST 23 and a multiple sequence alignment produced using Clustal-Omega. 24 Similarly, the GyrA amino acid sequence was used as the query for a tblastn to extract input sequences for a multiple alignment of the GyrA protein from each isolate. The amino acid at position 86 of GyrA and the nucleotides at position 2,074 and 2,075 in the 23 rRNA gene were used to determine the resistance genotype for resistance to quinolones 25 and macrolides/lincosamides/ketolides, 26 respectively. In addition, a point mutation at position 57 in the blaOXA-61 gene promoter region that prevents its transcription, rendering an isolate sensitive to AMP despite the presence of the gene, 27 was examined in addition to blaOXA-61 gene detection.
Concordance of susceptibility phenotypes and genotypes
The concordance between AST-based resistance phenotype and resistance genotype based on the genome sequence analysis described previously was determined for each of the six antimicrobials tested.
International antimicrobial susceptibility comparison
Australian C. jejuni and C. coli antimicrobial resistance phenotypes were compared with resistance data from clinical Campylobacter isolates from Europe and the United States reported by the European Food Safety Authority (EFSA) 28 and CDC, 29 respectively. Supplementary Table S1 summarizes the number of isolates tested in each country for susceptibility to ERY, CIP, and TET.
Antimicrobial resistance data on human Campylobacter isolates collected in 2017 in Europe were extracted from an EFSA summary report. 28 Only European countries that tested at least 10 isolates and reported antimicrobial susceptibility test results for three antimicrobials (ERY, CIP, and TET) were included in the analysis. In Europe, antimicrobial resistance was assessed using the disk diffusion, dilution, or dilution with gradient strip methods and results interpreted by the European Centre for Disease Prevention and Control (ECDC) using EUCAST criteria 19 as described previously. 28 In Europe, Campylobacter species are not routinely tested for sensitivity to AMO or GEN.
Antimicrobial resistance data on human C. jejuni (n = 991) and C. coli (n = 109) isolates collected in 2017 in the United States were obtained from the CDC's National Antimicrobial Resistance Monitoring System (NARMS) “NARMS Now: Human Data.” 29 In the United States the Trek Sensititre® system was used to assess antimicrobial susceptibility to nine antimicrobial agents—including ERY, CIP, and TET—and EUCAST ECOFFs 19 were used for interpretation. 30 Susceptibility data for AMO is not reported in the NARMS Now: Human Data and the majority (80%) of the states in the United States do not report results for GEN.
Nucleotide sequence accession numbers
Read sets for each of the 164 C. jejuni and C. coli isolates generated in this study have been submitted to the Sequence Read Archive at the National Center for Biotechnology Information (NCBI) under Bioproject PRJNA560409 (relates exclusively to this study). The run accession numbers associated with each isolate are presented in Supplementary Table S2.
Ethical consideration
This study was approved by the Australian National University Human Research Ethic Committee (Reference No. 2016/426), ACT Health Human Research Ethics Committee (Reference No. ETH.8.17.168), and Hunter New England Human Research Ethics Committee (Reference No. 17/08/16/4.03). No identifying human data were collected with Campylobacter isolates from NT, SA, Tas, Vic, or WA. Human ethics approval was obtained for patients with campylobacteriosis in the ACT, NSW, and Qld as part of the CampySource case–control study. 17
Data analysis
Data manipulation and statistical analysis were performed with R (version 3.5.3). 31 The rgeos 32 and rgdal 33 packages in R were used to generate a map of Australia to summarize locations and numbers of isolates. Isolates were aggregated and plotted by Australian Bureau of Statistics' Statistical Areas Level 4 (SA4) unless the SA4 size exceeded 100,000 km2 (in which case postcode area was used, if reported) or the SA4 was within one of the Greater Capital City Statistical Areas (in which case they were aggregated at this latter, higher level). 34
A descriptive analysis was used to compare phenotypic AST results with resistance results from Europe and the United States. Ninety-five percent confidence intervals (CIs) for the prevalence of resistance in Australian, European, and U.S. isolates were calculated using the binom package 35 in R with ggplot236 used to plot associated results.
Results
Among the 164 isolates that represent a snapshot of human campylobacteriosis across Australia in late 2018 and early 2019, we observe that the majority (84%) of cases are caused by C. jejuni (Fig. 2). The prevalence of C. coli was higher in Qld, WA, and SA, where 25% (n = 5), 24% (n = 9), and 20% (n = 4), respectively, of isolates were identified as C. coli (Fig. 2). In Vic, 95% of the isolates were C. jejuni, with the remainder C. coli (5%).
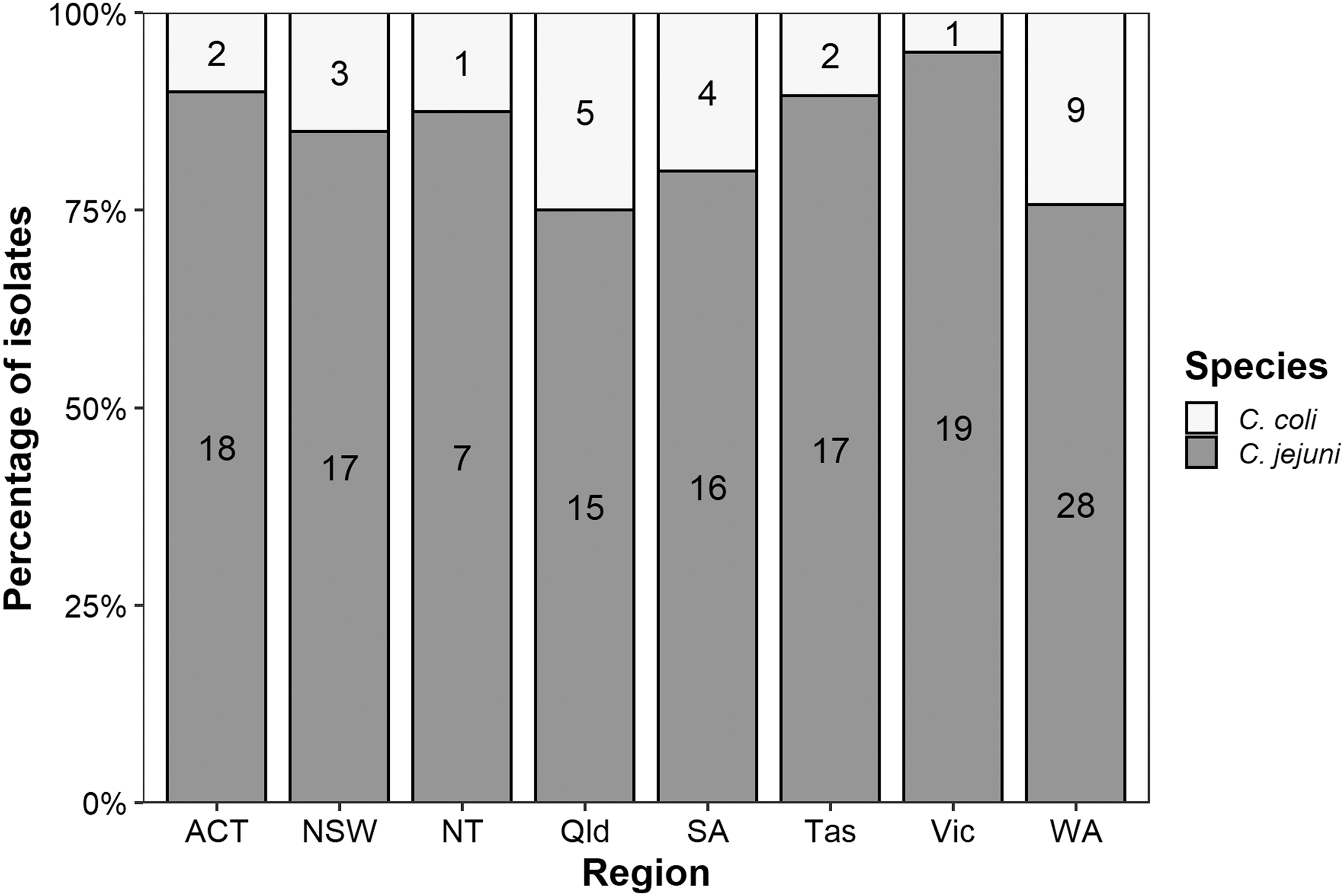
Distribution of clinical Campylobacter jejuni (n = 137) and Campylobacter coli (n = 27) isolates collected in eight regions (states and territories) within Australia. Respective species numbers from each state or territory are noted within each bar.
Among Australian isolates, the highest prevalence of antimicrobial resistance was observed for CIP (20.1%, CI95 = 14.3–27.1%), TET (14.6%, CI95 = 9.6–21.0%), and AMP (14.0%, CI95 = 9.1–20.3%) (Fig. 3 and Supplementary Table S3). Only three Campylobacter isolates were phenotypically ERY resistant and none were resistant to AMO or GEN. Of the 164 Australian Campylobacter isolates tested, 116 (70.7%) were susceptible to all antimicrobials tested (Fig. 4). Of the 164 isolates tested, 7 (4.3%) C. jejuni isolates were MDR (Fig. 4). Six of these were resistant to three, and one resistant to four drug classes. MDR isolates were observed in ACT, Tas, Vic, and WA (Supplementary Table S4). All ERY-resistant isolates were also CIP resistant, with two of these isolates also TET resistant. The prevalence of resistance to one or more antimicrobial was lowest in NSW (15%) and highest in NT and WA (38% each) (Supplementary Table S4). WGS revealed that three of the seven (43%) phenotypically MDR isolates contained a variant of the CmeABC efflux pump (RE-CmeABC).

Prevalence of phenotypic resistance of 164 clinical Campylobacter isolates tested for their susceptibility to AMO, AMP, CIP, ERY, GEN, and TET. Error bars indicate 95% confidence intervals. The number of isolates resistant to each antimicrobial is noted above the respective error bar. AMO, amoxicillin–clavulanic acid; AMP, ampicillin; CIP, ciprofloxacin; ERY, erythromycin; GEN, gentamicin; TET, tetracycline.

Antimicrobial resistance profiles of 164 clinical Campylobacter isolates tested for their phenotypic susceptibility to AMO, AMP, CIP, ERY, GEN, and TET. The number of isolates possessing each resistance profile is noted above the respective bar.
For five of the six antimicrobials tested, there was a high concordance (>85%) between resistant/susceptible phenotypes and genotypes (Table 1). The exception was ERY, which had lower phenotype–genotype concordance (66.6%). Of the three phenotypically ERY-resistant isolates, one isolate had a mutation at position 2,074, namely A2074T (18S1008H1), in the 23S rRNA gene known to confer resistance to macrolides, one isolate (19W1001H1) possessed the erm(B) gene. The remaining isolate (18T1005H1) had no genes or point mutations known to confer ERY resistance.
Comparison of Campylobacter Antimicrobial Resistance Phenotypes Based on In Vitro Antimicrobial Susceptibility Testing and Resistance Genotypes Based on Whole-Genome Sequencing
AMO, amoxicillin–clavulanic acid; AMP, ampicillin; CIP, ciprofloxacin; ERY, erythromycin; GEN, gentamicin; R, resistant; S, susceptible; TET, tetracycline.
All phenotypically CIP- or TET-resistant isolates had a genetic mutation or resistance gene known to confer resistance to the respective antimicrobial. The majority (79%; 129/164) of isolates tested in this study contained the blaOXA-61 gene, but only 12% (16/129) had a functional promotor genotype (i.e., a “T” at base 57). Seven of the 23 isolates that were phenotypically resistant to AMP had genotypically inactive blaOXA-61 gene promotors, although 2 possessed the blaOXA-184 and 2 the blaOXA-185 gene. For the remaining three phenotypically AMP-resistant isolates, two C. coli isolates had a novel A → G mutation at position 59 in the blaOXA-61-gene promoter region. This novel point mutation (A59G) was absent in all other isolates possessing the blaOXA-61 gene.
Among C. jejuni isolates from Europe, >66% were resistant to CIP, with very high proportions resistant in Portugal (96.5%, CI95 = 93.4–98.4%), Lithuania (91.5%, CI95 = 88.9–93.7%), and Spain (88.6%, CI95 = 84.3–92.1%) 28 (Fig. 5A). The lowest prevalence of CIP resistance in C. jejuni were found in Australia (19.4%, CI95 = 13.2–27.0%), Norway (24.5%, CI95 = 19.5–30.1%), and the United States (26.8%, CI95 = 24.1–29.7%). On average, <5% of C. jejuni isolates in Australia, Europe, and the United States were ERY resistant. TET resistance was observed in all countries examined, with the highest prevalence of TET resistance in C. jejuni observed in Portugal (81.9%, CI95 = 76.6–86.4%), Malta (81.3%, CI95 = 54.4–96.0%), Spain (80.4%, CI95 = 75.2–84.9%), and Cyprus (80.0%, CI95 = 59.3–93.2%). 28
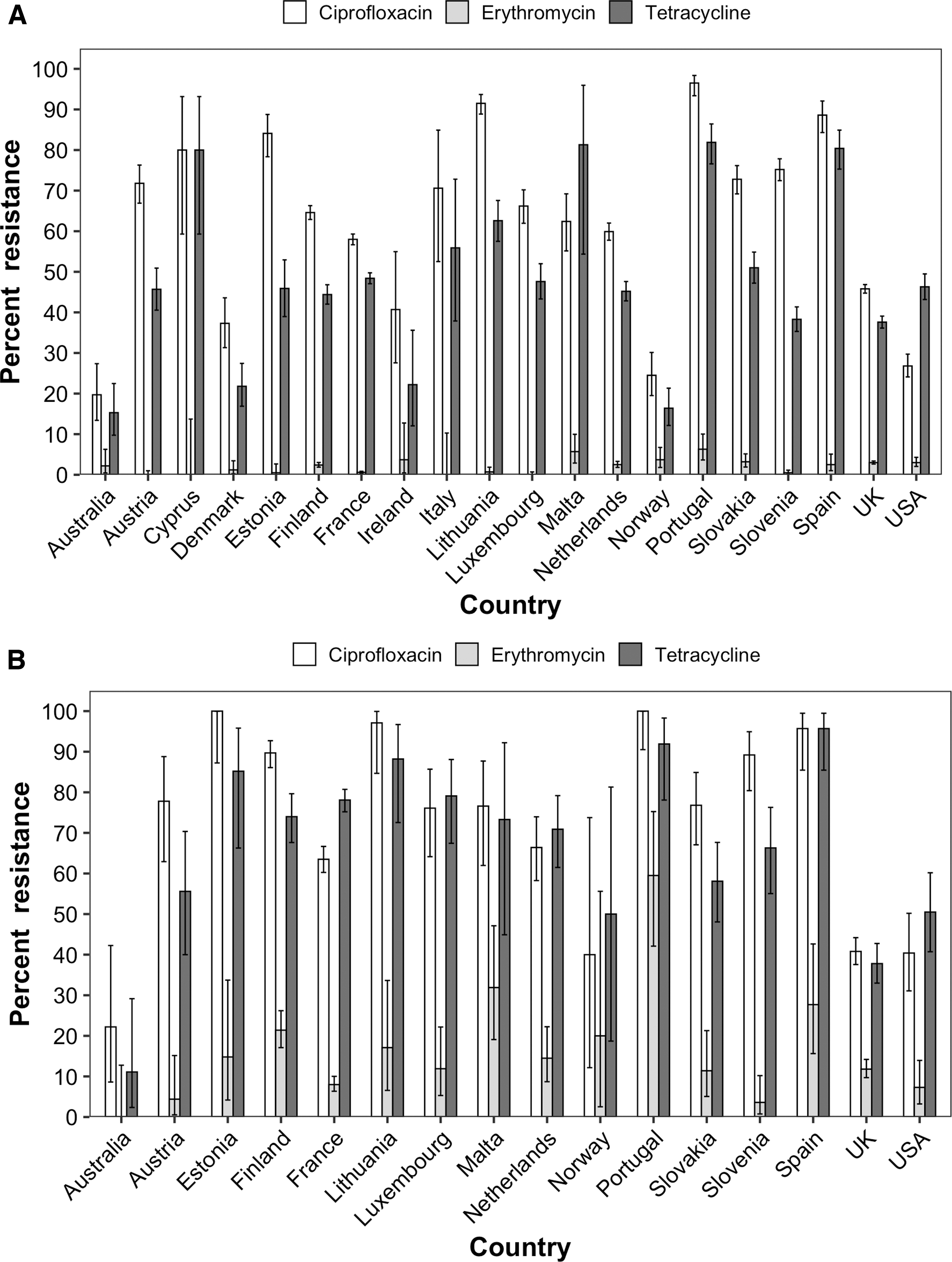
Antimicrobial resistance of
Among C. coli isolates from Europe, >77% of isolates were CIP resistant, with very high proportions resistant in Estonia (100.0%, CI95 = 87.2–100.0%), Portugal (100.0%, CI95 = 90.5–100.0%), Lithuania (97.1%, CI95 = 84.7–99.9%), and Spain (95.7%, CI95 = 85.5–99.5%) 28 (Fig. 5B). The lowest prevalence of CIP resistance in C. coli were found in Australia (24.0%, CI95 = 9.4–45.1%). On average, 14.3% of C. coli isolates in Europe and 7.3% of isolates in the United States were ERY resistant. Among the Australian C. coli isolates tested, none were phenotypically ERY resistant. TET resistance was observed in all countries examined, with the highest prevalence of TET resistance in C. coli observed in Spain (95.7%, CI95 = 85.5–99.5%), Portugal (91.9%, CI95 = 78.1–98.3%), Lithuania (88.2%, CI95 = 72.5–96.7%), and Estonia (85.2%, CI95 = 66.3–95.8%), whereas the lowest prevalence of resistance was found in Australia (12.0%, CI95 = 2.5–31.2%). 28
Discussion
In this study, we characterized the resistance phenotypes of 164 clinical C. jejuni and C. coli isolates using a six-drug panel and characterized respective resistance genotypes using WGS. Although there have been some reports describing antimicrobial resistance of C. jejuni in Australia, these were generally limited in scope to isolates collected in one state,3,11,37 with the exception of one study that reported antimicrobial resistance prevalence in Qld, SA, Tas, Vic, and WA in 2001–2002. 12 In this study, we characterized the resistance phenotypes and genotypes of two Campylobacter species (C. jejuni and C. coli) collected from eight Australian states and territories.
The prevalence of resistance in locally acquired C. jejuni isolates from five Australian states in 2001–2002 to AMP, ERY, CIP, and TET was 46.0%, 2.6%, 2.4%, and 6.6%, respectively. 12 In comparison, we found a lower prevalence of AMP resistance (14.0%), comparable levels of ERY (1.8%) and TET resistance (14.6%), and higher levels of CIP resistance (20.1%). However, our findings correspond well with a recent report from NSW where the percentage of clinical C. jejuni resistant to AMP, CIP, and TET was 15.4%, 13.7%, and 5.1% respectively. 3 We found prevalence of CIP resistance in Australia has increased markedly since 2001. 12 A contributing factor to the higher levels of resistance observed in isolates from NT and WA (each 38% c.f. average 26%) may be because of the higher frequency of travel to Southeast Asia, where antimicrobials are used without restriction by Australians in these jurisdictions. 38
Among MDR isolates in this study, 43% (3/7) possessed the CmeABC efflux pump variant (RE-CmeABC), which has been shown to increase bacterial resistance to multiple antimicrobial classes. 22 In agreement with previous reports that have shown that the prevalence of RE-CmeABC is higher in C. jejuni, 22 this variant was not detected in any C. coli isolate. The low prevalence of multidrug resistance among Campylobacter across Australia agrees with a recent finding in NSW of only 1% MDR C. jejuni. 3
There was a high concordance between in vitro phenotypic resistance to a given antimicrobial and presence of one or more resistance genes or point mutations known to confer resistance to that drug. For four of the antimicrobial agents, AMO, CIP, GEN, and TET, there was a 100% correlation between phenotypic resistance and the presence or absence of putative resistance genes or mutations. However, only an 87.0% concordance between phenotype and genotype was observed for isolates resistant to AMP. Two isolates that were phenotypically resistant to AMP possessed a novel A59G point mutation in the promoter region of the blaOXA-61 gene, which was absent in all other isolates tested and we believe has not been reported previously. Further studies are warranted to determine whether this point mutation, like the G → T transversion upstream of the blaOXA-61 gene, 27 also upregulates blaOXA-61 expression. Additional genetic factors influencing AMP resistance may be responsible for the discrepancies where no blaOXA-61 gene point mutations were identified, but the isolates were phenotypically resistant to AMP. Thus, further studies on AMP resistance mechanisms in Campylobacter are warranted.
Previous reports have shown a 100% correlation between ERY resistance phenotype and genotype.39,40 For the two ERY-resistant isolates for which the genotype could be used to explain phenotype, one isolate contained the erm(B) gene, which confers resistance to macrolides through methylation of the A2058 site in 23S rRNA, 41 and seven other resistance determinants 42 and the other had the A2074T mutation in the 23S rRNA gene. The erm(B) isolate and the isolate with genetically unexplained phenotype represent mechanisms for resistance that have not been previously observed in Australia; these isolates warrant further attention because of the importance of macrolide therapy in the treatment of campylobacteriosis.
Unlike other mechanisms of resistance that tend not to persist in the absence of selective pressure from antimicrobials, the erm(B) gene is present on a genomic island with other resistance genes, and it is likely that carriage of this gene can be sustained by pressure from antimicrobials other than ERY. 28 In China, an increase in the prevalence of the erm(B) gene has been observed since 2014. 43 Overall, our findings showing a high concordance between resistance phenotype and genotype, in agreement with other reports that have compared phenotypic AST with WGS results,39,40,44,45 suggest in silico analysis can be used to accurately predict phenotypic resistance.
Although known genetic determinants of resistance accounted for most of the phenotypic resistance observed, some isolates exhibiting phenotypic resistance did not possess the expected resistance genes or point mutations. For example, based on our current understanding of the mechanisms of resistance for AMP in Campylobacter, WGS would have incorrectly predicted the resistance phenotype of three isolates, which lacked all the genetic determinants known to confer AMP resistance. However, WGS in combination with AST can identify novel resistance mechanisms. This was the case for one WA isolate that was phenotypically ERY resistant and lacked mutations in the 23S rRNA gene but contained a unique multidrug resistance genomic island not previously observed in Campylobacter in Australia. This highlights the importance of determining the resistance phenotype in combination with WGS to infer the likely mechanism of resistance. Significantly, this not only allows us to identify isolates that may possess novel mechanisms of antimicrobial resistance (AMR), allowing these isolates to be specifically targeted for further characterization, but it also ensures that genetic inference of phenotype (based on genome sequencing) is accurate. In particular, this approach is essential for high-quality genomics-based inference of AMR phenotype in Public Health Microbiology.
In comparison with AST results from Europe and the United States, Australia has a very low prevalence of ERY, CIP, and TET resistance in C. jejuni and C. coli. Similar to results from Europe 28 and the United States, 29 the low prevalence of macrolide resistance in this study was not surprising as mutations in the 23S rRNA genes generally incur a fitness cost on Campylobacter28,46 and is consistent with the low use of antibiotic growth promotors in primary production in Australia.
The prevalence of CIP resistance in Australia (20.1%) was higher than expected as fluoroquinolones are not approved or used in Australian livestock. 47 Therefore, it is unlikely CIP resistance in domestically acquired Campylobacter infections evolved from local selective pressure. However, a recent report by the Australian Chicken Meat Federation found 14.8% of C. jejuni and 5.2% of C. coli recovered from poultry were fluoroquinolone resistant. 47 It is unclear how CIP resistance evolved in Australian poultry, but these authors speculate these isolates may have been introduced by reverse zoonosis (transmission from humans to chickens). 47
Globally, the prevalence of resistance to GEN is very low with 0%, <10%, and <5% of Campylobacter resistant in Australia, Europe, 28 and the United States, 30 respectively. Our results suggest that all six antimicrobials tested in this study are viable treatment options for campylobacteriosis in Australia. However, some of these antimicrobials (i.e., CIP) would not be suitable for treatment in some parts of Europe such as Lithuania, Portugal, and Spain where resistance is very high.
In contrast to previous reports that C. coli causes ∼9% of human Campylobacter infections in the United States 48 and 7% in England and Wales, 49 we found that C. coli accounted for 16% (27/164) of human campylobacteriosis cases in Australia. This is in line with our retail meat survey in Eastern Australia, which found C. coli in 33–56% and 58–77% of retail meat and offal products from chicken, 50 an established key risk factor for campylobacteriosis. 51 Campylobacteriosis is only notifiable to genus level in Australia, 15 thus limited data are available on the specific Campylobacter species causing disease in humans. The majority of Australian reports on human campylobacteriosis have focused solely on C. jejuni.3,11,12 It is therefore unclear whether the proportion of cases of campylobacteriosis caused by C. coli has increased over time.
Our study has some limitations. Campylobacteriosis patients in ACT, NSW, and Qld were interviewed as part of a larger case–control study, 17 and those that had travelled internationally in the 2 weeks before their illness were excluded from the study. However, we cannot rule out international or interstate travel for isolates obtained from individuals in NT, SA, Tas, Vic, or WA. Because of the small number of isolates collected from each Australian state and territory, we only had sufficient power to detect very large differences (i.e., >25% at 80% power) in the prevalence of resistance between jurisdictions; thus, regional data have been provided as Supplementary Tables S3 and S4.
To the best of our knowledge, this is the first report characterizing the status of antimicrobial resistance in both clinical C. jejuni and C. coli in Australia. Because of the low prevalence of C. coli (16%) recovered from human cases, C. jejuni and C. coli were analyzed together. However, a summary table of the Australian AST results by Campylobacter species has been included in Supplementary Table S5. Future studies should be undertaken on a larger number of C. coli isolates to determine if the prevalence of antimicrobial resistance differ between Australian C. jejuni and C. coli.
This study highlights that WGS can accurately predict phenotypic resistance to antimicrobials for which genetic mechanisms have been identified. WGS in combination with AST offers the advantage of identifying novel genetic mechanisms of resistance and the emergence of MDR isolates. Australia has some of the lowest prevalence of resistance to ERY, CIP, and TET among industrialized countries. By comparing the prevalence of antimicrobial resistance in Australia with those in Europe and the United States, we show that the prevalence of resistance vary greatly by country.
Global differences in the prevalence of antimicrobial resistance has important clinical implications as it highlights the benefit of characterizing the Campylobacter resistome in clinical cases by WGS and/or identifying where the patient travelled before treatment is prescribed. The low prevalence of ERY resistance across Australia suggests this antimicrobial remains an effective treatment option for locally acquired Campylobacter infections where antimicrobial treatment is warranted. However, surveillance monitoring of antimicrobial resistance in clinical Campylobacter species in Australia is necessary, particularly for ERY, to ensure early detection of emerging resistance.
Footnotes
Authors' Contributions
K.G. and M.D.K. conceived and designed the study. B.G.P. coordinated the study with the participating regional laboratories. D.B., A.M. and R.L.W. analyzed the data. R.L.W. drafted the article. D.B., A.M., L.V., A.V.J., M.V., J.J.S., B.G.P., K.G., and M.D.K. critically revised the article. All authors approve of the final article.
Acknowledgments
The CampySource Project Team comprises three working groups and a reference panel. The working groups focus on food and animal sampling, epidemiology and modeling, and genomics. The reference panel includes expert representatives from government and industry. The study is supported by the following partner organizations: the Australian National University, Massey University, The University of Melbourne, Queensland Health, Queensland Health Forensic and Scientific Services, New South Wales Food Authority, New South Wales Health, Hunter New England Health, Victorian Department of Health and Human Services, Food Standards Australia New Zealand, Commonwealth Department of Health and AgriFutures Australia—Chicken Meat Program. CampySource is also supported by collaboration with the following organizations: ACT Health, Sullivan Nicolaides Pathology, University of Queensland, Primary Industries and Regions South Australia, Department of Health and Human Services Tasmania, Meat and Livestock Australia, and New Zealand Ministry for Primary Industries. The CampySource Project Team consists of: Nigel P. French, Massey University, New Zealand Mary Valcanis, The University of Melbourne; Dieter Bulach, The University of Melbourne; Emily Fearnley, The Australian National University and South Australian Department for Health and Wellbeing; Russell Stafford, Queensland Health; Amy Jennison, Queensland Health; Trudy Graham, Queensland Health; Keira Glasgow, Health Protection NSW; Kirsty Hope, Health Protection NSW; Themy Saputra, NSW Food Authority; Craig Shadbolt, NSW Food Authority; Arie H. Havelaar, The University of Florida; Joy Gregory, Department of Health and Human Services, Victoria; James Flint, Hunter New England Health; Simon Firestone, The University of Melbourne; James Conlan, Food Standards Australia New Zealand; Ben Daughtry, Food Standards Australia New Zealand; James J Smith, Queensland Health; Heather Haines, Department of Health and Human Services; Sally Symes, Department of Health and Human Services, Victoria; Barbara Butow, Food Standards Australia New Zealand; Liana Varrone, The University of Queensland; Linda Selvey, The University of Queensland; Tim Sloan-Gardner, ACT Health; Deborah Denehy, ACT Health; Radomir Krsteski, ACT Health; Natasha Waters, ACT Health; Kim Lilly, Hunter New England Health; Julie Collins, Hunter New England Health; Tony Merritt, Hunter New England Health; Rod Givney, Hunter New England Health; Joanne Barfield, Hunter New England Health; Ben Howden, The University of Melbourne; Kylie Hewson, AgriFutures Australia—Chicken Meat Program; Danielle Cribb, The Australian National University; Liz Walker, The Australian National University; Rhiannon Wallace, The Australian National University; Angus McLure, The Australian National University; Ben Polkinghorne, The Australian National University; Cameron Moffatt, The Australian National University; Martyn D. Kirk, The Australian National University; and Kathryn Glass, The Australian National University. The authors thank Jutta Byrne, MDU PHL, Doherty Institute, for performing the in vitro AST.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was funded by a National Health and Medical Research Council grant (NHMRC GNT1116294), AgriFutures, Australian Government Department of Health, Food Standards Australia New Zealand, New South Wales Food Authority and Queensland Health. The National Health & Medical Research Council provided research fellowship funding for M.D.K. (APP1145997).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.