
Abstract
Aims:
mcr-1 and blaNDM-1 co-harboring isolates have been reported, usually reside on different plasmids, suggesting co-transfer possibility of the two genes from separate donors to the same recipient strain. This study aims at screening and characterization of mcr-1 carrying Enterobacteriaceae in Northern China, and studying the transfer ability of mcr-1 alone and in company with blaNDM-1 from a second donor.
Results:
Three Escherichia coli strains and one Klebsiella pneumoniae strain carrying mcr-1 gene were screened out from 1992 isolates in our study. Co-existence of multiple resistance genes was found in the mcr-1-carrying strains, but none of them carried blaNDM-1. One E. coli demonstrated an single nucleotide polymorphism (SNP) (A-G) at -10 region of mcr-1, and one E. coli showed 2 SNPs (G-T and G-A) in the Shine-Dalgarno sequence-like region of mcr-1. The mcr-1 gene was located on plasmids of about 33–276 kb, and capable of transferring alone in three out of four mcr-1-positive isolates by conjugation. Co-transfer ability analysis demonstrated that mcr-1 from E. coli 13-68, which could not be transferred alone to E. coli C600, was successfully transferred in company with blaNDM-1 from K. pneumoniae ATCC BAA-2146.
Conclusions:
mcr-1 showed low incidence in our Enterobacteriaceae isolates. Co-transfer ability of mcr-1 and blaNDM-1 from separate donors provides direct evidence for the emergence of the mcr-1 and blaNDM-1 co-harboring isolates.
Introduction
Polymyxins, representing a family of cationic polypeptide antibiotics with broad-spectrum antimicrobial activities, are the last-resort antibiotics against serious infections caused by CRE (carbapenem-resistant Enterobacteriaceae). 1 The mechanism of colistin resistance is usually associated with two-component systems in which the modification of lipid A decreases its affinity to colistin. However, a plasmid carrying gene encoding colistin resistance (mcr-1) was reported in many species, including Escherichia coli and Klebsiella pneumoniae. 2
The mcr-1 gene, encoding a phosphoethanolamine transferase, was first identified in China among Enterobacteriaceae isolates and later disseminated worldwide. 3 The mcr-1 gene was found to be located on plasmids of different types, like IncH, IncI2, IncP, IncFIB, IncX4, IncA/c or IncFII plasmids. 4 The mcr-1 gene was reported to be flanked by complete ISApl1 region in some studies, but other studies showed it was flanked by either a truncated ISApl1 element without the tnpA gene or sequences without the whole ISApl1 element. 5 The mcr-1 cassette contains a 2600-bp-long DNA sequence, including a promoter region (called P1-mcr-1), the coding region of mcr-1 gene, and a 723-bp-long open reading frame (ORF) named orf723 encoding a hypothetical protein. The P1-mcr-1 was identified within the first 79 bp of the cassette, corresponding -35 (TGGATT) and -10 (TATAAT) boxes. 6 The orf723 gene was reported to be transported with the mcr-1 gene from its original genetic context, and did not impact colistin susceptibility. 7 A truncated product of mcr-1 gene (193 amino acids in length) was reported, which was induced by a frameshift caused by a 22-bp duplication of bases 503–525 of the mcr-1 ORF (GAACGCCACCACAGGCAGTAAA). 8 The five residues (E246, T285, H395, D465, and H466) were important for MCR-1 protein activity as critical residues for substrate binding of MCR-1. 9
mcr-1 and blaNDM-1 co-harboring isolates have been reported,10,11 which will cause ineffectivity of almost all antibiotics. The two genes usually reside on different plasmids, suggesting co-transfer possibility of the two genes from separate donors to the same recipient strain. The aim of our study was to screen and characterize mcr-1 carrying Enterobacteriaceae clinical isolates from our laboratory collections from 1998 to 2015 in Northern China (total 1992 isolates), and to evaluate the in vitro transfer ability of mcr-1 alone or together with blaNDM-1 from the second donor.
Materials and Methods
Bacterial strains and screening of mcr-1-producing isolates
A total of 1992 Enterobacteriaceae isolates randomly collected from six hospitals from 1998 to 2015 were screened, including E. coli (n = 911), K. pneumoniae (n = 831), Enterobacter cloacae (n = 189), and Enterobacter aerogenes (n = 61). Strains were identified to species level using the Vitek 2 system (bioMérieux, Marcy l'Etoile, France) and the mcr-1-carrying strains were further confirmed by 16S rRNA sequencing. The mcr-1 genes were detected by polymerase chain reaction (PCR) using the primer pair CLR5-F/R (Table 1). 3 The amplicons were subsequently sequenced with Sanger sequencing.
Primers Used for Polymerase Chain Reaction Amplification
The underline bases were the restriction enzyme cutting sites.
F, forward primer; PCR, polymerase chain reaction; R, reverse primer.
Susceptibility testing
The minimum inhibitory concentrations (MICs) of 20 antibiotics on the mcr-1 carrying isolates were determined according to the Clinical and Laboratory Standards Institute (CLSI) using broth microdilution method. 12 Ampicillin, amikacin, gentamicin, kanamycin, streptomycin, cefoxitin, cefuroxime, cefixime, cefotaxime, ceftazidime, ceftriaxone, cefepime, tigecycline, colistin, levofloxacin, aztreonam, imipenem, meropenem, and rifampicin were from National Institutes for Food and Drug Control (Beijing, China); polymyxin B was from Sangon Biotech (Shanghai, China).
Multilocus sequence typing and the plasmid incompatibility type
The isolates containing the mcr-1 gene were typed by assigning alleles and sequence types (STs) from the Institute Pasteur multilocus sequence typing (MLST) website (http://bigsdb.pasteur.fr/ecoli/ecoli.html for E. coli; http://bigsdb.pasteur.fr/klebsiella/klebsiella.html for K. pneumoniae) according to the previously described scheme. 16 The plasmid incompatibility groups (Inc) of the strains were analyzed by the published PCR method. 13
Pulsed-field gel electrophoresis and Southern blot hybridization
The relatedness of the mcr-1-carrying isolates was studied by pulsed-field gel electrophoresis (PFGE) with XbaI digestion, and the plasmid contents in the isolates were evaluated by PFGE with S1 nuclease treatment (S1-PFGE). Briefly, the total bacterial DNA was prepared in agarose blocks and digested with XbaI (Takara, China) for 3 hr at 37°C or S1 nuclease for 15 min at 23°C. The DNA fragments were separated by a CHEF DRII system (Bio-Rad, Hercules, CA) for 23 hr at 6 V/cm and 14°C, with initial and final pulses conducted for 5 and 20 sec for XbaI-PFGE and initial and final pulses conducted for 5 and 25 sec for S1-PFGE. Genomic DNA of the Salmonella enterica var. Braenderup H9812 strain digested with XbaI was used as the reference standard. For determination of mcr-1 locations on plasmids, plasmid DNAs were transferred to a positively charged nylon membrane (Millipore, Billerica, MD) after S1-PFGE, and mcr-1 was detected by Southern blot hybridization with a digoxigenin-labeled mcr-1-specific probe using the DIG High Prime DNA Labeling and Detection Starter Kit II (Roche Diagnostics).
Conjugation of mcr-1 plasmids
To study the transfer ability of mcr-1 gene in the strains, conjugation was conducted with E. coli C600 as the recipient. For mcr-1 alone conjugation, strains carrying mcr-1 were mixed with E. coli C600 at a ratio of 1:10 (v:v) using fresh overnight cultures. One hundred microliters of the mixed cultures was then dropped carefully on a cellulose filter membrane (pore size 0.45 μm) already placed on plain Mueller-Hinton (MH) agar plates, and incubated at 37°C for 20 hr. The mixed cultures were harvested from the filter membrane, and plated on MH agar plates supplemented with rifampin (500 mg/L) and colistin (3 mg/L). For mcr-1 and blaNDM-1 co-transfer analysis, fresh overnight cultures of strains producing mcr-1, blaNDM-1-producing strain K. pneumoniae ATCC BAA-2146 or E. coli 13-1, and recipient strain E. coli C600 were mixed at a ratio of 1:1:1 (v:v:v). One hundred microliters of the mixed cultures was then dropped carefully on a cellulose filter membrane (pore size 0.45 μm) already placed on plain MH agar plates, and incubated at 37°C for 20 hr. Transconjugants were selected on MH agar plates supplemented with rifampin (500 mg/L), meropenem (50 mg/L), and colistin (3 mg/L). Species identification, MIC determination, and resistance gene detection of the transconjugants were analyzed accordingly.
mcr-1 carrying plasmid sequencing
The plasmids in transconjugants Trc-E. coli 13-43 E. coli C600 and Trc-K. pneumoniae 09-20 E. coli C600 were extracted by alkaline lysis method, and subjected to plasmid sequencing using single-molecule real-time method by PacBio RSII platform in Beijing Genomics institution. The sequencing results were screened to remove the poor and repeated sequencing fragments, and genomic fragment contaminations from E. coli C600. The remaining plasmid fragment data were then used in sequence assembly, and Gene Ontology, Swiss-Prot, Cluster of Orthologous Groups of proteins, and Non-Redundant Protein Database were used to annotate the position and function of each coding region on the plasmids.
Antibiotic resistance gene detection and genetic environment analysis of mcr-1
The presence of antibiotic resistance genes with different categories was detected in the mcr-1-carrying isolates by PCR as previously described.14,15 The whole encoding regions, mcr-1 cassette, including promoter regions and ISApl1-mcr-1 sequence in the isolates, were amplified using primers mcr-1 2Fs F/R, ISApl1-mcr-1 1F/1R, and ISApl1-mcr-1 KL 1F/1R (Table 1), and sequenced. The genetic contexts of the mcr-1 gene in the five mcr-1-positive isolates were characterized by multiplex-PCR amplifications of nikB, tnpA, HP1, pilP, virD4, and virB4 as described previously.6,16
Quantification of mcr-1 genetic and expression levels by quantitative PCR
The genetic and expression levels of the mcr-1 in the five mcr-1-producing strains were measured by SYBR Green-based real-time PCR (Fast SYBR® Green Master Mix qPCR Kit; Applied Biosystems) with rpoB housekeeping gene as the internal reference. Specific primers mcr-1-q 2F/2R for the mcr-1 gene and rpoB-q4 F/4 R for the rpoB gene were designed (Table 1). Total DNAs of the five strains were extracted by the bacterial DNA extraction kit and total RNAs were isolated from the different strains using the RNAprep Pure Cell/Bacteria Kit (Tiangen Biotech, Beijing, China) as recommended by the manufacturers. First-strand synthesis of cDNA from RNA was performed in 25 μL volume by the Moloney Murine Leukemia Virus (M-MLV) Reverse Transcriptase (Cat#.M170A; Promega). The genetic levels were detected with the whole DNA as templates and the expression levels of mcr-1 were detected with the cDNA as templates. Cycling conditions were 95°C for 2 min and 40 cycles of 95°C for 3 sec, 60°C for 20 sec, and 72°C for 7 sec, followed by a ramp from 65°C to 95°C for melting analysis. The experiments were performed in triplicate on different days.
Results
Screening the mcr-1-producing strains
Four mcr-1-harboring isolates were identified: E. coli 08-85, E. coli 13-43, E. coli 13-68, and K. pneumoniae 09-20. The presence of the mcr-1 genes in the four strains was confirmed with the consensus sequences (GenBank: KP347127.1). The mcr-1 positivity was 0.33% (three strains) in E. coli and 0.12% (one strain) in K. pneumoniae.
Minimum inhibitory concentrations
The four mcr-1-positive isolates had MICs ranging from 4 to 16 mg/L for colistin and polymyxin B (Table 2). These strains also were resistant to other types of antibiotics, including quinolones, aminoglycosides, and cephalosporins. The three E. coli isolates were resistant to ampicillin, gentamicin, cefoxitin, cefuroxime, cefotaxime, ceftriaxone, cefixime, levofloxacin, and rifampicin. E. coli 08-85 was also resistant to cefepime and streptomycin; E. coli 13-43 was resistant to aztreonam and E. coli 13-68 was resistant to kanamycin and streptomycin as well. K. pneumoniae 09-20 was resistant to amikacin, levofloxacin, cefoxitin, kanamycin, and rifampin. All the four strains were susceptible to meropenem and imipenem.
The Minimum Inhibitory Concentrations of Different Antibiotics Against the mcr-1-Positive Strains
AMK, amikacin; AMP, ampicillin; ATM, aztreonam; CAZ, ceftazidime; CFM, cefixime; CRO, ceftriaxone; CST, colistin; CTX, cefotaxime; CXM, cefuroxime; FEP, cefepime; FOX, cefoxitin; GEN, gentamicin; IMP, imipenem; KAN, kanamycin; LVX, levofloxacin; MEM, meropenem; MIC, minimum inhibitory concentration; PMB, polymyxin B; RIF, rifampin; STR, streptomycin; TGC, tigecycline.
Antibiotic resistance-related genes
The antibiotic resistance gene profiles in the four mcr-1-positive strains are listed in Table 3. E. coli 08-85 carried aac(3)-IIc and blaCTX-M, encoding resistance to aminoglycosides and β-lactams. E. coli 13-43, E. coli 13-68, and K. pneumoniae 09-20 carried more resistance genes than E. coli 08-85, leading to resistance against quinolones, aminoglycosides, and β-lactams.
The Antibiotic Resistance Related Genes in of the mcr-1-Positive Strains
MLST, multilocus sequence typing.
MLST and the plasmid incompatibility type
All the three E. coli strains belonged to the same ST type (ST5), and K. pneumoniae 09-20 belonged to ST1. The plasmid incompatibility types were determined. E. coli 08-85 harbored three incompatibility-type plasmids: FIA, FIB, and FII. E. coli 13-43 harbored two incompatibility-type plasmids: P and FII. E. coli 13-68 harbored four incompatibility-type plasmids: HI2, FIB, N, and FII. The mcr-1-harboring plasmids belonged to FIIK incompatibility type in K. pneumoniae 09-20.
PFGE analysis and conjugation
PFGE results with XbaI digestion showed the relatedness of the four mcr-1-positive strains (Fig. 1A) and the S1-PFGE results demonstrated the plasmid contents of the studied strains (Fig. 1B). S1-PFGE results demonstrated diverse plasmid contents of the strains, E. coli 08-85 harbored a total of two plasmids: ∼60 and 140 kb; E. coli 13-43 harbored a total of two plasmids: ∼33 and 80 kb; E. coli 13-68 harbored a total of three plasmids: ∼100, 132, and 276 kb; and K. pneumoniae 09-20 harbored a total of four plasmids: ∼54, 62, 104, and 216 kb. Furthermore, Southern blot hybridization results after S1-PFGE showed that mcr-1 in the five strains were located on plasmids with sizes ranging from 33 to 276 kb (Fig. 1C).
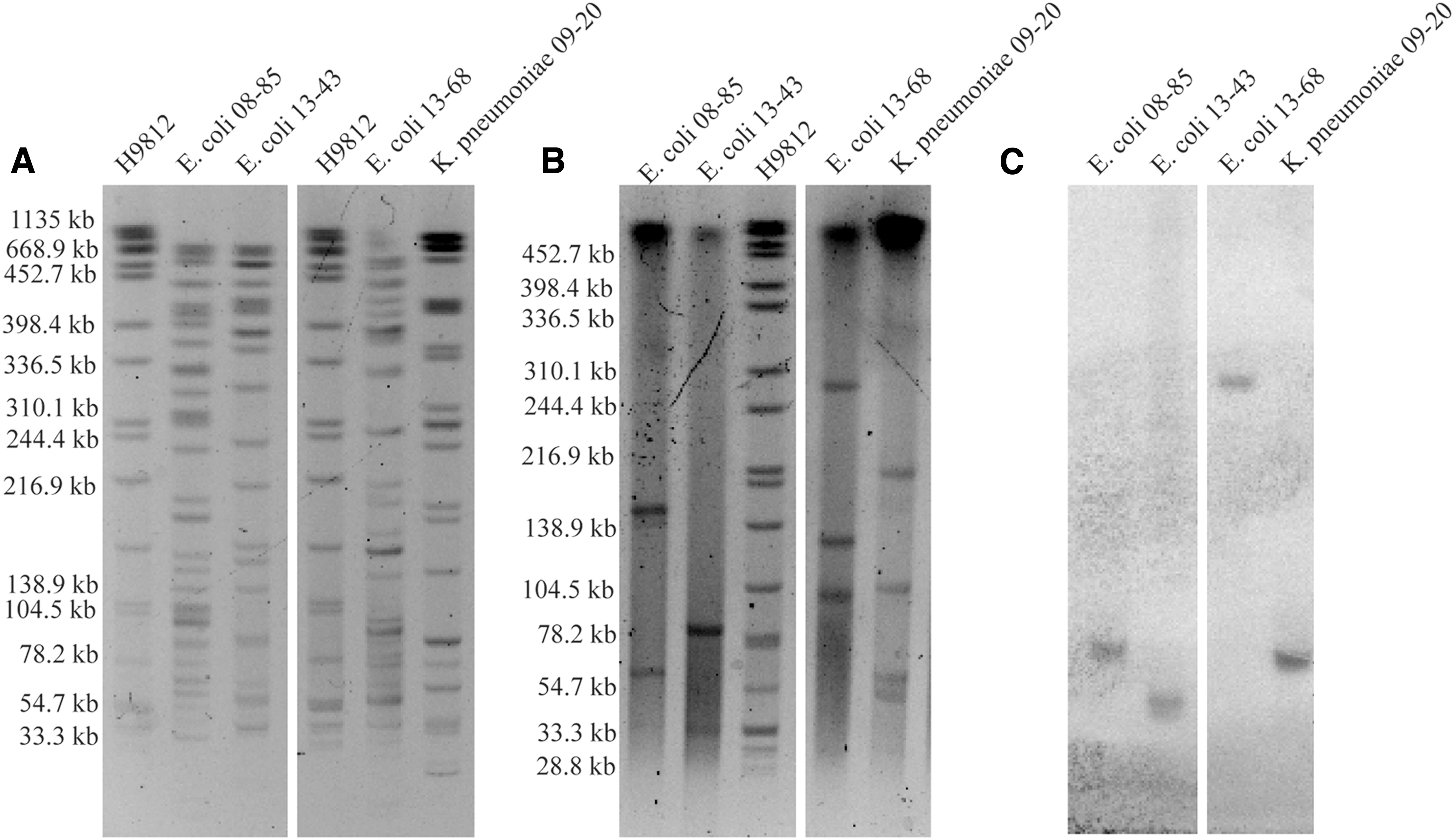
PFGE, S1-PFGE and Southern blot results.
Transfer ability of the mcr-1 gene from different strains was analyzed by conjugation with E. coli C600 as the recipient, and the results suggested that mcr-1 gene could be transferred from all the strains except E. coli 13-68 to E. coli C600. MIC determination of colistin and polymyxin B demonstrated at least fourfold increase in MICs against the transconjugants than those against the recipient isolate (Table 4). The presence of the corresponding plasmids in the representative transconjugants was confirmed by S1-PFGE (data not shown).
The Minimum Inhibitory Concentrations of Polymyxin B, Colistin, Meropenem, and Rifampin on the Transconjugants
Trc, transconjugant.
Co-transfer ability of mcr-1 together with blaNDM-1 from the second donor was evaluated using mcr-1-harboring strains as a first donor, and blaNDM-1-harboring strains K. pneumoniae ATCC BAA-2146 or E. coli 13-1 as a second donor, and E. coli C600 as a recipient. When K. pneumoniae ATCC BAA-2146 was used as the co-donor strain, the blaNDM-1 gene in K. pneumoniae ATCC BAA-2146 and the mcr-1 gene in mcr-1-producing strains, except E. coli 08-85, were successfully co-transferred to E. coli C600. What needs to be mentioned is that, mcr-1 gene from E. coli 13-68 isolate, which could not be transferred alone to E. coli C600 by conjugation, was successfully transferred in company with blaNDM-1 from K. pneumoniae BAA-2146 to E. coli C600. When E. coli 13-1 was used as the co-donor, only the mcr-1 gene in K. pneumoniae 09-20 was co-transferred with blaNDM-1. The mcr-1 gene and blaNDM-1 gene on different plasmids were successfully co-transferred to E. coli C600 strains as demonstrated by S1-PFGE (data not shown). The MICs of colistin, polymyxin B, and meropenem against the co-transconjugants were at least fourfold higher than the recipient strain (Table 4). The mcr-1 genes in single transconjugants and the blaNDM-1 genes of the co-transconjugants were stable, while the mcr-1 gene in the co-transconjugants lost after 25 consecutive days' transfer on plain MH medium.
mcr-1-carrying plasmid sequencing
The mcr-1 plasmids in Trc-E. coli 13-43 E. coli C600 and Trc-K. pneumoniae 09-20 E. coli C600 were determined to be 33.3 and 61.9 kb, respectively, by plasmid sequencing. No other reported resistance gene was found on the plasmids. However, genes related to toxin-antitoxin system and conjugal transfer were found, consistent with their transfer capability. The plasmid sequencing results have been deposited in GenBank with access numbers MG747473.1 and MG747472.1.
Encoding region and promoter region sequencing and genetic environment analysis of mcr-1
The complete mcr-1 cassette was identified in all isolates. ISApl1-mcr-1 mobile element was identified in E. coli 08-85, E. coli 13-68, and K. pneumoniae 09-20, suggesting the likelihood of ISApl1 carried from its original progenitor of mcr-1. The genetic environments surrounding mcr-1 in these strains are shown in Fig. 2A. All the strains have a HP1 gene downstream of mcr-1. In E. coli 08-85 and K. pneumoniae 09-20, the mcr-1 gene was surrounded by six genes (nikB, tnpA, HP1, pilP, virD4, and virB4), suggesting high similarity of mcr-1-carrying plasmids in these strains to pHNSHP45 from E. coli strain SHP45. 3 The mcr-1 gene was surrounded by tnpA and HP1 gene in E. coli 13-68. In contrast, HP1 gene was found to be the only gene surrounding mcr-1 gene in E. coli 13-43. One single nucleotide polymorphism (SNP) (A-G) locating at -10 region of mcr-1 was found in E. coli 13-43, and two SNPs (G-T, G-A) were found in the Shine-Dalgarno sequence-like region of mcr-1 in E. coli 13-68 (Fig. 2B).

The genetic environment of mcr-1 gene and the SNPs in the promoter regions.
The mcr-1 genetic and expression levels
The genetic and expression levels of mcr-1 in the four strains were analyzed by quantitative PCR, and the results are shown in Fig. 3. When compared to E. coli 08-85, the genetic levels of mcr-1 were similar in E. coli 13-43, while it was about 0.57-fold lower in E. coli 13-68 and 1.67-fold higher in K. pneumoniae 09-20. With the expression level of mcr-1 in E. coli 08-85 setting as 1, the expression levels of mcr-1 in E. coli 13-43, E. coli 13-68, and K. pneumoniae 09-20 were 0.40, 1.27, and 1.31, respectively.
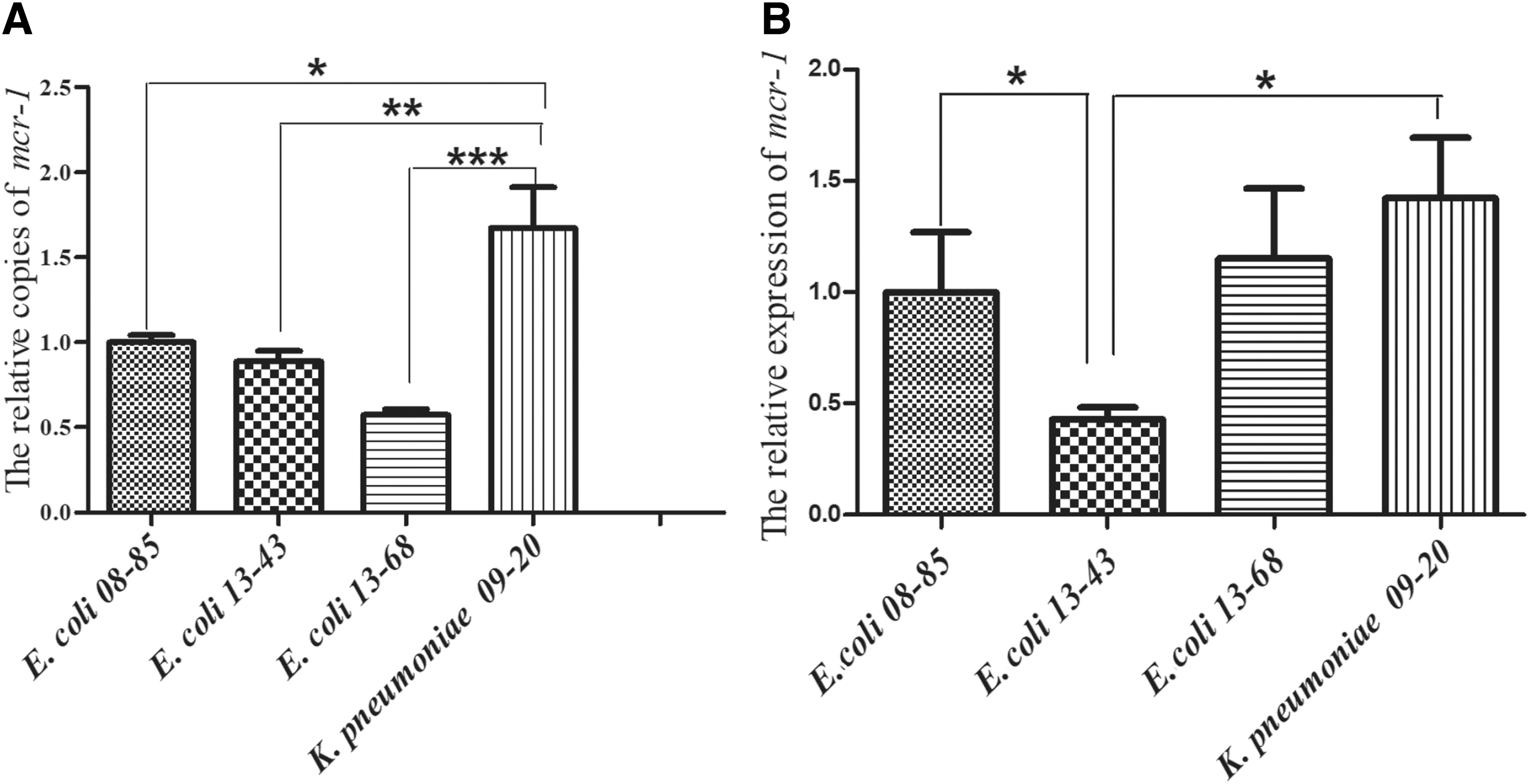
The relative genetic and expression levels of mcr-1 gene in the five clinical strains.
Discussion
The mcr-1 gene was first detected in three E. coli isolated in the 1980s in China, and the outbreak of mcr-1-harbouring E. coli from food-producing chicken started in 2009. 17 In the strain collection used in this study, the earliest strain carrying mcr-1 gene was isolated in 2008 (E. coli 08-85), in consistence with the time for increased appearance of mcr-1-carrying strain and colistin resistance after 2009. 18 The mcr-1 detection rates were 0.65% (2 out of 306) in E. coli and 0.36% (1 of 275) in K. pneumoniae when only strains isolated after 2009 were considered, lower than the reported 1% detection rate. 19 The mcr-1 gene usually resides on plasmids with various sizes, plasmid of 250 kb in a Salmonella isolate from pigs at slaughter (pHNSHP45-2), 20 plasmid of 33 kb in E. coli (pESTMCR), 21 and so on. In this study, mcr-1 was found on plasmids of ∼60 kb in E. coli 08-85, ∼33 kb in E. coli 13-43, ∼276 kb in E. coli 13-68, and ∼62 kb in K. pneumoniae 09-20.
The metallo-β-lactamase NDM-1 encoded by the blaNDM-1 gene inactivates all β-lactams except aztreonam. Most isolates harboring the blaNDM gene were only sensitive to tigecycline or colistin. 22 However, the function of mcr-1 could complement the function of the blaNDM and expand the resistance profiles to colistin. According to previous reports, mcr-1-harboring strains can also co-express blaNDM genes, including blaNDM-1, blaNDM-5, and blaNDM-9.4,23–26 One isolate with blaNDM-1 and mcr-1 residing on two different plasmids was isolated in China. 27 In this study, blaNDM-1 gene was not detected in the four mcr-1-carrying isolates, and a survey of 10 blaNDM-1-positive Enterobacteriaceae isolates in our laboratory did not find the co-expression of mcr-1 with blaNDM-1. However, two donor-one recipient conjugation with blaNDM-1-carrying strain and mcr-1-carrying strain as the donors demonstrated co-transfer of the two genes to the same recipient strain, which might be the genetic foundation of co-expression of mcr-1 and blaNDM-1 in one strain found in clinic. Importantly, the plasmid encoding mcr-1 in E. coli 13-68, which might not transfer alone to the recipient strain, could transfer to the recipient strain with the help of blaNDM-1-positive K. pneumoniae ATCC BAA-2146, suggesting the complementation between strains in resistance genes as well as in transfer equipments.
Five SNPs in the motif-forming residues of mcr-1 were reported to be essential for the function of MCR-1, and possibly through influence on the binding of the enzyme to its co-factor of zinc ions. 9 A single SNP in the promoter of mcr-1 (-36) was reported, and may affect mcr-1 expression. 8 In this study, we demonstrated that even though there were no mutations in the mcr-1-encoding regions, one SNP located at -10 region of mcr-1 in E. coli 13-43 and two SNPs in the Shine-Dalgarno (SD) sequence-like region of mcr-1 in E. coli 13-68 were detected. The mutations in the promoter regions may be related to the lowered expression levels of mcr-1 in E. coli 13-43 in comparison to E. coli 08-85, as the mcr-1 genetic levels were similar. The two SNPs in the SD sequence-region of mcr-1 might cause an increase in mcr-1 sequence in E. coli 13-68, considering the slightly higher expression of mcr-1 in the strain when the genetic level of mcr-1 was about half fold lower in comparison to E. coli 08-85. Colistin resistance was reported to be not affected by the number of mcr-1-carrying plasmids harbored by the strains, 21 but mainly related to the chemical modification of the bacterial lipid A. 9 Therefore, the capacity of colistin resistance may only be related to whether the mcr-1 gene is expressed or not, but not to the quantity of expression. In consistent with these findings, even though the mcr-1 expression levels of the three E. coli strains differed, the MICs of colistin against the strains were similar. It has been speculated that particular genetic features of ISApl1-mcr-1 mobile element could be the origin of wide diffusion of mcr-1. 18 In our study, complete ISApl1-mcr-1 mobile element was presented in three out of four strains.
In conclusion, four mcr-1-carrying Enterobacteriaceae isolates were identified from our laboratory collections in Northern China from 1998 to 2015 (totally 1993 isolates), with detection rate of about 0.2%. The co-transfer of blaNDM-1 and mcr-1 from two donors to one recipient strain provided direct evidence for the emergence of blaNDM-1 and mcr-1 co-harboring isolates.
Footnotes
Acknowledgment
We would like to acknowledge Shuai Bai from Jinggangshan University for his help in screening of mcr-1-producing isolates.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (Grant No. 81573475, 81621064), CAMS Initiative for Innovative Medicine (Grant No. 2016-I2M-3-014), and National Mega-project for Innovative Drugs (Grant No. 2019ZX09721001).