
Abstract
Background:
Hypermucoviscous carbapenem-resistant Klebsiella pneumoniae (hmCRKp) is emerging globally and approaching the worst-case scenario in health care system.
Aims:
The main objective in this study was to determine the hypermucoviscous characteristics among the carbapenem-resistant K. pneumoniae (CRKp) isolated from a teaching hospital in Malaysia. The association of hypermucoviscous phenotype with the virulence traits and clinical presentations were also investigated.
Methods:
A retrospective study was conducted in University Malaya Medical Centre (UMMC). The presence of hypermucoviscous K. pneumoniae was identified among a collection of CRKp clinical isolates (first isolate per patient) from 2014 to 2015 using string test. Correlation between clinical and microbial characteristics of the hmCRKp was investigated.
Results:
A total of nine (7.5%) hmCRKp were detected among 120 CRKp isolates. Majority of the isolates were hospital acquired or health care-associated infections. None of the patients had typical pyogenic liver abscess. All of the hmCRKp isolates harbored carbapenemase genes and were multidrug resistant. K1/K serotype, peg-344, allS, and magA were not identified among hmCRKp isolates, whereas aerobactin siderophore receptor gene (iutA), iroB, rmpA, and rmpA2 were detected. Only three hmCRKp isolates were resistant to serum bactericidal.
Conclusions:
All the isolates presented inconclusive evidence for the interpretation of hypervirulence. Therefore, more study should be performed in the future to have a better understanding of the virulence mechanisms in correlation with the clinical and microbial determinants.
Introduction
K
On the other hand, hypermucoviscosity has always been associated with hypervirulence based on the hypermucoviscous features that can be semiquantitatively defined by a “string test.” 7 However, there are increasing studies that reported the controversies between hypervirulence and hypermucoviscosity and suggested that the identification of hypervirulence could not be depending on string test.6,8,9 Due to the lack of consensus, the standard definition of hypervirulence remains ambiguous. Different combinations of criteria, including bacterial phenotype (hypermucoviscous), clinical manifestation, and virulence genes associated with hypervirulence, have been used to define hypervirulence K. pneumoniae (hvKp).6,9,10
Based on microbial characteristics, the hypermucoviscous phenotype is presented due to the overproduction of polysaccharide capsule material that is regulated by magA gene, mucoid phenotype A (rmpA), and rmpA2.11,12 Studies had shown that inhibition of transcription of rmpA and rmpA2 gene that is repressed by holo-fumarate nitrate reduction regulator plays a role in capsular polysaccharide production and virulence such as serum resistance and antiphagocytosis. 13 Isolates that phenotypically showed hypervirulent/hypermucoviscous characteristics without magA, rmpA, and rmpA2 genes have also been described. 14 Molecular epidemiology studies have successfully identified several important hypervirulence-associated genes in K. pneumoniae such as capsular serotype K1 and K2, a combination of iron acquisition system (enterobactin gene [entB], hydroxamate siderophore gene [iutA], yersiniabactin gene [ybtS] and iron transport/phosphotransferase function [kfu]), allantoin metabolism-associated gene (allS), and fimbrial and nonfimbrial adhesion genes such as mrkD, ycfM, and KPN. 15 Furthermore, putative transporter (peg-344) and salmochelin siderophore biosynthesis (iroB) have been shown experimentally with high accuracy in identifying hvKp isolates. 16
Most of the hvKp isolates are known to be multidrug susceptible. Nevertheless, multidrug-resistant hvKp, including ESBL and carbapenemase-producing hvKp isolates, had also been reported.17–19 Treatment and management of hvKp infection are difficult and associated with high morbidity and mortality.18,20 In fact, there is increasing evidence showing that hvKp is associated with other infections such as bacteremia, pneumonia, urinary tract infection, biliary tract infection, and soft tissue infection.
In this study, we aimed to determine the hypermucoviscous characteristics among the carbapenem-resistant K. pneumoniae (CRKp) isolated from a teaching hospital in Malaysia. The association of hypermucoviscous phenotype with the presence of hypervirulence genes and clinical presentations was also determined.
Materials and Methods
Ethics statement
The approval to collect clinical isolates and to access the clinical/patients' data was obtained from University of Malaya Medical Centre (UMMC) Medical Ethics Committee in 2015 (MECID.NO: 20154-1249).
Hospital setting, study period, and collection of isolates
Strain collection was conducted in UMMC, a tertiary teaching hospital with 1,617 beds located in Klang Valley, Malaysia. All K. pneumoniae clinical cultures identified as carbapenem resistant using VITEK II system and confirmed by E-test in the Medical Microbiology Diagnostic Laboratory (MMDL) of UMMC between years 2014 to 2015 were prospectively collected. All isolates were the first CRKp isolated from the patients' samples, but in some of the samples, the isolate may has been co-isolated with another organisms.
Selection of hypermucoviscous K. pneumoniae from the CRKp collection
Hypermucoviscous K. pneumoniae was defined by a positive string test, based on the formation of a viscous filament ≥5 mm when a bacteriology inoculation loop is stretching a K. pneumoniae bacterial colony on blood agar plate. 14 The antimicrobial susceptibility profile and clinical details at the time of the hypermucoviscous carbapenem-resistant K. pneumoniae (hmCRKp) isolation were obtained from the MMDL and patient's case note, respectively.
Molecular detection of carbapenemase genes
Polymerase chain reaction (PCR) was conducted to detect the presence of carbapenemase genes using primers and conditions published previously: blaKPC, 21 blaNDM, 22 blaVIM, 23 blaIMP, 23 and blaOXA. 24 In brief, for blaKPC amplification, each 25 μL reaction consisted the following components: 15.4 μL of ultrapure water, 5 μL of 5 × buffer, 1 μL of 25 mM of MgCl2, 0.5 μL each of forward and reverse primer, 0.3 μL of 10 mM of PCR nucleotide mix, 0.3 μL of Taq polymerase, and 2 μL of bacterial DNA templete. For blaNDM, blaVIM, blaIMP, and blaOXA amplifications, each 25 μL reaction consisted the following components: 15.1 μL of ultrapure water, 5 μL of 5 × buffer, 1 μL of 25 mM of MgCl2, 0.75 μL each of forward and reverse primer, 0.2 μL of 10 mM of PCR nucleotide mix, 0.2 μL of Taq polymerase, and 2 μL of bacterial DNA templete. The PCR cycling conditions were as follows: 94°C for 3 min; 35 cycles of 1 min at 94°C, 1 min at AT (annealing temperature), and 2 min at 72°C; and 72°C at 7 min. The PCR products were analyzed by agarose gel electrophoresis.
Molecular detection of porin genes
PCR was conducted to screen for the presence of porin genes using primers and conditions published previously (ompK 35, ompK 36, and ompK 37). 25 Each 25 μL reaction consisted the following components: 15.1 μL of ultrapure water, 5 μL of 5 × buffer, 1 μL of 25 mM of MgCl2, 0.75 μL each of forward and reverse primer, 0.2 μL of 10 mM of PCR nucleotide mix, 0.2 μL of Taq polymerase, and 2 μL of bacterial DNA templete. The PCR cycling conditions were as follows: 94°C for 3 min; 35 cycles of 1 min at 94°C, 1 min at AT, and 2 min at 72°C; and 72°C at 7 min. The PCR products were analyzed by using agarose gel electrophoresis.
Detection of virulence-associated genes
All the CRKp isolates that demonstrated hypermucoviscous characteristics and 10 randomly selected non-hypermucoviscous CRKp (non-hmCRKp) isolates were subjected to virulence gene detection. The detection of virulence genes such as ybtS, mrkD, entB, rmpA, K2, kfu, allS, iutA, and magA was performed as previously described by Compain et al. 15 The presence of peg-344, rmpA2, and iroB genes that previously demonstrated high accuracy in detecting hvKp was determined as described by Russo et al. 16 The amplicons of rmpA gene from hmCRKp-27 and hmCRKp-48 were purified and subjected to Sanger sequencing. The sequences were aligned with NTUH-K2044 (GenBank Accession No. AB289642) as the reference strain using MEGA 7.0 software 26 for point mutation determination.
Genotypic characterization of hmCRKp
All hmCRKp isolates were subjected to multilocus sequence typing (MLST) analysis using seven housekeeping genes (gapA, infB, mdh, pgi, phoE, rpoB, and tonB) according to Institut Pasteur's MLST scheme at https://bigsdb.pasteur.fr/klebsiella/primers_used.html The sequence types (STs) were assigned at https://bigsdb.pasteur.fr/cgi-bin/bigsdb/bigsdb.pl?db=pubmlst_klebsiella_seqdef
Clonal complex assignment and clonal relatedness
Clonal relationship of all the hmCRKp isolates was evaluated using eBURST v 3.0 at http://eburst.mlst.net/. Clonal complexes were defined as groups of STs sharing at least six of seven MLST alleles.
Serum bactericidal assay
The virulence of hmCRKp and non-hmCRKp isolates was determined by serum bactericidal assay as described by Podschun et al.
27
Bacterial culture was grown to mid-log phase and diluted to 106 cells/mL using normal saline. The bacterial suspension was incubated at 37°C in microtiter trays with normal human serum in 1:3 ratio. The viability of bacteria after 1-, 2-, and 3-hr incubation was determined using serial dilution and plating method. Each bacterial isolate was tested in three replicates. Each bacterial isolate was graded as highly sensitive (grade 1 and 2), intermediate sensitive (grade 3 and 4), and resistant (grade 5 and 6):
Grade 1: viable counts at 1 and 2 hr were <10% of the inoculum, at 3 hr <0.1% Grade 2: viable counts at 1 hr 10–100%, at 3 hr <10% Grade 3: viable counts at 1 hr >100%, at 2 and 3 hr <100% Grade 4: viable counts at 1 and 2 hr >100%, at 3 hr <100% Grade 5: viable counts at 1, 2, and 3 hr >100%, but decreasing during 3 hr Grade 6: viable counts at 1, 2, and 3 hr >100% and increasing throughout 3 hr
Results
Sample origin and isolates' identity
Among the 120 CRKp isolates collected from UMMC from year of 2014 to 2015, nine (7.5%) were string-test positive and identified as hmCRKp. These isolates were revived from tracheal secretion (n = 3, 25%) and one isolate each was isolated from urine, blood, sputum, pus (R) thigh, swab from peritoneal fluid, and tissue, respectively. In five of the nine samples with hmCRKp isolated, other bacteria have also been isolated.
Demographic and clinical characteristics of patients at the time of the hmCRKp isolation
Among the nine hmCRKp isolates, five (55.6%) were associated with hospital-acquired infection, one (11.1%) was identified as health care-associated infection, and three (33.3%) were clinically regarded as colonizers. A total of six (66.7%) hmCRKp were isolated from patients admitted to ICU with different clinical diagnosis, such as ventilator-associated pneumonia (n = 2), intra-abdominal sepsis secondary to ischemic perforated bowel (n = 1), bacteremia secondary to infected sacral sore/urosepsis (n = 1), septic shock secondary to infected sacral sore (n = 1), and sepsis secondary to disseminated abscesses (n = 1). The in-hospital mortality among the ICU hmCRKp cases was 50% (n = 3) (Table 1).
Clinical Cases of Patients from Whom hmCRKp Was Isolated
Colonization = isolation of the microorganism from any nonsterile body site (usually rectum or perianal area, and also respiratory tract/oral cavity, vagina, skin, and urine) in the absence of clinical findings of infection. 42
Hospital-acquired infection = Infection with positive culture obtained from patients already hospitalized for 48 hr or longer. 34
Health care-associated infection = Infection with positive culture obtained at admission or within 48 hr and if the patient had received intravenous therapy at home, had attended a hospital or hemodialysis clinic, or had received intravenous chemotherapy or was hospitalized in the preceding 3 months, or residing in a long-term care facility. 34
Empiric antibiotic = Antibiotic that is given to the patients with sign and symptoms of infection before the identification of organisms and susceptibility data were available.
Targeted antibiotic = Antibiotic that is given to the patients after the culture and susceptibility data are available.
Invasive device = Medical device that is introduced into the body, either through a break in the skin or body orifice. This includes central venous line, peripheral line, peripherally inserted central catheter, nasogastric tube, catheter bladder drainage, internal jugular catheter, and external ventricular drain.
Invasive procedure = One where access to the body is gained through an incision, percutaneous puncture, where instrumentation is used in addition to the puncture needle, or instrumentation through a natural orifice. It begins when entry to the body is gained and ends when the instrument is removed, and/or the skin is closed. Invasive procedures are performed by trained health care professionals using instruments, which include, but are not limited to, endoscopes, catheters, scalpels, scissors, devices, and tubes. 43
Underlying disorder = Acute or chronic comorbidity or condition not related to sepsis that alters short- and/or long-term survival of infectious diseases. 44
Clinical diagnosis = Diagnosis of the site infection at the time of CRKp isolation. As the outcome measure was all-cause mortality during current admission.
The patient had also isolated CRKp+carbapenem-sensitive E. coli from tissue (R) thigh, peritoneal pus, and peritoneal fluid samples received on the same day with the pus (R) thigh, but the CRKp from these samples had not been stocked or included in the study.
CRKp, carbapenem-resistant K. pneumonia; hmCRKp, hypermucoviscous carbapenem-resistant K. pneumonia; MDR, multidrug resistant; N/A, not applicable.
Antimicrobial susceptibility profile
All hmCRKp isolates were resistant to ampicillin, ampicillin-sulbactam, amoxicillin-clavulanic acid, ceftazidime, ceftriaxone, cefotaxime, cefuroxime, cefepime, cefoxitin, and piperacillin-tazobactam. Eight hmCRKp isolates were resistant to meropenem and one isolate was intermediate to meropenem. Nevertheless, 88.9% (n = 8) of isolates were susceptible to trimethoprim-sulfamethoxazole (Fig. 1).
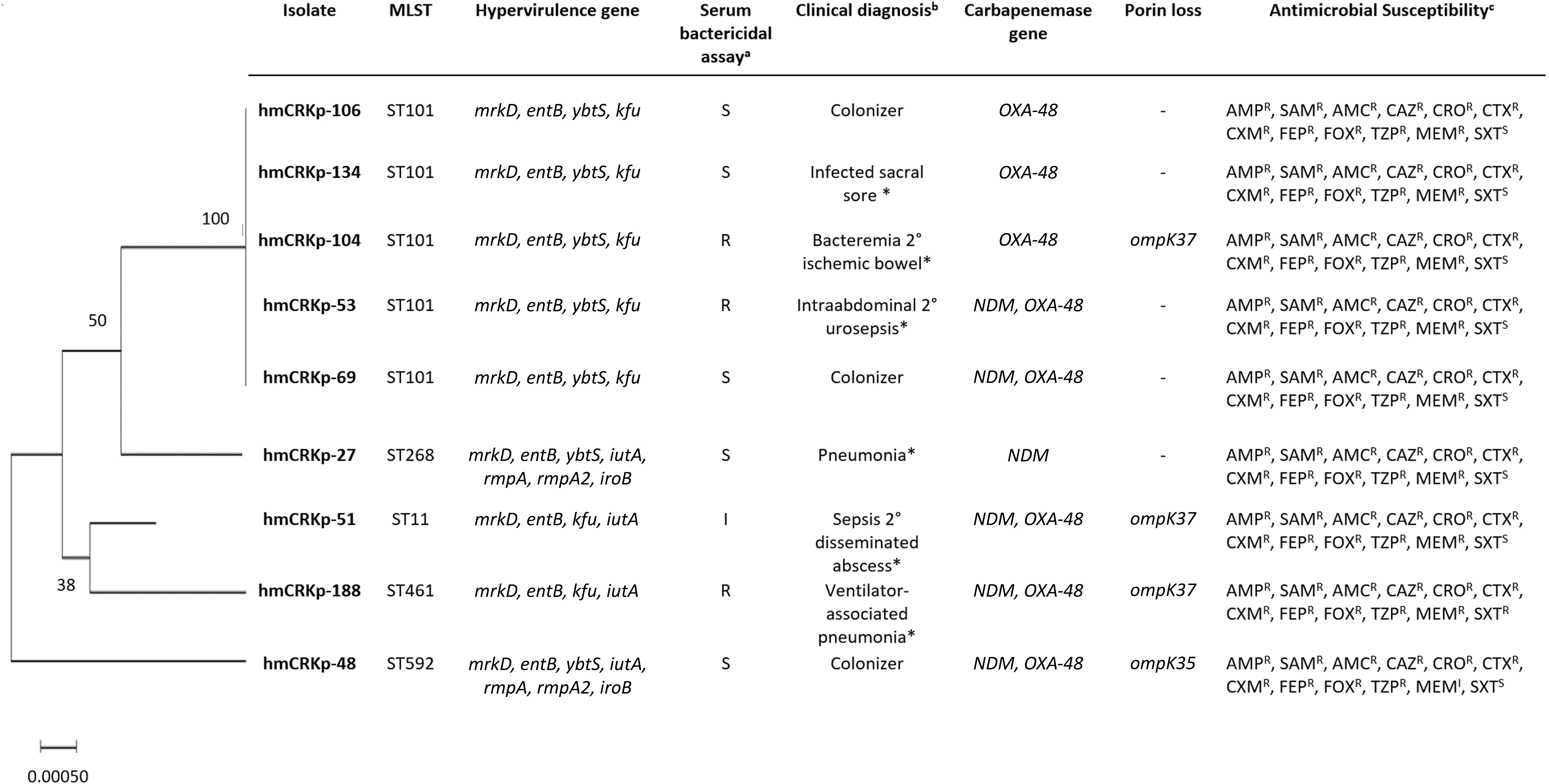
Maximum-likelihood dendrogram based on MLST for the hmCRKP isolates with the genotypic, phenotypic, and clinical characteristics. aThe results obtained for serum bactericidal assay were categorized as S (sensitive = grade 1 and grade 2), I (intermediate = grade 3 and grade 4), and R (resistant = grade 5 and grade 6). bClinical presentation of the cases with * indicating hospital-acquired infection. cTwelve antimicrobial agents were tested: AMP (ampicillin), SAM (ampicillin-sulbactam), AMC (amoxicillin-clavulanic acid), CAZ (ceftazidime), CRO (ceftriaxone), CTX (cefotaxime), CXM (cefuroxime), FEP (cefepime), FOX (cefoxitin), TZP (piperacillin-tazobactam), MEM (meropenem), and SXT (trimethoprim-sulfamethoxazole). The antibiotic susceptibility was indicated (I, intermediate; R, resistant; S, sensitive). hmCRKp, hypermucoviscous carbapenem-resistant K. pneumonia; MLST, multilocus sequence typing.
Carbapenem resistance genes and loss of porins
Out of the nine hmCRKp isolates, four harbored only a single carbapenemase gene (blaOXA-48, n = 3 and blaNDM, n = 1). Five isolates were blaOXA-48 and blaNDM dual carbapenemase producers. The most common carbapenemase gene detected was blaOXA-48 (n = 8, 88.9%), followed by blaNDM (n = 6, 66.7%). blaVIM, blaIMP, and blaKPC were not detected among hmCRKp isolates. ompK 36 was detected in all hmCRKp isolates; however, four isolates lost either ompK 35 or ompK 37 (Fig. 1).
Comparison of hypervirulence-associated genes among hmCRKp and non-hmCRKp
All the hmCRKp isolates (n = 9, 100%) and 10 non-hmCRKp (66.7%) in this study harbored at least 2 types of siderophore genes reported by Compain et al. 15 Among the 12 targets, iutA, iroB, rmpA, and rmpA2 were only detected in hmCRKp isolates (n = 2, 22%), while peg-344, allS, magA, and K2 were not detected from any isolate regardless of the hypermucoviscous characteristics. Four genes were present in both hmCRKp and non-hmCRKp (ybtS, entB, mrkD, and kfu). Based on the sequencing results, no mutation was found in the rmpA gene that was present in the two hmCRKp isolates (Table 2).
The Presence of Hypervirulence Genes and Serum Bactericidal Ability in hmCRKp and Non-hmCRKp Strains That Showed Different Clinical Charactristics (Infection/Colonization)
MLST and phylogenetic analysis
A total of five STs were observed among the studied isolates. No new ST was found in this study. The predominant ST identified was ST101 (n = 5), while the other four isolates were assigned as ST268, ST11, ST461, and ST592, respectively. A maximum likelihood tree has been generated based on the concatenated sequences of the seven genes in the order of gapA-infB-mdh-pgi-phoE-rpoB-tonB. ST298 was 50% similar to ST101, while ST592 were distinctly different from all isolates (Fig. 1). eBURST analysis of the STs in our study and other STs on the PubMLST database revealed that only ST268 was grouped with ST234 and ST36 as clonal complex (Fig. 2).

Comparative eBURST analysis showing the clonal assignment of our study isolates compared to the isolates present in Klebsiella pneumoniae MLST database.
Resistance of serum bactericidal among hmCRKp and non-hmCRKp
A total of three hmCRKp isolates were resistant to serum bactericidal where the highest serum resistance level (grade 6) was observed in one isolate. One hmCRKp isolate was intermediate, while the other five hmCRKp and all non-hmCRKp isolates were susceptible to serum killing (Table 2).
Discussion
To our best knowledge, this is the first report of carbapenem-resistant hmKp in Malaysia. In our study, 9 hmCRKp isolates from 9 patients were successfully identified from a collection of 120 CRKp isolates over a 2-year period. The low prevalence of CRKp isolates that showed hypermucoviscous characteristics based on string test in this study concurred with the finding of Yao et al. 28 where only four hmCRKp isolates were identified within a 4-year period. On the other hand, although Li et al. 29 reported high prevalence (62.1%) of hvKp from community-acquired infections, none was carbapenem resistant.
It is believed that hvKp isolates are more virulent and always cause PLA. 30 However, these phenomena have not been observed in our study. In contrast, our study presented similar findings with Yao et al., where no significant clinical characteristic was found among the string test-positive isolates compared to those non-hypermucoviscous. In our study, among six patients with hmCRKp infections, two patients had hospital-acquired ventilator-associated pneumonia, while others presented intra-abdominal sepsis secondary to ischemic perforated bowel, bacteremia secondary to infected sacral sore/urosepsis, sepsis secondary to disseminated abscesses, and a septic shock secondary to infected sacral sore. In fact, hvKp isolates have been associated with other infections such as bacteremia, pneumonia, urinary tract infection, biliary tract infection, and soft tissue infection.4–6
Throughout the year, several genetic determinants have been identified as important virulence factors. Previously study by Bachman et al. 31 reported the enhanced growth of K. pneumoniae co-harboring entB and ybtS in the lungs of a murine model and postulated that acquisition of both siderophores is able to increase virulence among hvKp. 31 Besides, mrkD gene, a type 3 fimbrial adhesion, is known to be playing an important role in adherence to cell surfaces before progressing to infection and promotes biofilm formation, 32 while kfu, which mediates the uptake of ferric iron, has also been reported in many hypervirulent isolates.16,33 Although the mentioned genes have been strongly associated with hypervirulence phenotype, our study was in agreement with the previous reports where ybtS, entB, mrkD, and kfu genes might not be the ultimate determinants for hypervirulent as these genes were also being detected in non-hmCRKp isolates.34–36 In addition, unique clinical presentation associated with hypervirulence (PLA) was also not being observed among the non-hmCRKp isolates with the mentioned virulence genes. Based on the clinical records, these non-hmCRKp isolates in our study were associated with bacteremia, urinary tract infection, wound infection, and right hypochondrial abscesses.
Among the hmCRKp isolates, two isolates that harbored iutA, iroB, rmpA, and rmpA2 genes were detected. These two isolates were identified as ST268 and ST592, respectively, which have not been associated with K1 and K2 serotypes (Fig. 1). 37 Based on the literature review, ST268 mainly comprised K20 isolate, while ST592 was composed of K57 isolates.34,38 Our study is also in agreement with Guo et al. 34 where rmpA was present in the K20 and K57 isolates with hypermucoviscous phenotype.
The serum bactericidal assay showed that only three hmCRKp in this study were resistant and one was intermediate to serum killing. Resistant to human serum is recognized as an important virulence factor to evade human first line of defense and has been correlated with the onset of infection and severity of symptoms. 39 Study had shown that hmKp with one or two hypermucoviscous-associated genes (magA or/and rmpA) had the highest serum resistance level compared to non-hmKp (without magA or/and rmpA). 40 However, in our study, hmCRKp isolates with rmpA gene were susceptible to serum killing and concurred with the findings of Loraine et al. 41 Hsu et al. 42 also reported that although rmpA was shown to enhance capsular polysaccharide production and mucoviscosity, this gene did not play any role in serum resistance or proliferation in nonimmune human serum.
Previous study suggested a close relationship between CC23 and hvKp with serotype K1 using MLST typing. 3 CC23 hvKp was prevalent among the studies conducted in China and South Korea.34,43 Whole-genome sequencing analysis suggested that CC23 may exhibit multiple international transmission 3 ; however, in our study, we did not detect any STs of CC23. It is also noteworthy that all STs identified in our study were not the prevalent STs such as ST65 like and ST86 like, which are highly associated to K2 serotype hvKp. The dominant ST identified among the isolates of our study was ST101, which is less commonly reported serotype for hvKp 44 and suggested an independent local establishment of this ST. In fact, ST101 has been the dominant ST among the CRKp in our hospital since 201345 and it is important to assess the development of resistance and virulence among this important bacterial population. However, due to the low prevalence of hmCRKp, there was no significant correlation between different variables such as hypermucoviscous characteristics, virulence genes, serum resistance, and clinical presentation. Therefore, more CRKp should be tested in the future to have a better understanding of the epidemiology of hmCRKP and to improve the definition of hypervirulent among the clinical K. pneumoniae isolates.
Conclusions
In conclusion, nine hmCRKp isolates had been identified among the CRKp isolates in our hospital based on positive string test, but none of these isolates caused the typical PLA as previously reported. Comparison of the virulence profiles and serum bactericidal between hmCRKp and non-hmCRKp was performed. Although specific virulence genes were only detected in hmCRKp, isolates of both categories showed high level of resistance in serum bactericidal. Hence, continuous efforts are needed in assessing the sensitivity and specificity of these criteria in identification of hypervirulent variants to improve current clinical diagnostic and treatment strategy.
Footnotes
Acknowledgments
The authors would like to thank University of Malaya and UMMC for facilities and utilities. We would like to thank Professor Sanjay Rampal for his professional advice on data presentation. We would like to thank my PhD scholarship for funding support.
Disclosure Statement
No competing financial interests exist.
Ethics Approval
The research ethical approval was obtained from UMMC Medical Ethics Committee in 2015 (MECID.NO: 20154-1249) and had conformed to the principles embodied in the Declaration of Helsinki.
Funding Information
This work was supported by International Society of Infectious Diseases [IF016-2015] and Fundamental Research Grant Scheme [FP013-2018A].