
Abstract
Multidrug-resistant (MDR) superficial bacterial infections caused by carbapenem-resistant Enterobacter sp. and Klebsiella sp. have emerged as major threats toward global health care management. In search of a novel antimicrobial, our main objectives were to explore the antimicrobial, antibiofilm, and wound healing potential of glutathione and citrate-capped copper oxide nanoparticles (CuNPs) against gram-negative MDR pathogens Klebsiella quasipneumoniae and Enterobacter sp., ensuring the lowest possible host cell nano-cytotoxicity and minimum susceptibility of the CuNPs toward oxidation. The CuNPs were found to elicit reactive oxygen species (ROS) generation within bacterial cells, inhibiting the bacterial growth and division. They contributed to the remodeling of the bacterial lipopolysaccharide, induced membrane lysis, and promoted antibiofilm activities by reduced cell–cell aggregation and matrix destabilization while displaying excellent biocompatibility against HEK-293 and HeLa cell lines. The CuNPs were also instrumental in preventing postsurgical wound infections and aiding in wound closure in the murine excisional wound model, as observed in albino Wistar rats, forcing us to believe that the CuNPs are bioactive in wound therapy. The results are encouraging and demands further experimental exploitation of the particles in treating other MDR gram-negative infections, irrespective of their resistance status.
Introduction
The ever-evolving pressure of natural selection on the drifting of antigen structure and function has driven the effectivity of antimicrobials, once known as magic bullets, to become a victim of a silent tsunami. This antimicrobial attenuation has triggered the nefarious genesis of several drug resistance groups like multidrug resistant (MDR), extensively drug resistant, and pandrug resistant. Alarmingly, these groups include several microbial genera, previously known to be opportunistic or nonpathogenic. The selection of resistant bacterial strains has resulted in compromised and reduced antibiotic efficacy for advanced antimicrobials like the third-generation cephalosporins, polymyxins, and carbapenems.1–4 In 2017, the World Health Organization released a list of antibiotic-resistant pathogens against which there was an urgent need for novel antimicrobials. 5 These organisms were divided meticulously into medium, high, and critical groups. Carbapenem and third-generation cephalosporin-resistant Enterobacteriaceae were included in the group against which there was a “critical” need for new antimicrobials. 6 This is precisely why the latest technological interventions have focused on the detection of these carbapenem-resistant pathogens from clinical samples 7 for early detection of infection, as the control of these organisms is a complex affair once infection progresses. Superficial cellular components, like polyanionic lipopolysaccharides (LPSs), are structurally diverse and impart remarkable MDR capabilities to these organisms through effective repulsion of hydrophobic and charged antimicrobials.8,9 The inherent negative charge of the LPS poses a threat to these organisms as drugs can be designed to electrostatically target the LPS polar head groups, 9 but unfortunately, this strategy has been greatly underutilized. To add to the misery, these pathogens form extracellular polymeric matrices known as biofilms, which impart certain “emergent properties” to its residents, including elevated resistance against antimicrobials, resulting in an increase in health care costs and exacerbation in the rise of antibiotic resistance. 10 The biofilm residents are also able to perform complex cell–cell interactions governed by quorum sensing between members of the same and different resident species contributing to their survival and adaptive capabilities like multidrug resistance. 11 Almost 80% of biofilm-forming pathogens are associated with persistent infections with a dramatic increase in resistance and tolerance toward most therapeutic antimicrobials compared with their planktonic counterparts.12,13 Hence the suppression of biofilm production or corroding the already developed biofilm could be viable strategies for the prevention of the emergence of these resistant bacteria. Alarmingly, MDR gram-negative organisms, causing postsurgical or opportunistic wound infections, significantly impede the process of healing because of the formation of these biofilms on the wound surface resulting in an urgent requirement of effective antibiofilm and wound healing agents. 14
In recent years, nanomaterials have proved invaluable to medical science as an alternative to conventional treatment regimes because of the rise in chemotherapeutic resistance in both infectious and noninfectious diseases. Owing to the high reactivity and unique physiochemical properties of nanoparticles compared with their bulk form, 15 they have profound modulatory effects on biological systems. For some metallic and nonmetallic/polymeric nanoparticles, the antibacterial activity can extend to the verge of biocidal activity,16,17 which can be attributed to their intricate interactions with the bacterial cell membrane causing lysis or affecting bacterial metabolic pathways.18,19 Different silver and copper formulations have been shown to kill bacteria by elicitation of reactive oxygen species (ROS) and binding to the thiol groups of bacterial proteins.17,20 Copper, as a metal, is known to possess impressive antimicrobial activity, 17 but on nanoparticle formation, gets rusted easily, 21 preventing their extensive use in medical, food, and mechanical sciences. In addition, being a metal nanomaterial, there is always the threat of CuNPs being toxic on exposure directly or through the environment. 22 Fortunately, a lot of research is being dedicated to synthesizing biocompatible CuNPs and composites with potential for environmental and the clinical application as antibacterials.
In this specific study, our prime objective was to explore the in vitro antibacterial efficacy along with the in vivo wound healing potential of the glutathione—citrate-capped copper oxide nanoparticles (CuNPs) against two gram-negative cephalosporin and carbapenem-resistant MDR pathogens namely Klebsiella quasipneumoniae (Klebsiella) and Enterobacter sp. (Enterobacter), both implicated in notorious wound infections.23,24 The central idea was to aim at precise bacterial molecular targets like the gram-negative LPS for CuNP attachment and dissolution, ROS elicitation for the consequential oxidative damage to cell structures and membranes, and the exopolysaccharide-based biofilm matrices for corrosion of the existing biofilms and prevention of biofilm neoformation to completely inundate the antimicrobial-resistive capabilities of the MDR organisms. Most importantly, the biocompatibility of the CuNPs was ascertained by studying the probable toxicological effects of the particles against HeLa and HEK-293 cell lines. The study concluded with the investigations concerning the CuNP-mediated wound healing and antisepsis using albino Wistar rats.
Materials and Methods
Antibiotic resistance profiles were obtained using the well-known antibiotic disk diffusion assay for multiple antibiotics. The physicochemical characters of the CuNPs were studied using high-resolution transmission electron microscopy (HR-TEM), zeta potential, X-ray diffraction (XRD), and Fourier transform infrared spectrometry (FT-IR). Minimum inhibitory concentration (MIC), minimum bactericidal concentration (MBC), growth curves, and time–kill curves demonstrated the inhibitory effects of the CuNPs on Enterobacter and Klebsiella. ROS generation was confirmed by fluorescence microscopy and flow cytometry using 2, 7-dichlorofluorescein diacetate (H2-DCFDA). Sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) and silver staining of bacterial LPS showed molecular alterations in the O antigen modal length on CuNP administration. The cellular damage to the test MDR organisms was determined using membrane disruption assays, live/dead assay, and atomic force microscopy (AFM). Biofilm disruption and clearance were ascertained through the crystal violet (CV) and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assays along with scanning electron microscopy (SEM). Finally, we prepared a CuNPs containing nano-ointment (NO) to study the wound healing effects of the CuNPs on albino Wistar rats after ensuring the biocompatibility of the particles by testing them on HeLa and HEK-293 cell lines. All animal studies were conducted after obtaining ethical approval from the Institutional Animal Ethics Committee (IAEC) of the University of Kalyani as per guidelines put forward by the Committee for the Purpose of Control & Supervision of Experiments on Animals (CPCSEA), New Delhi, India. The detailed experimental procedures can be found in the Materials and Methods section in supporting information.
Results
Antibiotic resistance profile
Both the test organisms Enterobacter and Klebsiella were resistant to several antimicrobials including cefotaxime and imipenem (Supplementary Fig. S1 and Supplementary Tables S1 and S2). Extended-spectrum beta-lactamase double-disk diffusion test, and polymerase chain reaction showed that both the test organisms were positive for BlaCTX-M (Supplementary Fig. S2) but were negative for production of BlaSHV and BlaTEM. The imipenem resistance hinted at metallo beta-lactamase/carbapenemase production. 25
Physicochemical characteristics of CuNPs
The method of synthesis, HR-TEM micrograph, and exact dimensions of the CuNPs have been kept confidential owing to future patenting possibilities but can be readily obtained on request from the corresponding author. The particles were spherical with an average size of <10 nm. The CuNPs possessed a low zeta potential of −9.85 mV (Fig. 1a) despite being exceptionally stable at room temperature, further strengthening the belief that stabilization of all nanoparticle dispersions cannot be explained by classical electrostatic stabilization models. The obtained XRD peaks of the polycrystalline CuNPs were matched with JCPDS card no. 05-0667 and 04-0836 to analyze their composition. It was determined that they were predominantly composed of crystalline cuprous oxide (Cu2O), which were found to exist in 110, 111, 200, 220, and 311 crystalline planar phases and metallic copper crystallites existing in the 111, 200, and 311 planar phases (Fig. 1b). The Cu2O and Cu crystallites were most likely formed because of the complete and incomplete reduction of the Cu2+ ions by citrate and hydrazine. 17

Physicochemical characterization of CuNPs:
The bands appearing in the CuNP FT-IR spectra had the following probable assignments. The band near 1384.5 cm−1 was likely owing to –OH bending vibrations of capped glutathione and citrate, whereas the broad band near 3320–3280 cm−1 signified stretching of hydrogen-bonded –OH groups (Fig. 1c). The two bands at 2852.7 and 2922 cm−1 appeared owing to asymmetric and symmetric C-H stretching modes of hydrocarbon chains in both glutathione and citrate. The amide I band of the protein carbonyl stretch of glutathione and –COO of citrate was noted at 1585.8 cm−1. The strong –NH stretching band of the amino acids were assigned at 1460.4 cm−1. The intense band at 620.4 cm−1 was designated to Cu-O interactions 26 of Cu2O and is per our XRD data.
Growth and susceptibility analysis
Preliminary inhibition studies performed over a 4-h time period demonstrated clearly that the growth of both organisms was inhibited by 200 and 400 mg/L concentrations of CuNPs (Supplementary Fig. S3). MIC for both organisms against the CuNPs was found to be 350 mg/L, as complete cessation of observable growth occurred at this concentration. The MBC value was found to be 400 mg/L. For Enterobacter, the IC50 of CuNPs was calculated to be 188.6 mg/L and for Klebsiella, the IC50 of CuNPs was 179.9 mg/L (Supplementary Fig. S4). On performing the growth curve analysis of both the organisms versus increasing doses of CuNPs, significant retardation in growth was observed for all four test CuNP concentrations (Fig. 2a) compared with untreated controls. The time-kill curves demonstrated that compared with untreated bacteria, there was >3-log10 reduction in bacterial growth for both organisms against MIC of CuNPs, whereas for IC50, there was a 1.59-log10 reduction in Enterobacter and 0.67-log10 reduction in Klebsiella viable cell counts, respectively (Fig. 2b).

Bacterial growth analysis and production of ROS by CuNP treatment:
Generation of ROS
The production of ROS owing to CuNPs treatment was measured by the oxidation-sensitive fluorescent probe H2-DCFDA. Figure 2c shows that there was a significant increase in dichlorofluorescein (DCF) fluorescence at both 0-h and 4-h (IC50)-treated Enterobacter and Klebsiella cells compared with untreated controls, but not overwhelmingly high, as seen in the positive control. Quantification of the mean fluorescence intensity (MFI) based on fluorescence microscopy showed that there was a > 5 × increase in DCF fluorescence intensity in the 4-h CuNP-treated Enterobacter cells compared with the 0-h treated cells. Similarly, a > 3 × increase in fluorescence intensity was observed for 4-h CuNP-treated Klebsiella cells compared with 0-h samples (See Supporting Information/Supplementary Data). Flow cytometry analysis showed an increase in percent cellular visualization in Enterobacter cells from 0.9 × to 1.79 × for the 0-h and 4-h treated sets, whereas a 1.79 × to 2.86 × increase was noted for Klebsiella cells (Fig. 2c, d) confirming the production of ROS in these organisms, affecting the growth of Enterobacter and Klebsiella cells.
Effect of CuNPs on bacterial cells: change in integrity and morphology
In this study, we observed anomalous elongation of CuNP-treated Enterobacter and Klebsiella cells in a time-dependent manner (Fig. 3a). A significant increase of A260 in the cell-free extracts of control and treated Enterobacter and Klebsiella was observed for both bacterial candidates after the administration of IC50 CuNPs. In Enterobacter, the increase in A260 was observed from the seventh hour post-treatment (Fig. 3b), whereas, for Klebsiella, it was from the fifth hour compared with respective controls. Confocal laser scanning microscopy-assisted live/dead assay using 4′, 6-diamidino-2-phenylindole (DAPI) and propidium iodide (PI) revealed a very low DAPI uptake by the control cells (Fig. 3c). But on CuNP treatment, the number of DAPI-stained cells along with stained PI cells went up considerably. The simple interpretation of this phenomenon is that owing to varying degrees of nonspecific, random membrane damage, some cells became semipermeable, making way for DAPI to enter but not PI, thus the blue fluorescence. At the same time, many cells were heavily damaged with highly compromised membranes and were stained with PI or DAPI+PI resulting in red or magenta fluorescence, respectively.

Loss of cellular integrity of CuNP-treated Enterobacter and Klebsiella cells:
SDS-PAGE followed by silver staining of LPS obtained from control and CuNP-treated Enterobacter and Klebsiella cells showed that the treated bacteria produced more LPS with significant shortening in the size of Oag in the treated cells compared with the untreated controls (Supplementary Fig. S5). AFM of the bacterial ultrastructure of the control Enterobacter and Klebsiella cells clearly showed that the cellular shapes are legible and the cells appear undamaged (Fig. 4). The extent of damage to the cells on treatment with the CuNPs appears significant. The treated Enterobacter cell looks deformed with signs of membrane leakage with a large surface bleb along the z-axis. Contrarily, the Klebsiella cells seem to have elongated multifolds resulting in clumping of the cells along with cellular disintegration.
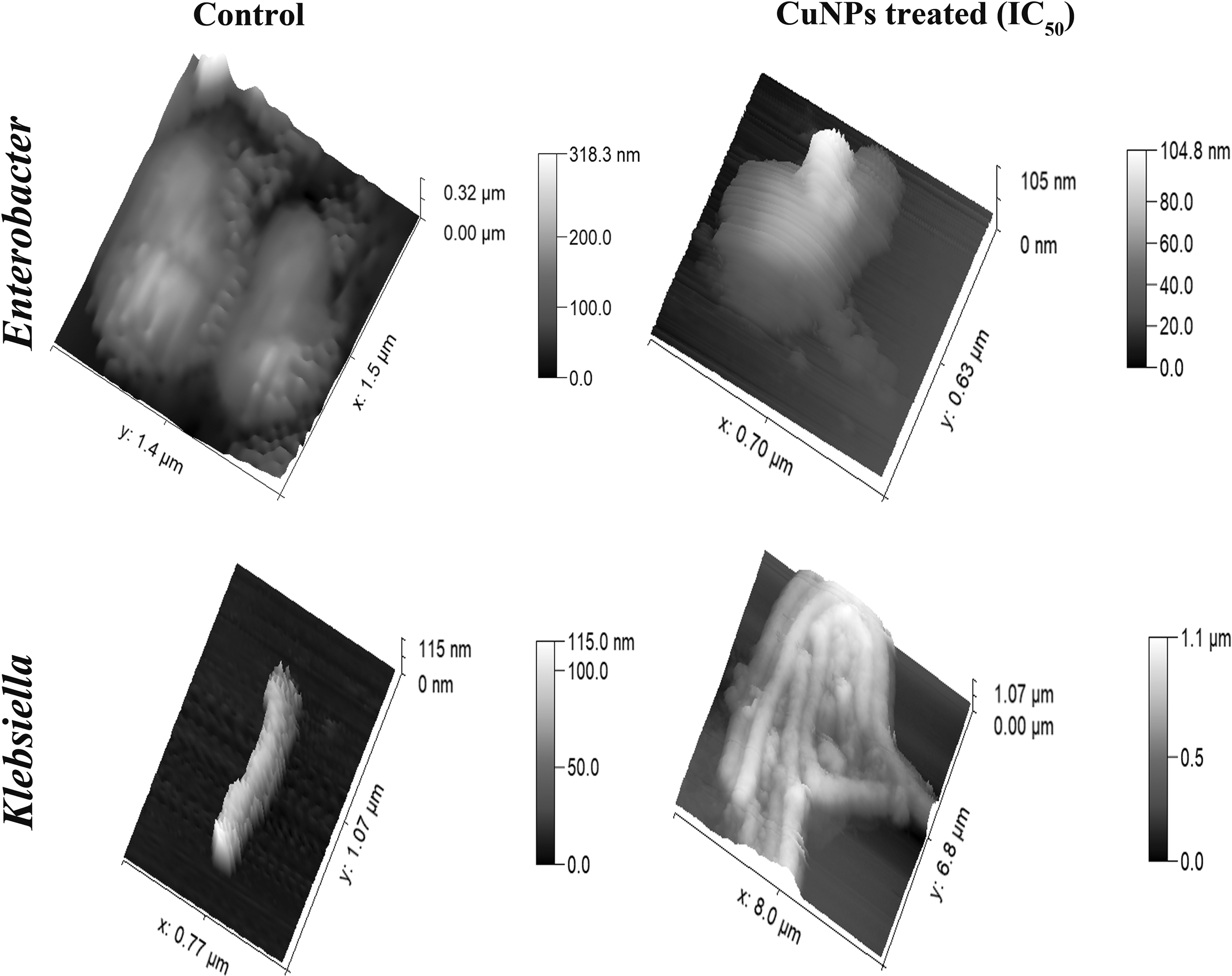
AFM micrographs showing the bacterial surface ultrastructure post-CuNP treatment: Untreated Enterobacter cells showing smooth surface topology and normal cell size as captured by AFM compared with a CuNP (IC50)-treated cell showing membrane leakage and surface blebbing with an overall irregular cellular morphology (top panel); Untreated Klebsiella cell showing normal cellular shape and size compared with CuNP (IC50)-treated cells that seem to have elongated many folds to form cellular clumps with visible membrane damage (bottom panel). AFM, atomic force microscopy.
Antibiofilm activity
Both Enterobacter and Klebsiella were vigorous extracellular polymeric substance (EPS) producers (Supplementary Fig. S6) and formed intact, uniform biofilms in unchallenged conditions. Results of the CV assay showed that there was a dose-dependent inhibition and eradication of biofilms for both the organisms (Fig. 5b, c). When the formation of biofilms in the presence of increasing concentrations of CuNPs was studied, 26.46% of the Enterobacter biofilm matrix was eradicated by the lowest CuNP concentration (100 mg/L) (Fig. 5b). As the dose was increased up to MIC, a 3.2-fold increase in eradication was observed. Similar eradication trends were observed for 400 and 500 mg/L. For Klebsiella, 25.76% eradication was achieved for 100 mg/L and for the MIC, an impressive 3.6-fold matrix clearance was found. For 400 and 500 mg/L, the clearance was 86.03% and 86.1%, respectively. Against preformed Enterobacter biofilms, the eradication was 20.92% for 100 mg/L, 65.13% for MIC, 74.74% for 400 mg/L, and with a maximum of 90.99% for 500 mg/L. The effect of the CuNPs on Klebsiella showed a similar eradicative pattern (Fig. 5c).
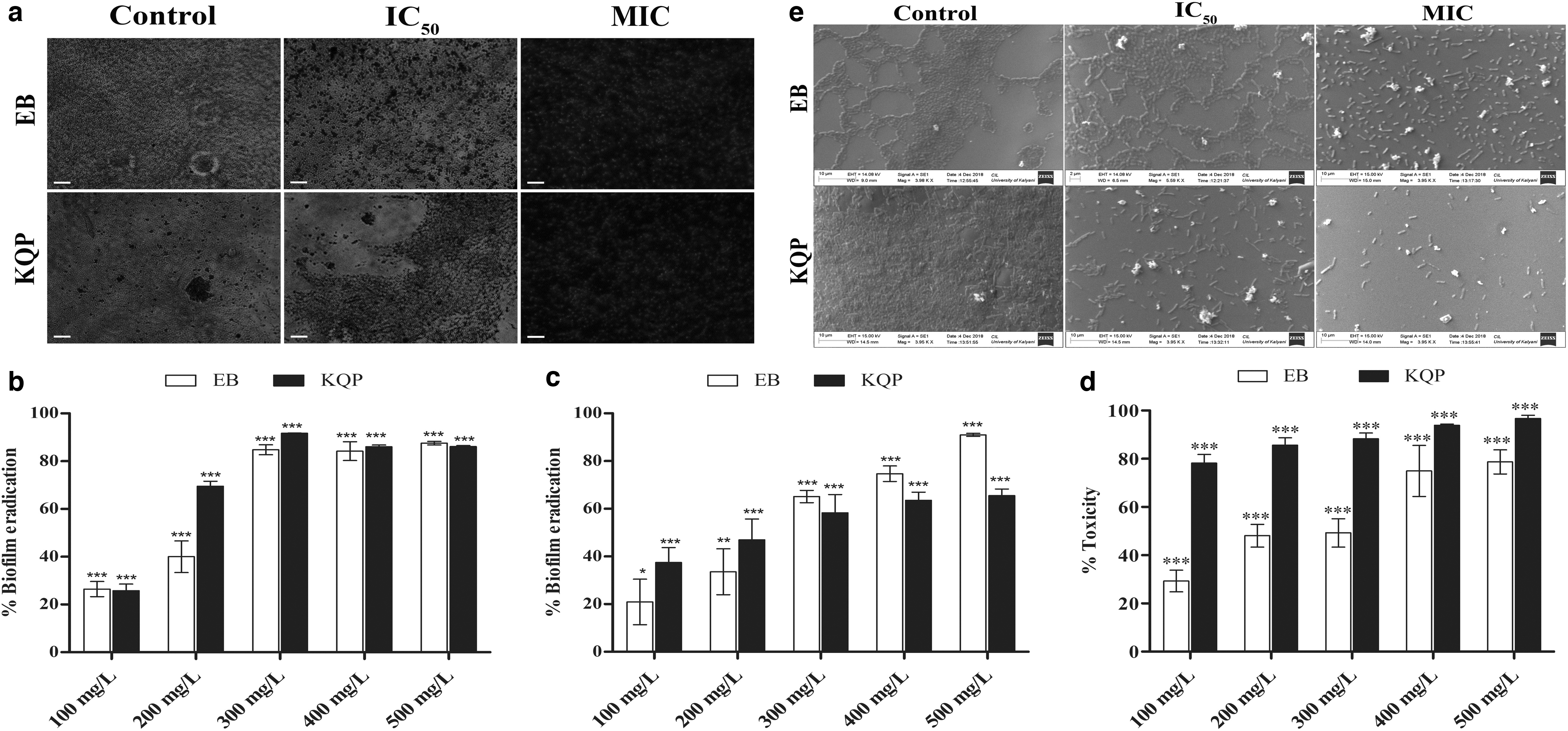
Antibiofilm activity of the CuNPs:
An MTT reduction assay was performed to check the viability of bacteria in the CuNP-treated biofilms. With increasing CuNP concentrations, the death of the metabolically active Enterobacter cells in the biofilms steadily rose from 21.27% to 70.71% (Fig. 5d). In contrast, the CuNPs proved to be extremely toxic toward the biofilm residing Klebsiella cells as only 21.82% cells survived the lowest CuNP concentration and the viability came down to 3.31% for 500 mg/L.
The results of fluorescence microscopy and SEM showed that the bacteria formed beautiful, well-packed, and uniform biofilm matrices in normal, physiological conditions (Fig. 5a, e). On administration of IC50 CuNPs, the biofilm integrity and uniformity were severely disturbed as prominent regions of disrupted matrices surfaced. At MIC, the biofilms appeared visibly dismantled with a drastic decrease in biofilm inviolability, evident by complete loss of cell–cell adherence and extensive cellular damage.
Cytotoxicity test on human cell lines
The evaluation of toxicity studies of any synthesized CuNPs on human cell lines is of paramount significance before considering any clinical application of the particles. The MTT assay on HEK-293 cells showed the there was 92.92% survivability of cells at 200 mg/L, whereas 86.54% of cells were viable at 100 mg/L concentration. Unexpectedly, there was an increase in the cellular viability at the highest CuNP concentration, which was ∼98.34% as seen in Fig. 6a. In the case of HeLa cells, the viability was found to be 92.12% at 200 mg/L followed by a decrease in viability to ∼84.34% (100 mg/L) and finally culminating at 97.31% for 3.125 mg/L. Results of the trypan blue assay showed a steady increase in the percent cellular viability from 77.98% to 97.95% for HEK-293 and from 79.36% to 99.45% in HeLa cell lines with decreasing CuNPs dosage (Fig. 6b). It has been reported previously that elicitation of ROS by nanoparticles can cause overestimation of cellular viability because of the superoxide-induced reduction of tetrazolium salts like MTT and (2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide) (XTT) to MTT-formazan and XTT-formazan, respectively. 27 As our CuNPs are capable of eliciting ROS within cells, (see Supplementary Fig. S7) there is a strong probability that this phenomenon facilitated the reduction of tetrazolium into formazan crystals, contributing to the overestimation of cellular viability at high concentrations for both HEK-293 and HeLa cell lines that were further corrected and implicated by the trypan blue assay.

Cytotoxicity of CuNPs toward HEK-293 and HeLa cell lines:
In vivo wound healing in albino Wistar rats
The role of CuNPs in wound healing was investigated by comparing the rates of dermal wound closure in CuNPs containing NO-treated and untreated control groups using the excisional wound healing assay. In NO-treated rats, the excisional wound diameter was reduced by >80% by the seventh day compared with a little over 20% for the untreated controls (Fig. 7a). The NO-treated wounds closed completely by the 13th day of excision, whereas ∼40% of the wound diameter remained open for the untreated controls (Fig. 7b). Furthermore, on-site infection of Enterobacter and Klebsiella considerably delayed the wound healing process in the untreated controls compared with the absence of any infection in the NO-treated wounds. On gross examination of the wounds during the healing process, it was observed that the quality of wound healing was significantly higher and scarless in the NO-treated wounds compared with the untreated controls.
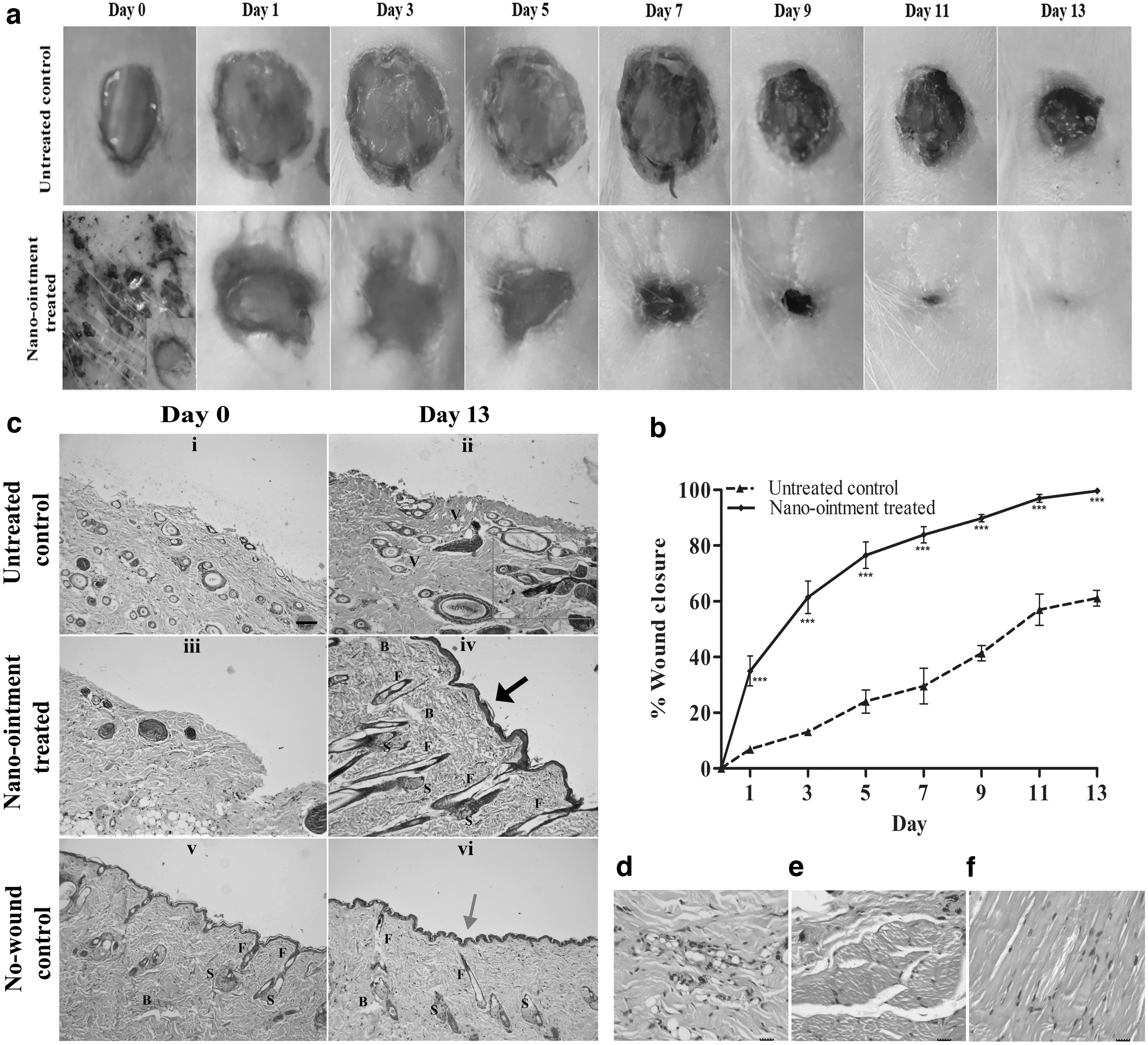
In vivo wound healing:
The hematoxylin and eosin-stained sections of healing wounds of NO-treated, untreated controls, and nonwounded controls pointed out a few key differences in the wound healing process of the NO-treated and untreated control animals. On the 0th day, there was a complete absence of epidermis and papillary dermis in both control and treated sets with scanty reticular dermis (Fig. 7c-i, c-iii). By the 13th day, neoformative epidermis layer and epidermal ridges were observed in the NO-treated rats compared with little or no formation of the epidermis in the untreated controls (Fig. 7c-ii, c-iv). This neoformative and keratinized squamous epithelium of the NO-treated rats was found to be thicker compared with the nonwounded controls, suggesting the formation of more cell layers than normal (Fig. 7c-iv, vi). In addition, the formation of significantly greater numbers of primitive hair follicle structures and sebaceous glands can be observed in the NO-treated animals in comparison with the untreated controls and nonwounded controls. There were clear signs of early keloid scarring in the untreated controls evident from the formation of glassy collagen (Fig. 7c-ii). The NO-treated sections showed normal angiogenesis, whereas numerous congested and ill-differentiated blood vessels were observed in the untreated controls. The collagen arrangement was also more regular and ordered in the NO-treated samples. The stained sections of the untreated controls showed more neutrophil and macrophage infiltration indicating a high inflammatory reaction to the wound infection (Fig. 7d). Contrarily, lesser numbers of inflammatory cells in the hypodermal and dermal tissues of the NO-treated sections revealed that the inflammatory reaction was comparatively milder, indicating infection clearance and anti-inflammation facilitated by copper, restoring the nonwounded state (Fig. 7e, f and Supplementary Fig. S8).
Collectively, the results of the wound healing assay, as well as the histopathological investigations, suggests that the CuNPs containing NO actively promotes reepithelialization, typical angiogenesis, hair follicle, and sebaceous gland regeneration, as well as the ordered arrangement of the collagen fibers compared with the untreated control's poor wound healing capability. This accelerates orderly wound closure in rats, along with on-site clearance of infections caused by potentially hostile MDR pathogens.
Discussion
Both organisms that we have studied, that is, Enterobacter sp. and K. quasipneumoniae displayed resistance against a carbapenem (imipenem) along with third- (cefoperazone, cefotaxime) and fourth-generation (cefepime) cephalosporins, and yet, the nanotechnological control of these organisms or their close carbapenem-resistant relatives has not been reported for wound infections. In literature, the most effective nanomaterials that have been tested against these “critical” pathogens are silver nanomaterials (AgNMs), which have shown effective biocidal activity against related carbapenem-resistant organisms like Acinetobacter baumannii. 28 Although copper nanomaterials have shown impressive antimicrobial activity against MDR organisms, they have been rarely put to test against carbapenem-resistant Enterobacteriaceae. The probable reason is the availability of rapidly synthesizable and stable AgNMs, which are abundant in the literature. The tendency of rapid self-oxidation and agglomeration of CuNPs has made this research area even more arduous. 29 Our synthesized CuNPs followed a synthetic route involving stabilization by glutathione, previously used for stabilizing gold nanoparticles, 30 imparting impressive nanoparticulate stability, making it the first antibacterial copper nanomaterial to be glutathione stabilized. The growth and time-kill curves conclusively prove that our CuNP formulation is a potent biocide that does not require any synergistic application with another nanoparticle or antibiotic. 31 The relatively high MIC and biocidal concentration of the particles compared with other CuNPs32,33 coupled with their specificity for gram-negative cells may indicate that they are moderately reactive, which in turn may be an explanation of their robust biocompatibility against HeLa and HEK-293 cell lines.
Based on the design of the study, the results obtained, and the existing literature, a working mechanism of action of the CuNPs can be proposed. The probable initiation of the antimicrobial action of the CuNPs against the test pathogens is by the electrostatic binding of the particles to the bacterial outer membrane (OM) using the polar head groups of OM phospholipids and LPS, 34 causing the shortening of the Oag modal length of the bacterial LPS in the process. Longer Oags have a distinct preventive role in complement activation and evasion in gram-negative bacteria. 35 Protruding tips of long-chain O antigens of LPS molecules present in the OM causes significant hindrance in the formation of the membrane attack complex (MAC) on the bacterial cell surface. Longer Oag modal lengths have been shown to prevent C1 components of the host complement system from accessing bacterial outer membrane for the direct activation of the classical pathway. 36 Thus, shortening in the size of the Oags ensures the deposition of the complement proteins and MAC formation on the bacterial cells and facilitates antibody-mediated cellular lysis. 37 This is possibly the first report of Oag modal length alteration of LPS by NPs, which can have a profound negative influence on the virulence and pathogenesis of gram-negative pathogens and warrants further investigation. The fact that our CuNPs had negligible effects on gram-positive organisms (data not shown) shows that the particles must have interacted with the bacterial cells physically through the gram-negative LPS and the antibacterial action of the particles were not purely owing to the release of copper ions onto the growth media. 38 This is a significant observation as it narrows down the spectrum of activity of our CuNPs for more specific applications against MDR gram-negative organisms, reducing the toxicological impact on the microbiome along with the delaying of the emergence of resistance. The CuNPs on adsorbing to the gram-negative OM, most likely caused its depolarization along with the reduction in its typical negative charge, making it more permeable. 17 The entry of the CuNPs into the oxidizing gram-negative periplasmic space39,40 probably resulted in the formation of stable copper cations (Cu2+) by the oxidation of Cu2O and metallic copper, causing copper ion leaching from the nanoparticles. These released Cu2+ ions from the nanoparticles then participated in a set of reactions known as “Fenton-like reactions,” a process that promotes the generation of hydroxyl radicals from hydrogen peroxide, 41 similar to the Fenton reactions for Fe2+. On successful initiation of this process, the bacteria were probably killed by ROS-induced lipid peroxidation, DNA damage, and protein degradation. 42 This ultimately resulted in impaired cell division because of the hampered activity of essential cell division proteins like ftsZ, 43 leading towards abnormal elongation of the treated bacteria as has been reported previously from our laboratory. 44 The CuNPs also caused lysis of the bacterial cells, a characteristic biocidal effect of metal NPs.45,46 It should be noted that gram-negative bacteria have extremely well-evolved systems to prevent copper ions from participating in the aforementioned deleterious cellular reactions 47 and once attached to the bacterial cells, the release of copper ions from our CuNPs must have been fast and sustained and presumably bypassed the bacterial defense comprising copper ion efflux by resistance–nodulation–cell division proteins, the periplasmic Cu/Zn superoxide dismutases, and the gram-negative envelope stress response for effective inundation of their defense.
Evidence suggests that the effectivity of engineered nanomaterials against complex biofilm systems depends on several factors like conditions of exposure, stability, concentration, and size.48,49 This implies that the small size of our CuNPs contributed substantially toward its excellent biofilm penetrative and clearing ability. 50 From the perspective of biofilm clearance, citrate acted as a strong chelating agent resulting in the destabilization of the biofilm matrix by chelation of biofilm stabilizing cations like Mg2+. 51 Although recent publications claim that surface modification of CuNPs may result in decreased antimicrobial activity, 28 we believe that the “organic corona” surrounding the NPs comprising citrate and glutathione is vital for the attachment of the CuNPs to the biofilm matrix, which facilitates the penetration of the EPS network by diffusion50,52 and should be investigated further. The penetrated and destabilized matrix, in all probability, facilitated the killing of the biofilm-protected bacteria in the same way as the planktonic ones.
In conclusion, copper ions are known to possess excellent wound healing abilities and have been used in wound dressing materials and antibiofilm nanofiber scaffolds.53,54 Copper has been shown to possess interleukin-10-inducing capabilities along with the facilitation of release of growth factors like vascular endothelial growth factor (VEGF) from healing cells, aiding in anti-inflammation, and wound contraction in rats. 55 Photothermal wound healing therapy involving nanocopper has been recently reported, but understandably requires the involvement of sophisticated instruments and infrastructure. 56 In contrast, our NO is not only cheap and easy to formulate compared with other nanocopper-containing hydrogels, 57 but at the same time accelerates the healing of surgical wounds in rats while preventing on-site infections and biofilm formation, which is a genuine concern in postoperative scenarios. In this worldwide medical crisis, especially in a low and middle-income country like India, our CuNPs can be an effective and inexpensive option in treating complicated topical MDR infections. Its biocidal, antibiofilm, and wound healing capabilities coupled with its cheap manufacturing can result in an effective medicinal formulation, greatly benefiting the rural and underprivileged population of this country.
Footnotes
Acknowledgment
The authors express their utmost gratitude toward Arghya Bandyopadhyay, for his assistance in the synthesis of the CuNPs, and Priya Yadav for her efforts in facilitating the nanotoxicological evaluations of the particles. The authors also acknowledge; CSIR-Central Glass and Ceramic Research Institute, Kolkata; Central Instrumentation Laboratory, University of Kalyani; Department of Botany, University of Kalyani; Department of Chemistry, University of Kalyani and Department of Molecular Biology and Biotechnology, University of Kalyani for providing access to their various instrumental facilities. In addition, the authors thank Anoop Kumar Mukhopadhyay, Dean of Science and Professor of Physics at Manipal University, Jaipur for his invaluable suggestions during the course of this specific study.
Disclosure Statement
No competing financial interests exist.
Funding Information
University of Kalyani provided University Research Scholarship to S.S. DST-PURSE program (University of Kalyani) and DST Nanomission (Project No. SR/NM/NS–1447/2014 G) provided financial support to carry out this research work.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.