
Abstract
Aim:
In Algeria, colistin is used as a metaphylactic treatment in the poultry industry for the treatment of Gram-negative gastrointestinal infections and also as a feed additive to promote animal growth. The aim of this study was to investigate the importance and genetic characteristics of colistin-resistant Enterobacterales from chicken meat in Western Algeria.
Results:
A total of 181 samples of chicken meat were collected from three poultry farms across three provinces in Western Algeria. The presence of colistin-resistant Enterobacterales isolates was screened on selective media. Resistance and virulence profiles were characterised by PCR and sequencing. The clonal relatedness of the different mcr positive isolates was studied using repetitive sequence-based PCR (Rep-PCR) and multilocus sequence typing. Transferability and characteristics of plasmids harboring mcr-1 positive gene were performed using conjugation, PCR-based replicon typing, and whole-genome sequencing. A total of 22 isolates with acquired colistin resistance were identified giving an overall prevalence of 12.2% (22/181): 17 Escherichia coli (predominantly ST224 [n = 4, 23.5%]) and 5 Klebsiella pneumoniae (ST17 [n = 2, 40%], ST646 [n = 2, 40%], and ST944 [n = 1, 20%]). mcr-1 gene was exclusively found in 11 E. coli (prevalence of 6.1% [11/181]) and was associated with IncFV (n = 7) and IncFIIK (n = 4) plasmids. All the isolates had a commensal origin (n = 11). One isolate harbored virulence profile, a high colistin resistance (minimum inhibitory concentration = 96 mg/L), with some new mutations in the chromosomic colistin-resistant genes and different pathogenicity islands typically identified in uropathogenic E. coli.
Conclusions:
This study reports the diffusion of mcr-1 producing Enterobacterales from chicken meat in Western Algeria. This represents a worrisome situation needing continuous monitoring.
Introduction
Recently, colistin is highly reconsidered as last resort antibiotic to treat infections caused by multidrug resistant (MDR) or extremely drug resistant Gram-negative bacteria (GNB) in human therapy. 1 This antibiotic has also been prescribed in veterinary medicine for many years and is still widely consumed. 2 In animal farming, predominantly in pig, poultry, and cattle production, it has been used to treat gastrointestinal infections and for nontherapeutic goals (mainly for prevention, metaphylaxis, and growth promotion in some countries). 3 In poultry production, the main indication of colistin, administered orally in feed and drinking water, is the treatment of colibacillosis. 3 In African countries, injudicious purchase of the antimicrobial drug by farmers without prescription has led to improper use and very high consumption. 4 Unfortunately, the wide consumption of colistin is considered to be the source of the emergence of colistin resistance in GNB with increasing number of outbreaks reported in humans and animals worldwide. 2
Acquired resistance in colistin emerges by the addition of a positive cation (e.g., modification of lipid A by addition of phosphoethanolamine or 4-amino-4-deoxy-
Studies on the mcr-1 gene revealed that this gene has been circulating for many years, and food-producing animals could represent its main reservoir. 6 Pathogens carrying mcr genes can be transmitted to humans through consumption of undercooked food or noncompliance with hygiene practices especially in Algeria where the surveillance and monitoring of colistin use in poultry seem to be absent. 2 The uncontrolled administration of colistin in poultry was suggested to be the source of the potential spread of the mcr-1 gene, which was detected in bacteria present in feces of chickens in East Algeria.7,8 These studies mandate prompt investigation of the possibility of food-producing animals to be one of the major source of human contamination by MDR bacteria.
The aim of our study was to evaluate the occurrence of colistin-resistant Enterobacterales isolated from chicken meat in Western Algeria, to evaluate the presence of clonal strains and to characterize the genetic support of this resistance.
Materials and Methods
Sampling and microbiological procedures
All the experiements were approved by the Algerian national veterinary authority (authorization number: AVN 13051). From February 2017 to July 2017, we randomly collected (by a random draw) 181 chicken (6 weeks of age) from the three main poultry public and legal farms localized in three cities in Western Algeria (Oran, Tiaret, and Saida) and containing 4,000, 3,500, and 3,000 chickens, respectively (Fig. 1). Six-week-old chickens were randomly selected. They were slaughtered by bleeding (performed by the cutting of carotids) by a trained veterinary in a sterile condition at room temperature. Neither incision at the buttonhole nor evisceration was performed to avoid gut contamination. The thigh muscle section was then performed with a sterile blade; the first piece of flesh incised at the surface was removed, and the second deep piece was collected and directly placed in a sterile vial containing physiological waer. Samples were taken and immediately transported at +4°C to the laboratory of microbiology at the National Institute of Veterinarian Sciences (Tiaret, Algeria) for analysis. The meat was crushed with a stomacher homogenizer, and 25 g of each sample was cultured in 225 mL of Luria Bertani broth (Pasteur Institute, Algiers) supplemented with 32 mg/L of vancomycin (to avoid Gram–positive bacteria contamination) and incubated overnight at 37°C. Following incubation, 200 μL of each culture was enriched into MacConkey broth (Fluka, St. Louis) supplemented with 2 μg/mL colistin and incubated for 18–24 hr at 37°C.
9
For the selection of colistin-resistant (col-R) isolates, 200 μL of each culture was then plated on selective chromogenic media CHROMID® Colistin R agar (bioMérieux, Marcy l'Etoile, France). The identification of the col-R isolates was performed by the Vitek2® and VITEK

Area sampling and distribution of colistin resistant Enterobacterales isolates in chicken meat in different cities of Western Algeria.
Antibiotic susceptibility testing
For each isolate, susceptibility to antimicrobial agents was tested by the disk diffusion method (Bio-Rad, Marnes La Coquette, France) on Mueller-Hinton agar plates according to recommendations of the European Committee on Antimicrobial Susceptibility Testing (EUCAST, 2018). The minimum inhibitory concentration (MIC) of isolates to colistin was determined by microbroth dilution (Umic®; Biocentric, Bandol, France) over a range of dilutions (0.12–128 mg/L). All experiments were performed twice independently, and Escherichia coli ATCC 25922 was used as a control strain. Susceptibility patterns were interpreted according to the EUCAST 2018 guidelines. Enterobacterales isolates with colistin MICs ≥2 mg/L were considered as colistin resistant.
Molecular genotyping and characterization of colistin-resistant isolates
Total DNA was extracted using the EZ1 DNA Tissue Kit on the BioRobot EZ1 extraction platform (Qiagen, Courtaboeuf, France). All isolates were screened for the presence of acquired colistin genes (mcr-1 to mcr-9). 10 Genes encoding penicillinases and extended-spectrum β-lactamases (ESBLs) (blaTEM, blaSHV, and blaCTX-M) were detected by PCR using specific primers and confirmed by sequencing the PCR products, as described previously.11,12 Sulfonamide resistance genes (sul-1, sul-2, and sul-3) and plasmid-mediated quinolone resistance (PMQR) encoding genes [qnrA, qnrB, qnrS, qepA, oqxAB, and aac(6′)-Ib-cr] were screened as previously described.11,12 Screening of chromosomal mutations in the mgrB, pmrAB, phoPQ, and acrAB genes in col-R Enterobacterales isolates was performed by PCR using specific primers and confirmed by sequencing. 1 Klebsiella pneumoniae ATCC43816 and E. coli K12 were used as control.
The clonal relatedness of the col-R isolates was evaluated using repetitive sequence-based PCR (rep-PCR) and multilocus sequencing type (MLST). Rep-PCR was performed on the DiversiLab® System (bioMérieux) following the manufacturer's instructions. Isolates with identical strain patterns were considered indistinguishable if the similarity percentage was ≥95%. MLST analysis was performed using the Pasteur Institute's MLST scheme for K. pneumoniae (bigsdb.web.pasteur.fr) and Achtman MLST for E. coli.
Whole genomic sequencing
The whole-genomic sequencing of MCR-1-producing E. coli (A50) with colistin MIC of 96 mg/L was performed on the Illumina genome analyzer IIX system by GenoScreen SA (Lille, France). The quality and potential defect of tested sequences were estimated using Fastqc Software (version 0.11.7). Reads were assembled using SPAdes v3.10.1, and assemblies were annotated using BOWTIE2. R Software and packages of Bioconductor (Biostrings and ShortRead) were used for querying the assemblies. Sequence reads were trimmed and filtered according to quality criteria and mapped against E. coli K12 genome (GCF_000005845.2).
Plasmid characterization
Plasmid DNA was extracted from the isolates using the Plasmid Miniprep Kit (QIAGEN). Conjugation experiments were performed using a broth mating method as described previously 5 : MCR-1 producers served as donor and E. coli J53 rifampicin-resistant (RifR) as a recipient. Transconjugants were selected on Luria-Bertani agar plates supplemented with rifampicin (256 mg/L) and colistin (2 mg/L). Transconjugants were confirmed by antimicrobial susceptibility and PCR detection of the colistin resistance (mcr-1) gene performed on plasmid extract. Replicon typing of the isolated plasmid DNA was done by a PCR-based replicon-typing method as described previously. 13
Plasmid from transconjugant Tc-A50* was extracted for sequencing and to evaluate the genetic environment of the mcr-1 gene in the E. coli A50. The experiment was performed on the Illumina genome analyzer IIX system by GenoScreen SA as previously described. 12
Phylogenetic groups and virulence genes
For E. coli isolates, PCRs were performed to determine the phylogenetic groups (A, B1, B2, C, D, E, F, and clade I) as previously described. 14 The E. coli isolates were tested by PCR for the presence of a panel of 20 virulence factors (VFs)-encoding genes: fimH, papG1, papG2, papG3, papAH, papC, papEF, sfaS, focG, afa/draBC (fimbriae and/or adhesins); cnf1 and hlyA (toxins); iutA, fuyA, and iroN (iron uptake); kpsMTI, kpsMTII, and traT (protectins); malX, and irp2. 12 Furthermore, E. coli isolates were also investigated for the presence of virulence genes (iss, iucD, tsh, ompT, cva/cvi, and astA) associated with colibacillosis and ireA, an iron-regulated gene present in avian pathogenic E. coli. 15
For K. pneumoniae isolates, the presence of VF associated genes was assessed by PCR assays 12 : fimH, mrkD, and cf29a corresponding to adhesin encoding genes; ureA corresponding to toxin encoding gene; entB, ybtS, kfu, and iutA corresponding to iron uptake encoding genes; magA, rmpA, kpsMTII corresponding to protectin encoding genes, with the genes of the capsular serotypes K5, K57, K54, and K20; and allS, uge, and wabG.
GenBank accession numbers
The whole-genome sequence of E. coli A50 isolate was accessible in GenBank accession no. PRJNA558906. The complete sequence of the pA50 plasmid carrying the mcr-1 gene has been deposited at The DNA Data Bank of Japan/The European Molecular Biology Laboratory/GenBank under the accession number SAMN12501891.
Results
Prevalence of col-R Enterobacterales in chicken meat and antibiotic susceptibility
Of 181 chicken meat samples collected, 285 isolates were identified as resistant to colistin (MIC ≥2 mg/L). The isolates were identified as Proteus sp. (n = 77, 27.0%), Morganella spp. (n = 68, 23.9%), Serratia spp. (n = 60, 21.0%), Providencia spp. (n = 58, 20.4%), E. coli (n = 17, 6%), and K. pneumoniae (n = 5, 1.7%). As the four first species are naturally resistant to colistin, we focused our research on the 22 remaining isolates dectected in 22 samples, giving an overall prevalence of 12.2% (22/181) (Table 1).
Main Characteristics of Colistin-R Enterobacterales Isolated from Chicken Meat in Western Algeria
AMR, antimicrobial resistance; PL, Phylotypes; AMX, amoxicillin; TIC, ticarcillin; PIL, piperacillin; CXN, cefalexin; NAL, nalidixic acid; OFX, ofloxacin; CIP, ciprofloxacin; SXT, cotrimoxazole; COL, colistin; MIC, minimum inhibitory concentration; MLST, multilocus sequencing type; NA, not applicable.
Among these col-R isolates, total resistance to nalidixic acid, ofloxacin, and ciprofloxacin was noted (n = 22, 100%), followed by cotrimoxazole (n = 21, 95.5%). Resistance to amoxicillin, ticarcillin, and piperacillin was observed in 11 E. coli isolates (50%). All the isolates were susceptible to carbapenems, cefotaxime, ceftazidime, cefepime, fosfomycin, and aminoglycosides. Interestingly, MICs of colistin varied between 2 and 32 mg/L for E. coli (except A50 isolate which exhibited a high MIC of colistin 96 mg/L) and 4 and 256 mg/L for K. pneumoniae (Table 1).
Molecular characterization of col-R isolates
Among the 17 col-R E. coli, 11 isolates (64.7%) harbored the mcr-1 gene, giving a prevalence of 6.1% (11/181) (Table 1); no mcr genes were detected among col-R K. pneumoniae. All mcr-1 encoding E. coli isolates were recovered from three farms: Oran, n = 5 (45.5%); Tiaret, n = 4 (36.4%); and Saida, n = 2 (18.2%) (Table 1; Fig. 1).
Among six col-R mcr negative E. coli isolates, PCR and sequencing of the chromosomal genes showed mutations in different genes as follows: phoP (V108D), phoQ (S138D [n = 6], L96S [n = 3], L96A [n = 2], and L87P [n = 1]), pmrA (R81S [n = 4] and G144S [n = 2]), and pmrB (R256G [n = 4], Y315F [n = 1], and D292N [n = 1]). Likewise, all mcr negative K. pneumoniae harbored chromosomal mutations in phoP (L87P [n = 3], A30S [n = 2]), pmrA (S42N [n = 4], G53C [n = 4], E57G [n = 1]), and pmrB (T1570 [n = 3], Q56S [n = 2], and R256G [n = 2]). We identified some new mutations never previously described in K. pneumoniae isolates in pmrB (G20S) and phoQ (A70K, D90H/N, P72N, D74E, Q92S, I75L, E77D, and K94E) genes (Table 2).
Chromosomal Mutations and Amino Acid Deletions Responsible for Acquired Colistin Resistance in Enterobacterales Strains Isolated from Chicken Meat in Western Algeria
In bold, new mutations.
ND, not determined; ST, sequence type.
The E. coli A50 isolate possessed both the mcr-1 gene and novel mutations in differently acquired colistin resistance encoding genes: deletion of I363 in pmrB gene, a silent mutation in phoP gene (468T > C), and diverse mutations in qseC and qseB (Supplementary Table S1).
Coexistence of ESBL and PMQR genes was not detected but 10 of the 17 col-R E. coli isolates (58.8%) harbored blaTEM −1 gene. Among them, E. coli A50 had both blaTEM −1 and blaSHV −1 genes (Table 1). Finally, 21 of the 22 col-R Enterobacterales isolates (95.5%) harbored a sulfonamide resistant gene with sul1 (n = 21), sul3 (n = 9), and sul2 (n = 8) genes (Table 1).
Clonality of the col-R isolates
The Rep-PCR revealed distinct lineages among the 17 col-R E. coli isolates with a total of nine different patterns detected in mcr-1-positive isolates and three in mcr-negative isolates (Fig. 2). Three isolates belonged to pattern A, corresponding to mcr-1-positive E. coli isolated from Oran. The remaining patterns contained two (three Rep-PCR patterns) or single isolates (eight Rep-PCR patterns). The Achtman MLST confirmed the diversity of these isolates with only two sequence types (STs, ST224 and ST168) found in three and two isolates, respectively. The other 12 STs were distinct (Table 1; Fig. 2).
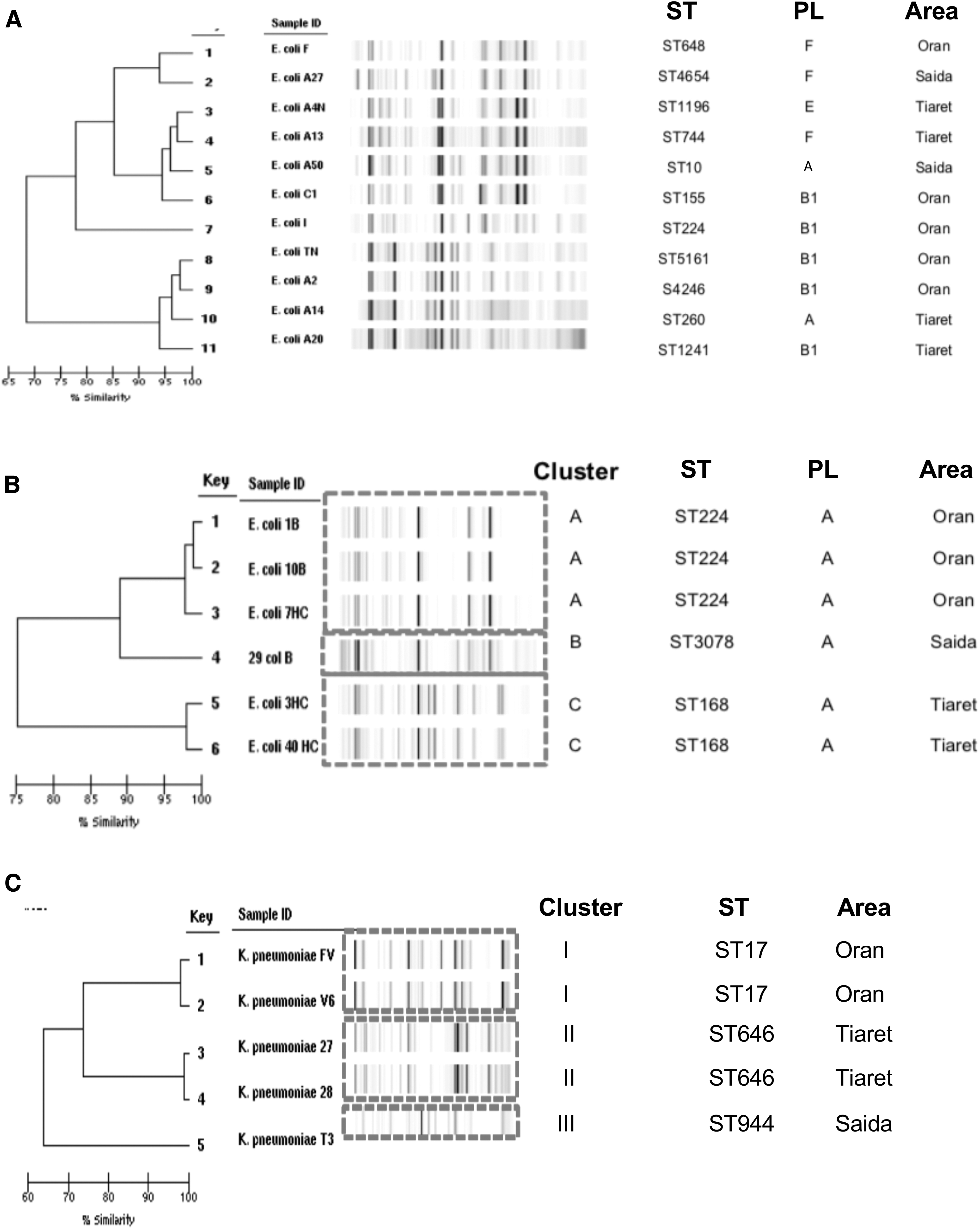
Dendrogram illustrating the genetic relatedness (DiversiLab), ST, and geographical distribution of 22 colistin-resistant Enterobacterales strains isolated from chicken meat in Western Algeria.
The Rep-PCR analysis of the five col-R K. pneumoniae isolates also showed diverse origin with the existence of three clusters belonging to three STs: ST17 (pattern I, n = 2), ST646 (pattern II, n = 2), and ST944 (pattern III, n = 1) (Table 1; Fig. 2).
Plasmid profiles of E. coli
Majority of the mcr-1 encoded col-R E. coli had IncF plasmid (90.9%), but single isolate carried IncHI2 plasmid (9.1%) (Table 1). All these plasmids carrying mcr-1 gene were successfully transferred by conjugation (Supplementary Table S2). All transconjugants exhibited similar resistance patterns, the presence of mcr-1 gene, and only one type of plasmid compared to parenteral strain (Supplementary Table S2).
Phylogroups and virulence profiles of col-R isolates
Phylogroup analysis of E. coli revealed three different categories as follows: phylogroup A (n = 8, 47.1%), phylogroup B1 (n = 5, 29.4%), and phylogroup F (n = 4, 23.5%) (Table 1; Fig. 2). Interestingly, one isolate (A50) belonged to the phylogroup A but harbored some virulent traits (Supplementary Table S3).
Concerning virulence profiles, fimH, iroN, iutA, iucD, traT, cva, and iss genes were present in all col-R E. coli isolates, whereas papG2, papAH, afa/draBC, astA, and fuyA were absent (Supplementary Table S3). Nine of the 11 mcr-1-positive E. coli and five of the six mcr-1-negative E. coli harbored ompT. Six of the 11 mcr-1-positive E. coli had papC, hlyA, ireA, and malX genes. Comparison between mcr-1-positive and mcr-1-negative isolates showed an equivalent repartition of the majority of the genes except for papC and ireA genes, which were significantly associated with six mcr-1-positive isolates (p < 0.01) (Supplementary Table S3). The five col-R K. pneumoniae had low numbers of VFs, possessing 6–9 genes per isolate of the 19 genes screened. All these isolates harbored fimH, ureA, mrkD, entB, uge, and wabG. Neither capsular serotype genes nor hypermucoviscous phenotype was detected (Supplementary Table S3).
Genomic environment of mcr-1 and particularities of the uropathogenic E. coli A50
The genome size of E. coli A50 was 4,973,096 bp with a 50.5 GC%. This isolate possessed four out of five genes (sfa/foc, afa/dr, iutA, kpsMTII, and papA/papC), characteristic of extraintestinal pathogenic E. coli (ExPEC). Moreover, different pathogenicity islands (PAIs) (PAI II536, PAI III536, PAI IV536), typically identified in uropathogenic E. coli (UPEC) strains, 16 and the presence of traD gene were observed in E. coli A50 (Fig. 3). MLST analysis assigned this isolate to ST10.
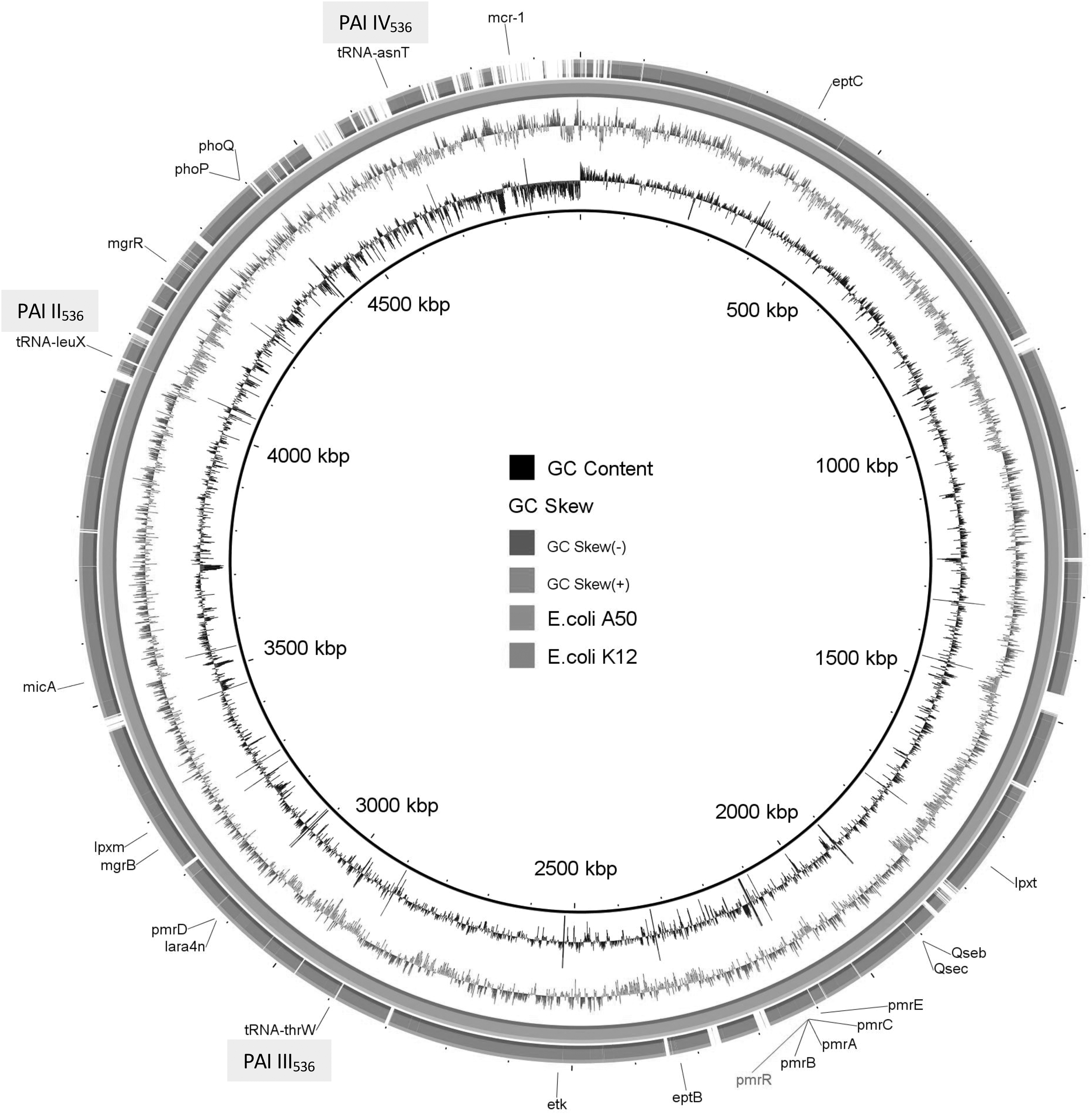
Genetic organization and structure of the uropathogenic E. coli strain (A50) carrying mcr-1 with colistin minimum inhibitory concentration = 96 mg/L. Comparative analysis of E. coli A50 compared to E. coli K12 using the BLAST Ring Image Generator. The concentric rings display similarity between the reference sequence in the inner ring and the other sequences in the outer rings. The varying color levels indicate a BLAST result with matched degree of shared regions, as shown to the right of the ring. Names of some genes are shown by different colors. In gray, the different pathogenicity islands are indicated.
The complete sequence of plasmid carrying mcr-1 gene of E. coli A50 (GenBank accession no. SAMN12501891) revealed that this plasmid was 115,239 bp in size, containing 66 open reading frames with an average of 42.3 GC% content and belonging to the IncFIIK incompatibility group. Using the Neighbor-Joining algorithm, pA50 was compared with other IncF plasmids (e.g., pMR0716_mcr1 and pChi7122–3) that harbored the mcr-1 gene distributed worldwide. This analysis revealed maximum score (2 × 105), coverage (90%), and identity (98.9%) with pChi7122-3 (Fig. 4). As expected, pA50 was successfully transferred to E. coli J53 confirming that the mcr-1 is located on a conjugative plasmid. In addition to the mcr-1 gene, transconjugant isolate possessed the blaTEM −1 gene and carried sulfonamide resistance genes (sul1, sul2, sul3) (Supplementary Table S3), which were in accordance with the resistant phenotype.
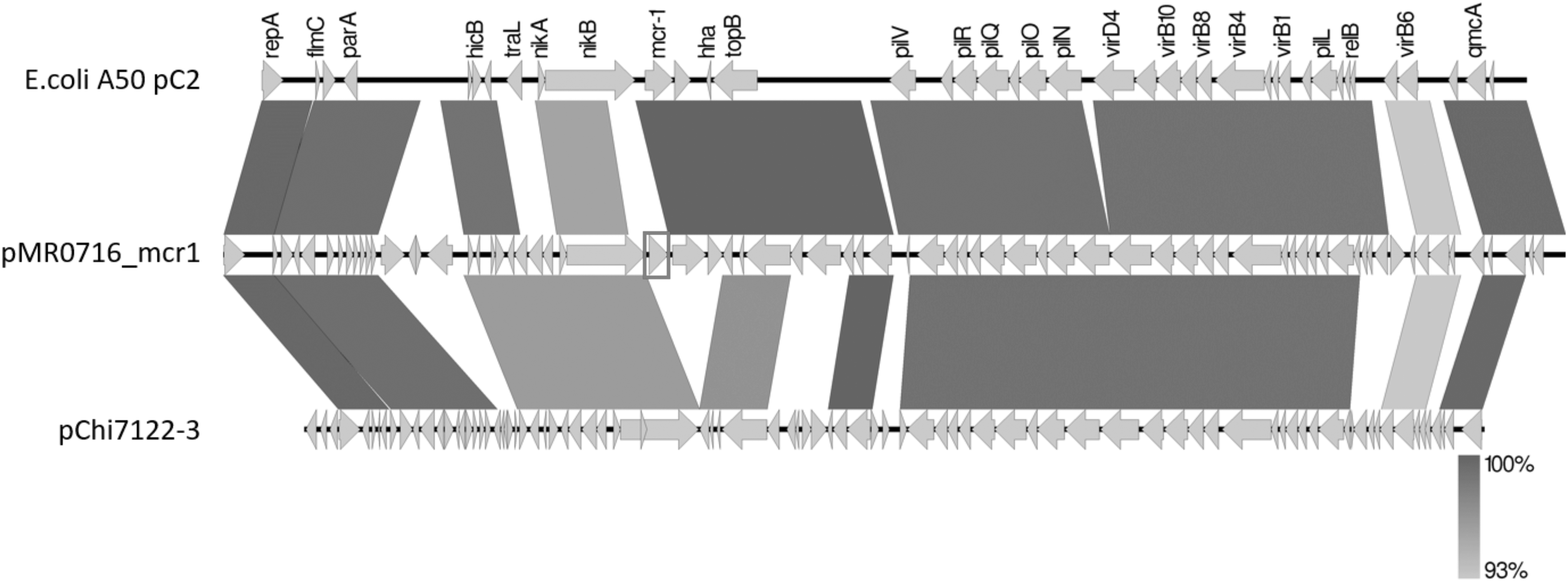
Plasmid structure of the mcr-1-harboring pC2-mcr-1 plasmid characterized in our study in comparison with pMRO716 and pChi7122–3. Gray shading indicates shared regions of homology, whereas white shading indicates a mutation in the region. The intensity of the gray indicates the percentage of homology between plasmidic sequences. Open reading frames are indicated by arrows.
Discussion
After the first description reporting a high frequency of the mcr-1 gene from a collection of Enterobacterales isolated from chicken meat in China, several studies have reported this worldwide problem with prevalences varying between 1.9% and 28% in Asiatic countries (Japan and China),17,18 1.5% and 24.8% in European countries,19,20 and 19.5% in Brazil.21,22 This emergence and diffusion of mobile colistin resistance genes are largely disseminated by horizontal and/or lateral transfer into diverse ecosystems. 23 In Algeria, Enterobacterales carrying mcr-1 have been isolated from humans, environment, wildlife, and poultry feces.7,24–27 This trend was also noted in Tunisia where studies have reported the presence of this gene in chicken meat.28,29 In our study, we showed the presence of col-R Enterobacterales and notably the importance of mcr-1 gene in chicken meat from three public Algerian farms confirming a worrying situation. This observation is not suprising while colistin is of considerable importance in veterinary medicine, especially in the treatment of gastrointestinal infections in production animals, as well as in prophylaxis or metaphylaxis. In the farms visited, a supplementation of the food with 75,000 UI of colistin was systematically performed before feeding chicken. This addition was done each day during the 6 weeks to avoid chicken infection or contamination. We also noted a very high level of resistance to (fluoro)-quinolones (n = 22, 100%) and cotrimoxazole (n = 21, 95.5%) in the col-R isolates, two other antibiotic families commonly used during poultry farming (20 mg of ofloxacin and 0.15 mg of sulfamide were administrated each day to each chicken) (Fig. 1). All together, we objectified the impact of the antibiotics used (colistin, cotrimoxazole, and ciprofloxacin) in these three farms on the selection and the emergence of MDR bacteria. 28 If in developed countries, the use of antibiotics is strictly controlled and documented notably in veterinary use; in Algeria, veterinary antibiotics are often readily sold in shops and markets without prescriptions. 30 This misuse of antimicrobial agents has been related to the incidence of MDR in food-producing animals. 4
Interestingly, the presence of mcr-1 gene was exclusively found in E. coli isolates. Since its description, this resistant mechanism has classically emerged in this species 5 before to diffuse in other Enterobacterales and Moraxella species. 23 E. coli is generally considered a useful indicator of antimicrobial resistance due to its medical importance and its presence in a wide range of hosts. 4 Our study confirms that poultry is a reservoir for col-R E. coli strains in Algeria, as recently suggested. 8 Ten out of 11 mcr-1-positive isolates harbored no chromosomal mutations in any of the different genes screened. This result is in accordance with the lower colistin MICs in mcr-1-positive isolates versus mcr-1-negative isolates. 31 Among the mcr-1-negative E. coli, there are different chromosomal mutations in the two-component systems: phoP/phoQ and pmrA/pmrB, the main targets of col-R E. coli isolates. 32 Likewise, all mcr-negative K. pneumoniae harbored chromosomal mutations in phoP and pmrB. Mutations in both PmrAB and PhoPQ component systems have been previously described among col-R E. coli and K. pneumoniae strains, 32 notably isolated from chicken meat originated from Algerian farms. 1 However, we identified some new mutations never previously described in K. pneumoniae isolates in pmrB (G20S) and phoQ (A70K, D90H/N, P72N, D74E, Q92S, I75L, E77D, and K94E) genes (Table 2). Classically, the most important chromosomal mechanism of col-R in K. pneumoniae is an alteration of the mgrB gene, which encodes a negative regulator of phoP/phoQ system. 33 Further study is needed to fully understand the impact of these mutations on the acquired colistin resistance mechanism, especially since no mutations were detected in mgrB, pmrK, and ccrA genes. Globally, mcr-1 and pmrAB mutations are the main reported colistin resistance mechanisms in our collection as previously observed in an Algerian study. 1
If the use of colistin has induced the emergence of plasmid-mediated mcr-1 gene, we observed no presence of ESBL or PMQR genes in our isolates. The high consumption of quinolones (20 mg each day) in the three studied farms has conducted high level of resistance to this antibiotic family but with probably exclusive chromosomal mutations in gyrase-encoding genes. However, the amoxicillin (20 mg each day to each chicken) and sulfamide (0.15 mg) use on the three farms during the growth period of chicken could be linked to the high resistance to these antibiotics and the selection of isolates carrying penicillinase and sulfonamide resistant genes. This association was very recently observed in E. coli isolates recovered from fish gut in the Mekong Delta, Vietnam. 34 This coexistence of resistant genes was noted in ESBL-producing E. coli, a resistance mechanism absent in our study. These results, associated with the unknown situation of usage of various antimicrobial agents in the veterinary setting, support the role of antibiotic overuse in the emergence of MDR bacteria.
We observed that the Enterobacterales strains isolated in our study belonged to a great variety of STs suggesting that the diffusion of col-R pathogens in the three poultry farms of Western Algeria had diverse origins. Among the different STs reported, E. coli ST648 has been described in Barbary macaque in Algeria and harbored blaCTX-M-1, blaTEM−1, and qnrB19, 24 while E. coli ST744 has been detected in broiler chickens and harbored blaCTX-M-1, tetA/B, qnrB, and sul1/sul2/sul3. 35 Interestingly, E. coli ST155 and ST10 have been diffused in different sources (animals and patients), and E. coli ST1241 has been also detected in seawater in the same Algerian region. 26 Majority of the mcr-1 encoded col-R E. coli had IncF plasmid (90.9%). All these plasmids carrying mcr-1 gene were successfully transferred by conjugation, in accordance with studies on IncF plasmid family originated from diverse host strains and different species isolated from different geographical locations. 36 This group is the most frequently described plasmid type present in MDR bacteria isolated from human and animal. IncF plasmids are frequently associated with the carriage of ESBLs, carbapenemases, aminoglycoside-modifying enzymes, and PMQR-encoding genes, 37 an association not found in our isolates. All transconjugants exhibited similar resistance patterns, the presence of mcr-1 gene, and only one type of plasmid compared to parenteral strain. While no clonal dissemination was detected in our study, a plasmidic dissemination between Enterobacterales is likely. Moreover, we observed that the col-R K. pneumoniae had no hypermucoviscous phenotype compatible with a low virulence lineage. 38 Moreover col-R E. coli belonged mainly to phylogroups A and B1 suggesting that these isolates had a commensal origin. 39 Carriage of antibiotic resistance genes on transposable elements and mobile plasmids in commensal E. coli is common. This renders the resistome highly dynamic, as recently suggested. 37 It is now admitted that some commensal strains can initiate long-term intestinal colonization maintaining plasmid harboring MDR marker. 40 This situation contributes easily to the dissemination of the resistant genes to diverse bacteria among different niches (environment, human, and animals), as observed recently for OXA-48-producing Enterobacterales in this country. 12
Among the isolates, one E. coli, A50, harbored different particularities. Its high colistin MIC is explained by the association of both the mcr-1 gene and novel mutations in different acquired colistin resistance encoding genes. Recently, Zhang et al. suggested that mcr-1 facilitates selection of high-level colistin resistance mutations in E. coli but not in K. pneumoniae, 31 a same trend observed in our collected strains. The mutations detected in the A50 isolate were different than those detected in the others, possibly explaining these elevated MICs. As described above, the genetic analysis showed two novel mutations in the pmrAB two-component system, including a deletion in position 363 in the pmrB gene and a silent substitution (A84G/S29G) in the pmrA gene (Supplementary Table S1). We also detected mutations in the phoP gene (T468C) and in qseC (23 mutations) and qseB (3 mutations: C254A, T508C, T639C [silent]) (Supplementary Table S1). The contribution of these mutations to colistin resistance should be investigated.
Moreover, E. coli A50 belongs to the phylogroup A but harbors some virulent traits and PAIs (Fig. 3), characteristic of ExPEC. The presence of these virulent factors in E. coli A50 is of concern demonstrating the ability of commensal lineage to acquire virulence traits and possess potential to cause an infection. This also suggests that the intestinal microbiota represents an important site of genetic exchange between bacteria.39,41 The commensalism or virulence of E. coli derives from a complex balance between the host status and the expression of VFs. Many E. coli with pathogenic traits potentially inhabit gut of healthy subjects. The way between commensalism and pathogenicity is made fainter by horizontal gene transfer. This transfer favors the diffusion of VFs and antimicrobial resistance markers. The presence of traD gene in E. coli A50 could explain the presence of virulence traits because this gene is a key in the conjugation process, encoding a main component in the transferosome of type IV secretion systems. 42 A study has previously noted that some Avian Pathogenic E. coli strains can colonize the human gut and become a source of UPEC strains. 43
Analysis of the genetic environment of mcr-1 from this isolate showed that the mcr1-pap2 element in pA50-IncF is integrated downstream of the nikB, which keeps both functional domains (calls SMC y NBD94). No repeat/insertion sequence was detected in the neighborhoods of the mcr-1 gene. Furthermore, our study revealed an absence of ISApl1 genetic element upstream of the mcr-1 gene and the presence of pap2 gene in the downstream region of mcr-1 gene. The ISApl1 was described to be involved in the transposition of the mcr-1 gene cassette. 44 Studies have previously noted the absence of this element and concluded that its loss could contribute to the stability of mcr gene on plasmid. 45 The pA50 harboring mcr-1 was easily transferred to another E. coli. The presence of the tra region is also compatible with mobile plasmids. This result is in accordance with other studies, which have reported the great versatility of this plasmid for adaptation to diverse bacteria which has been particularly identified in strains isolated from animal, food, human, and environmental samples and associated with the production of ESBLs, carbapenemases, methylases, and, more recently, MCR-1 enzyme. 46
Our investigation confirms the presence of mcr-1 in E. coli isolated from chicken meat in Western Algeria. Strains harboring the transferable plasmid-mediated colistin resistance had a predominantly commensal origin, which facilitates its dissemination but can also present some virulent traits. The intensive use of antibiotics in the poultry industry could cause the emergence of resistance against many antibiotics in Enterobacterales. These resistances can also be transmitted to human ExPECs as previously noted. 45 More studies are necessary to screen mcr in food-producing animals to understand the spread of this gene in this country where colistin is widely used in veterinary settings. Our study also reflects, once again, the urgent need of a better control in the global market of veterinary drugs.
Footnotes
Acknowledgments
A.P., A.S., and J.-P.L. belong to the Federation Hospitalo Universitaire Infections Chroniques, Aviesan (FHU INCh). The authors thank the Nîmes University hospital for its structural, human, and financial support through the award obtained by our team during the internal call for tenders “Thématiques phares”. The authors thank Sarah Kabani for her editing assistance.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Institute of Health and Medical Research (grant INSERM U1047), the Partenariat Hubert Curien Tassili grant (grant number 17MDU980), and the microbial ecology laboratory at University of Abderrahman Mira (Bejaia, Algeria).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.