
Abstract
Acinetobacter baumannii and Pseudomonas aeruginosa are among the most prevalent pathogens causing a wide range of serious infections in hospitalized patients and contaminating intensive care units and inanimate surfaces. The purpose of this study was to investigate the mechanism of carbapenem resistance in clinical and hospital environmental isolates of A. baumannii and P. aeruginosa recovered from a Libyan hospital. From a total of 82 Gram-negative bacteria, 8 isolates of A. baumannii and 3 isolates of P. aeruginosa exhibited resistance to imipenem with minimum inhibitory concentrations ranging from 16 to >32 μg/mL. Five isolates of A. baumannii harbored blaOXA-23 gene, from which three isolates were collected from patients and two from hospital environment. Only one isolate harbored blaNDM-1 gene, which was responsible for carbapenem resistance in A. baumannii. The OprD gene seems to be disturbed by an insertion sequence (IS) in two isolates and affected by polymorphism in one isolate. Pulsed-field gel electrophoresis results showed high genetic diversity among carbapenemase producing A. baumannii. This study highlights the dissemination of blaOXA-23 and blaNDM-1 genes in a Libyan setting. Therefore, infection prevention and control practices, antimicrobial stewardship initiatives, and antimicrobial resistance surveillance systems should be implemented to prevent the wide spread of antimicrobial resistance.
Introduction
The emergence and spread of multidrug-resistant Gram-negative bacteria constitutes a major public health problem worldwide. 1 Indeed, several studies have already demonstrated the association between infections owing to multidrug-resistant Gram-negative bacteria with a high mortality rate and long hospital stays. 2 Inanimate surfaces have often been described as the source for outbreaks by bacteria causing health care-associated infections (HAIs). Acinetobacter baumannii and Pseudomonas aeruginosa are among the most common pathogens causing serious infections in hospitalized patients and responsible for contaminating environmental setting in hospitals, particularly in intensive care units (ICUs). 3
A. baumannii isolates are increasingly reported causing severe infections among immune-compromised patients, mainly ventilator-associated pneumonia, bloodstream infections, bacteremia, urinary tract infections, wound infections, and meningitis. 4 Acinetobacter infections have frequently been reported as a major cause of HAIs. 5 This pathogen has furthermore the ability to survive for long periods and could easily spread in hospital environments and dry surfaces. 6 It can express high levels of multidrug resistance to antimicrobials, including aminoglycosides, fluoroquinolones, polymyxins, and trimethoprim/sulfamethoxazole, making infections often difficult to treat. 7 Carbapenems are the most effective drugs against infections caused by Gram-negative bacteria including Acinetobacter species. 8 However, carbapenem resistance is currently considered a significant health care problem because of the limited availability of treatment options. 9 In A. baumannii, carbapenem resistance is mostly owing to the production of carbapenem-hydrolyzing β-lactamases belonging to Ambler class D (CHDLs), including the oxacillinase family, OXA-23, OXA-24/40, OXA-58, OXA-143, OXA-235, and the intrinsic oxacillinase-encoding gene blaOXA-51-like associated with ISAba1. It can also be expressed by the production of Ambler class A β-lactamases (GES, KPC) and class B metallo-β-lactamases (MBLs) such as imipenemases (IMP), VIM, and NDM. 10 Among the MBLs, the NDM-type β-lactamases are the most emergent carbapenemases capable of hydrolyzing all β-lactams with the sole exception of aztreonam. To date, only NDM-1 and NDM-2 have been reported in A. baumannii. 10
P. aeruginosa is an opportunistic human pathogen implicated in a variety of acute and chronic infections, such as respiratory, urinary, and gastrointestinal tract infections as well as bacteremia. It is mainly found in immune-compromised patients suffering from, for example, cancer, HIV, and cystic fibrosis. This microorganism is one of the most important nosocomial pathogens and is responsible for infections with a high mortality rate. 11 P. aeruginosa is recognized by its intrinsic resistance to antibiotics and for its ability to acquire antibiotic resistance encoding genes. 12 In P. aeruginosa, carbapenem resistance can be triggered by enzymatic and non enzymatic mechanisms. Some early reports underlined the predominant role of outer membrane protein (Opr) in carbapenem-resistant P. aeruginosa phenotype (CRPA). 13 Different classes of carbapenemases have been reported in CRPA isolates around the world, mostly MBLs including VIM, IMP, AIM, SPM, GIM, SIM, DIM, and NDM, in addition to other enzymes encoded by different carbapenemase-producing genes such as KPC and OXA-type enzymes. 14 However, despite the increasing prevalence of MBL producing CRPA, mutational inactivation of OprD is recognized to be the major imipenem resistance mechanism in the absence of acquired carbapenemases. 15
This study aims to assess the antimicrobial susceptibility profile, and to explore and better understand the main mechanisms of carbapenem resistance in clinical isolates of A. baumannii and P. aeruginosa from clinical and environmental samples collected at Misurata Medical Center (MMC),Misurata, Libya.
Materials and Methods
Bacterial isolates
A total of 82 isolates were collected at MMC in Libya during May to June 2017. Among these, 34 were isolated from patients and 48 were recovered from hospital environment. All isolates were identified using the API 20E and the API 20NE strips (bioMérieux, Marcy-l'Étoile, France) for the Enterobacteriaceae and the non-Enterobacteriaceae isolates, respectively, and confirmed using matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry (MS; Microflex™; Bruker Daltonik, Bremen, Germany) with flex control and biotyper 3.0 software (Bruker Daltonik).
The clinical isolates were recovered from different sites (i.e., hands, noses, and wounds) and from various wards (i.e., newborn, orthopedic, ICU, neonatal ICU, and surgery departments). Furthermore, the environmental isolates were collected from diverse sampling sites (i.e., bed mattresses, warmer of baby incubator, mechanical ventilators, baby incubators, tables, and patient's tables) and from different wards (i.e., orthopedic, surgery, and ICU) (Table 1).
Phenotypic Features of 24 Non-Enterobacteriaceae Isolates Collected from Misrata Medical Center, Libya (Susceptible to Imipenem)
AK, amikacin; CAZ, ceftazidime; DO, doxycycline; F, nitrofurantoin; FEP, cefepime; FF, fosfomycin; ICU, intensive care unit; IMP, imipenem; MEM, meropenem; RA, rifampicin; SXT, trimethoprim/sulfamethoxazole; TIC, ticarcillin; TIM, ticarcillin/clavulanic acid; TOB, tobramycin; TZP, piperacillin/tazobactam.
Antibiotic susceptibility testing
Antimicrobial susceptibility tests were performed using the standard disk diffusion method on Mueller–Hinton agar as recommended by the Antibiogram Committee of the French Society for Microbiology/European Committee for Antimicrobial Susceptibility Testing (CA-SFM/EUCAST). Sixteen antibiotics were tested for each isolate from Enterobacteriaceae and non-Enterobacteriaceae. For Enterobacteriaceae, the antibiotics used were amoxicillin, amoxicillin/clavulanic acid, cefepime, piperacillin/tazobactam, cephalothin, ceftriaxone, imipenem, ertapenem, amikacin, gentamicin, trimethoprim/sulfamethoxazole, ciprofloxacin, fosfomycin, nitrofurantoin, and doxycycline. For non-Enterobacteriaceae, a range of antibiotics has also been tested: ticarcillin, ticarcillin/clavulanic acid, piperacillin/tazobactam, cefepime, ceftazidime, meropenem, imipenem, amikacin, tobramycin, ciprofloxacin, rifampicin, trimethoprim/sulfamethoxazole, fosfomycin, nitrofurantoin, and doxycycline (Bio-Rad, Marnes-la-Coquette, France). The isolates were considered resistant to imipenem if the diameter of the inhibition zone was <17 mm. For isolates with an inhibition zone diameter <17 mm, minimum inhibitory concentrations (MICs) of imipenem were determined by E-test (bioMérieux) and isolates were considered resistant when they had an imipenem MIC >8 μg/mL. The results were interpreted according to the CA-SFM breakpoints (2018).
Molecular detection and characterization of carbapenemase-encoding genes
Real-time polymerase chain reaction (PCR) and standard PCR were performed to screen for the presence of carbapenem hydrolyzing enzyme-encoding genes. Concerning A. baumannii isolates, carbapenemase-encoding genes were detected using specific primers for blaOXA-23, blaOXA-24, blaOXA-48, blaOXA-58, blaNDM, blaIMP, blaVIM, and blaKPC. The amplicons were purified and sequenced using the Big Dye terminator chemistry on an ABI 3130XL automated sequencer (Thermo Fisher Scientific, Waltham, MA). All sequences obtained were analyzed using BlastN and BlastP to search the NCBI database (www.ncbi.nlm.nih.gov/blast). 16 For P. aeruginosa isolates, carbapenemase-encoding genes were detected using specific primers for blaKPC, blaOXA-23, blaOXA-24, blaIMP, blaVIM, and blaNDM.
Molecular detection and characterization of the OprD-encoding gene
The amino acid changes of the protein OprD were inspected on imipenem-resistant P. aeruginosa isolates using specific primers as previously described. 17 Mutations were determined by comparison with the sequence of the P. aeruginosa PAO1 strain (GenBank accession no. CAA78448). 16
Genetic typing of carbapenemase-producing A. baumannii
To evaluate the clonal relatedness of the A. baumannii isolates, pulsed-field gel electrophoresis (PFGE) of the ApaI digested genomic DNA was performed using a CHEF-DRIII system (Bio-Rad, Hemel Hempstead, United Kingdom) as previously described. 18 The Info-Quest™FP v.4.5 software (Bio-Rad Laboratories) was used for dendrogram construction by the UPGMA (unweighted pair group method with arithmetic mean) method based on Dice's similarity coefficient. Isolates were considered to belong to the same PFGE cluster (pulsotype) if their Dice similarity index was ≥85%. 19
Genetic typing of carbapenemase-producing P. aeruginosa
Genotyping of all P. aeruginosa isolates by PFGE using of PstI digested genomic DNA was performed using CHEF-DRIII system (Bio-Rad, Hemel Hempstead, United Kingdom). According to the interpretative criteria of Tenover et al., 20 isolates were classified as indistinguishable, closely related, possibly related, or different. Indistinguishable isolates (no band differences) and closely related isolates (2–3 band differences) were considered to be the same genotype, whereas possibly related and different isolates (4–6 and >7 band differences, respectively) were considered different genotypes. 21
Results
Bacterial identification
A total of 82 Gram-negative bacteria were identified using conventional microbiological tests and MALDI-TOF MS. Enterobacteriaceae represented 57.31% of the isolates (No. of isolates = 47), including Klebsiella pneumoniae (39), Serratia marcescens (3), Providencia stuartii (3), Enterobacter cloacae (1), and Escherichia coli (1). Thirty-five isolates were identified as nonfermenter bacteria including A. baumannii (8), Acinetobacter pittii (1), Acinetobacter parvus (1), Acinetobacter schindleri (1), P. aeruginosa (20), Pseudomonas stutzeri (3), and Pseudomonas fluorescens (1). These isolates were recovered from clinical specimens and hospital environment. Among those, 34 isolates were recovered from hospitalized patients from different wards (orthopedic, surgery, neonatal ICU, and ICU), from various sites (wounds, noses, and hands) and 48 isolates were from hospital environment swabs (patient table, window handle, door, warmer of baby incubator, and bed mattress) (Tables 1 and 2).
Phenotypic and Genotypic Features of 11 Non-Enterobacteriaceae Carbapenem-Resistant Strains Collected from Misrata Medical Center, Libya
CIP, ciprofloxacin; IS, insertion sequence.
Isolates from hospitalized patients
A total of 34 isolates of clinical specimens were recovered from immune-compromised patients, the majority were from nasal cavities (22) followed by wounds (7) and hands (5) from different wards: newborn (6), orthopedic (5), ICU (8), neonatal ICU (4), and surgery (11).
As given in Supplementary Tables S1, the isolates showed a variable resistance pattern, with noted resistance to several antibiotic classes, including β-lactams, quinolones, and sulfonamides. The results showed that only one isolate of S. marcescens exhibited resistance to ertapenem, but being susceptible to imipenem.
Three isolates of A. baumannii and P. aeruginosa isolates presented a high level of carbapenemase activity with MICs of imipenem of 16 to >32 μg/mL. BlaOXA-23 gene was found in two isolates of A. baumannii recovered from nasal swabs in patients hospitalized in the ICU. None of the isolates harbored blaoxa-24, blaoxa-58, and blaoxa-48 (Table 2). For P. aeruginosa, VIM-encoding gene was detected only in one isolate recovered from epidermal cavity from patient admitted to the surgery ward. Sequencing revealed the presence of blaVIM-2 gene (Table 2). No other carbapenemase-encoding genes were detected among these isolates.
The amplification of OprD gene using specific primers was expected to have an amplicon of 1.332 bp, whereas a fragment with a size of 2.645 bp was produced in two isolates (Fig. 1). After purification and sequencing, it was shown that the OprD-encoding gene was disrupted at nucleotide position 180 pb by the insertion of a 1.197 bp fragment (Fig. 2). Our insertion sequence (IS) showed a 100% homology rate with ISPa25 according to BLASTN on NCBI GenBank. Indeed, one isolate carried modifications on their OprD-encoding gene sequence with a premature stop codon based on comparison with the sequence of the PAO1 reference strain (Table 2 and Fig. 2). We report here the co-occurrence of blaVIM-2 and oprD porin loss in one isolate of P. aeruginosa retrieved from patient's epidermal cavity admitted to the surgery ward.
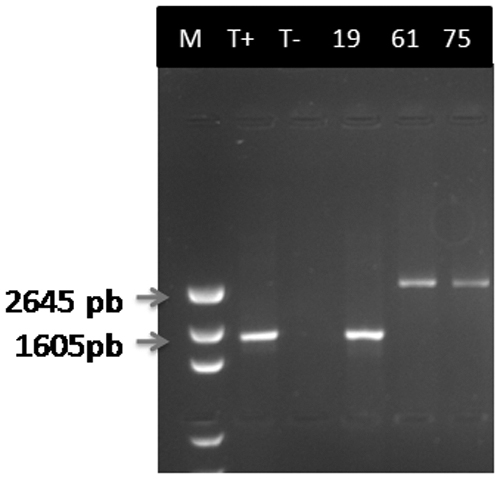
1.5% Agarose gel electrophoresis profile of the OprD-encoding gene detected in the three carbapenem-resistant Pseudomonas aeruginosa isolates.
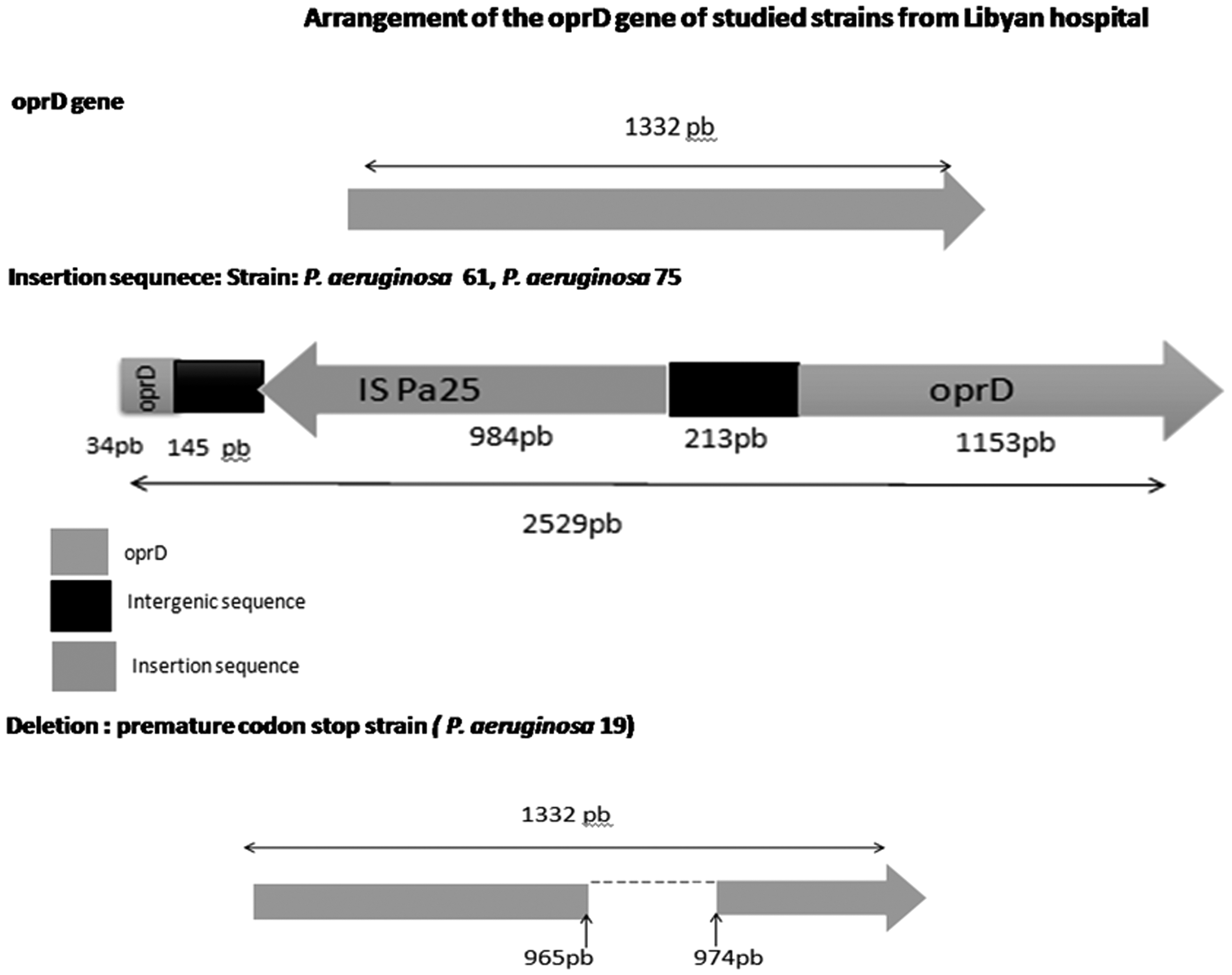
Schematic representation of the obtained OprD-encoding gene sequence compared with that of the Pseudomonas aeruginosa reference strain PAO1. (The two P. aeruginosa strains, 61 and 75, harbored the insertion sequence ISPa25; one P. aeruginosa isolate, assigned to the number 19, contained a premature stop codon.) IS, insertion sequence.
Hospital environment isolates
Identification of the 48 isolates recovered from the hospital environment performed by MALDI-TOF showed that 29 isolates were identified as Enterobacteriaceae represented by K. pneumoniae (27), P. stuartii (1), and E. cloacae (1). On the contrary, 19 nonfermenting isolates were found in the hospital environment and were as follows: A. baumannii (6), P. aeruginosa (9), P. stutzeri (3), and P. fluorescens (1). Hospital environmental isolates were collected from diverse sampling sites: bed mattresses, baby incubator warmer, mechanical ventilator, tables, and patient's tables from different wards: orthopedic, surgery, and ICU.
As described in Table 2, most of the isolates showed different patterns of resistance to several antibiotic classes such as β-lactams, quinolones, and sulfonamides. Contrariwise, amikacin was found to be the most active on all isolates.
Five isolates of A. baumannii were resistant to imipenem and showed high level of resistance to this antibiotic with MICs >32 μg/mL. These isolates were recovered from different wards (orthopedic, ICU, and newborn intensive care unit [NBICU]) from various sites (patient table, bed matters, baby incubator, and warmer of baby incubator); alternatively, none of the P. aeruginosa isolates were resistant to imipenem. Carbapenem resistance was mainly attributed to the carriage of the blaOXA-23 gene detected in three A. baumannii isolated from baby incubator and bed mattress from the neonatal ICU. However, NDM-1 was detected only in one isolate recovered from a patient table in orthopedic ward.
Genetic relatedness of carbapenem-resistant A. baumannii
PFGE profiles, under digestion with ApaI, of A. baumannii isolated from clinical and environmental settings are given in Fig. 3. Because of the high similarity between A. baumannii isolates in terms of resistance phenotypes, these isolates were selected for PFGE analysis. Genotypic analysis of A. baumannii identified four major PFGE patterns, named from A to E, which differed in migration of at least one DNA fragment and showed a similarity of at least 80% by dendrogram analysis. Pulsotypes A1, A2, and A3 were found to belong to one cluster with 94% similarity; these isolates were isolated from hospital environment of the newborn department. Pulsotype A1 and A2 showed 100% similarity and positive for OXA-23 despite being isolated from different sources (warmer of baby incubator and baby incubator). This cluster presents 76% similarity with pulsotype B, which includes one isolate from a patient in the surgery ward. On the contrary, pulsotypes C1 and C2 showed 100% similarity; C1 was OXA-23 producer isolate recovered from a bed mattress in the ICU, whereas C2 isolate was also OXA-23 producer but from a nasal swab from an ICU patient, which may reflect contamination of the bed mattress by patients secretions or nasal cannula.

PFGE-APaI pattern of the carbapenem-resistant Acinetobacter baumannii isolates and dendrogram profile. PFGE, pulsed-field gel electrophoresis.
Cluster D resembles C1 and C2 by 94% and belongs to the strain isolated from a patient, the isolate in cluster D was also isolated from a nasal swab from ICU patient and found OXA-23 producer.
Cluster E was different by 40% suggesting that an independent source is responsible for its dissemination; this cluster included an isolate producing NDM-1, swabbed from a patient table in the orthopedic ward in the hospital.
Genetic relatedness of carbapenem-resistant P. aeruginosa
In this work, the PFGE as an epidemiological tool allowed us to compare and identify the genetic relatedness of these isolates. Although this study had limitations in the low number of tested isolates (Fig. 4), genotypic analysis of P. aeruginosa identified three major PFGE patterns, namely F, G, and H. Based on the results of our study, pulsotypes are unrelated with 50% of homology between F and G and 40% between H, F, and G. Isolates were recovered from different patient's sites (hands and nasal swabs) from three different wards (orthopedic, ICU, and surgery). The hypothesis suggests that more than one clone are circulating and responsible for the infection.

PFGE-PstI pattern of the carbapenem-resistant Pseudomonas aeruginosa isolates and the corresponding dendrogram profile.
Discussion
Overall, carbapenem resistance has rapidly spread worldwide and the prevalence of imipenem-resistant isolates has reached a high percentage in some countries like Brazil, which reached 94% during 2014. 22 Carbapenem-resistant Gram-negative bacilli, A. baumannii and P. aeruginosa, are increasingly reported and may be difficult to eradicate. 23 The outbreak of carbapenem-resistant A. baumannii (CRAB) has been detected worldwide, including regions of the Mediterranean basin. 24 In the above study, the screening for OXA-type carbapenemases revealed that blaOXA-23-like was found in five isolates of CRAB, which is consistent with the two previous studies conducted in Libya (Mathlouthi et al. 25 ; Mathlouthi et al., 201626). However, in Tunisia, a neighbor country, several studies conducted by Charfi-Kessis et al., 27 Ben Cheikh et al., 28 and Dziri et al. 29 demonstrated the widespread blaOXA-23 carbapenemase-producing enzymes in different hospital settings within various geographic area (particularly in the Northern region). Our results showed that clinical specimens harboring blaOXA-23 were detected in two patients hospitalized in the surgery and ICU wards. These findings are in agreement with other studies describing that most prevalent OXA-23 gene in A. baumannii that was detected in ICU 30 wards even in Tunisia. 31
Furthermore, hospital environment provides an excellent ecological niche for the development of different microorganisms that could have clinical influence. 32 In our study, we found imipenem-resistant A. baumannii producing OXA-23 enzyme in the hospital environmental setting of two critical wards (ICU and NBICU), from different sites (bed mattress and warmer of baby incubator), suggesting that A. baumannii pathogen can survive in humid environments such as warmer of baby incubator and also in dry inanimate surfaces, such as bed mattress, and may constitute a great reservoir for the transmission of carbapenemases through horizontal transfer between patients and health care workers (HCWs). 33 It has been previously shown that contamination of the environment surrounding patients such as window handles, bed sheets, bed rails, beside, hands of HCWs, medication trolleys, and equipment might be the main source of A. baumannii transmission and outbreaks. 34 These findings can be explained by the higher carbapenemase activity of OXA-23 and/or the acquisition of carbapenem resistance through horizontal gene transfer. 35
In Libya, the circulation of NDM-1-producing A. baumannii isolates in inanimate surfaces has never been described and the source of NDM is not well documented. In Tunisia, a country that borders Libya, several studies were carried out on the occurrence of A. baumannii colonization and/or infections with the description of the molecular mechanisms of carbapenem resistance.28,36 However, in Libya, carbapenemase mechanisms are not well documented and only few recent studies have investigated such mechanisms 37 provided by the two predominant enzymes: oxacillinase OXA-23 and MBL NDM-1. 25 Our results show that none of the isolates harbored blaOXA-24, blaOXA-48, blaKPC, or blaIMP.
PCR assays targeting carbapenemase-encoding genes showed that one isolate recovered from patient's epidermal cavity at the surgery ward harbored VIM-2. This variant appeared in Marseille, France, in 1996. 5 Since then, VIM-2 had been spread as the predominant MBL variant among P. aeruginosa 38 in the Mediterranean area, in Spain, 39 Greece, 40 Italy, 41 Lebanon, 17 and even in various North African countries, such as Tunisia, 42 Algeria, 43 and recently in Morocco. 44 The study performed by Mathlouthi et al. 25 is the only study that highlighted the occurrence of blaVIM-2 in carbapenem-resistant P. aeruginosa from clinical specimens recovered from tracheal and wound samples from patients hospitalized in ICU and burn wards. Even in Tunisia, limited studies have been conducted on the resistance of carbapenems in P. aeruginosa.25,38,45
In addition, it has been shown that one of the most important mechanisms for imipenem resistance in P. aeruginosa is the absence of OprD production owing to insertions, mutations, and/or deletions in the OprD-encoding gene. In addition, numerous studies have described the presence of ISs truncating the OprD gene in CRPA. 46 These findings are in agreement with our results suggesting that the OprD-encoding gene was disrupted at nucleotide position 180 pb by an insertion of a 1,197 bp and this IS showed 100% homology with ISPa25. The presence of IS elements that disrupt the OprD gene and confer resistance to imipenem in clinical isolates of P. aeruginosa has been reported in many areas such as South Africa (ISPa26), 47 Spain (ISPa133), 48 the United States (ISPa8, 49 ISPa1635), and France (ISPa46). 50 Mobile genetic elements, such as IS can help Gram-negative bacteria to survive and adapt to altered environmental niches through the interruption of genes and genomic modifications.
In this study, the findings suggest a dynamic exchange of A. baumannii isolates between patients and their environmental surroundings. These pathogens can be transmitted from patient-to-patient, patient to a HCW, patient to environment and vice versa, which has in part been shown by the presence of the same pulsotype with high similarity. The strength of this study lies in the detailed molecular analysis of resistance mechanisms, whereas the small number of strains and their relatedness is a weakness. This investigation will give rise to future multicenter studies to obtain insight information into molecular epidemiology of A. baumannii isolates from different geographical regions in Libya. A plausible explanation that NDM-1-producing A. baumannii was isolated from patient table in orthopedic ward would be poor hand hygiene of HCWs. 51 In fact, these isolates producing NDM-1 and other MBLs were not related in timely manner.
Our finding suggests that no relationship between P. aeruginosa isolates and probably more than one clone is circulating and responsible for infection. However, as no environmental or staff sampling were performed, the source of contamination could not be assessed and further investigation should be performed.
Finally, education of cleaners and nursing staff, enhancing a frequent cleaning and disinfection of contaminated areas, decontamination of the patient's environmental surroundings may help to limit the spread of multidrug-resistant strains in the hospital environment and reduce the risk of an epidemic. Further studies are required to understand the molecular epidemiology of these organisms to develop strategies to combat their emergence and spread.
Footnotes
Acknowledgment
The authors thank Linda Hadjadj, a Technician in MEPHI (Microbes Evolution Phylogenie et Infections, Faculté de Médecine et de Pharmacie, Aix-Marseille-Université), for her grateful assistance.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was partly funded by IHU Méditerranée Infection, Aix-Marseille-University in France. This work was supported by the Tunisian Ministry of Higher Education and Scientific Research and Campus France Under the PHC-Utique project (Code 18G0819) which offered a scholarship to K.S. to attend the IHU Méditerranée Infection, Aix-Marseille-University in France.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.