
Abstract
Objective:
This study's main objective was to analyze the antibiotic susceptibility profile of Escherichia coli isolates obtained from a fecal sample of a captive Baird's tapir (Tapirus bairdii) in Costa Rica.
Materials and Methods:
The fecal sample was collected inside the enclosure on March 3, 2017, right after the animal defecated. Samples were cultured on MacConkey agar plates nonsupplemented and supplemented with 2 μg/mL of cefotaxime. Bacterial identification and antibiotic susceptibility were performed with the Vitek 2 Compact System and the Kirby Bauer disk diffusion method, respectively. Polymerase chain reaction amplification was performed to detect blaCTX-M beta-lactamase genes. Resistant isolates were subjected to whole-genome sequencing (WGS).
Results:
After evaluating several antibiotic classes, a multidrug-resistant E. coli strain with extended-spectrum beta-lactamase phenotype was isolated. Resistance to cefotaxime, cefepime, ampicillin, ampicillin/sulbactam, and tetracycline was detected. WGS analysis showed the presence of blaCTX-M-1, blaTEM-1B, and tet(B) genes. The presence of IncN plasmids and Col156 was also detected.
Conclusion:
Our findings are according with the notion that animals' high density enhances the spread of resistant determinants in a captive environment in a limited space, where the likelihood of direct or indirect contact with other animals and humans is more frequent.
Introduction
Tapirs (Tapirus spp.), as strict herbivores, play an essential ecological role as an ecosystem engineer, mainly through efficient long-distance seed dispersal and selective browsing, the reason why they are called the “gardeners of the forest” contributing to mitigate climate change.1–3 Currently, Baird's tapir (Tapirus bairdii) is classified as an endangered species, according to International Union for Conservation of Nature. 4 With <5,000 living individuals in all its distribution range, due to threats such as habitat fragmentation, hunting, and recently vehicle collisions.4–6
It has been calculated that <1,500 free-ranging individuals inhabit Costa Rica. 5 Approximately 15 captive tapirs are estimated in the country (A. Sánchez-Núñez [Wildlife Coordinator, National System of Conservation Areas, SINAC], personal communication). There are several reasons why some tapirs are held in captivity in Costa Rica. However, most of the time, these are animals that have been captured to receive medical attention, and after treatment, they are not good release candidates. Despite the reasons they are held in confinement, little is known about captive tapirs' health status in Costa Rica. Nevertheless, it is known that the systematic collection of data related to their health status is essential to develop conservation strategies for this endangered species in the wild. 7
The characterization of the antibiotic resistome of captive wild animals is not commonly performed as a part of the health status assessment; however, several reports support the notion that animals in captivity are more likely to acquire resistant determinants. 8 This is relevant not only because these individuals may develop infections with antibiotic-resistant bacteria but also because they may act as reservoirs. When these animals are eventually released, there is a potential risk of transmission and dissemination of resistance determinants throughout the environment as well as other animals or humans.8–10
Escherichia coli constitutes a high percentage of the large intestine microbiota from mammals. It is a reservoir for antibiotic resistance genes, which may have an essential role in the epidemiology of resistance determinants spillover.8,10–12 Given that E. coli easily become resistant due to point mutations or horizontal transfer of genetic material, these bacteria have been used to measure antimicrobial resistance prevalence. 11 Among the resistant mechanisms in Gram-negative (GN) bacteria, E. coli producing extended-spectrum beta-lactamases (ESBL), mainly from the CTX-M type, are considered an exciting topic in public health since they have caused many outbreaks and infections in humans worldwide. 13
We previously reported 98% (59/60) of pansusceptible E. coli isolates from fecal samples free-ranging Baird's tapirs in Costa Rica. 14 Conversely, multidrug-resistant E. coli from lung and fecal isolates were reported in captive South American tapirs (Tapirus terrestris) in Colombia and the Czech Republic, respectively.15,16 Besides multidrug-resistant phenotype, Klimes et al. also detected an E. coli producing ESBL with the phenotype CTX-M-15 in the captive South American tapir. 16 In addition, phenotype CTX-M-1 was recently isolated in the same species in captivity. 17
In this study, we report the isolation of a fecal ESBL-producing E. coli in a captive Baird's tapir (T. bairdii) from Costa Rica and its subsequent genetic characterization using WGS. To our knowledge, this is the first report related to the detection of a multidrug-resistant E. coli harboring ESBL of the CTX-M-1 class in a fecal sample of a captive Baird's tapir in Costa Rica, Central America.
Materials and Methods
Sample collection
A female captive tapir (weighing ∼250 kg) had biological, nutritional, and veterinary care. The enclosure where the female was held, consisting of an area of ∼600 m2, with several trees, a small water tank (8 m2) and next to it, several enclosures with different animal species such as coyotes (Canis latrans), red fox (Urocyon cinereoargenteus), collared peccaries (Tayassu tajacu), puma (Puma concolor), mantled howler monkey (Alouatta palliata), and spider monkey (Ateles geoffroyi), among others.
During a routine physical examination, the animal presented a good physical condition, no lesions in the eyes, skin, or hooves. However, near a month before examination, the animal had been orally treated with ceftiofur (5 mg/kg per 300 kg) due to a submandibular abscess of unknown etiology. For economic reasons, the oral treatment was suspended and changed to topical. After the physical examination, two fecal samples were collected inside the enclosure on March 3, 2017, right after the animal defecated. After collection, samples were stored at 4°C and transported to the Bacteriology Laboratory of the Escuela de Medicina Veterinaria, Universidad Nacional de Costa Rica, for analysis.
E. coli isolation
Samples were directly cultured on standard MacConkey (Oxoid, Hampshire) agar plates nonsupplemented and supplemented with 2 μg/mL of cefotaxime (Sigma-Aldrich, MO) and incubated for 24 hr at 37°C. Typical lactose positive E. coli colonies were identified with Vitek Compact 2 automated system (BioMèriux, Marcy-l'Étoile) using the GN card. E. coli ATCC 25922 (cefotaxime susceptible) and Klebsiella pneumoniae ATCC 700603 (cefotaxime resistant, ESBL positive) strains were used for quality control of the culture media.
Antimicrobial susceptibility testing
The antibiotic susceptibility test was performed with Vitek Compact 2 automated system (BioMèriux) using the AST-N279 card that includes the following antibiotics: amikacin, gentamicin, cefotaxime, ceftazidime, cefepime, ampicillin, piperacillin/tazobactam, ampicillin/sulbactam, imipenem, meropenem, ciprofloxacin, nalidixic acid, sulfonamide/trimethoprim, colistin, and nitrofurantoin. Susceptibility to tetracycline (30 μg) (Oxoid, Hampshire) was tested by Kirby Bauer disk diffusion method. M100 from CLSI (2020) was used for interpretation. 18
Detection of ESBL genes
ESBL screening test was performed using the combination disk test with cefotaxime (30 μg) and ceftazidime (30 μg) disks with and without clavulanate (10 μg). 14 CTX-M detection was performed using the methodology previously described. 19 In brief, CTX-M polymerase chain reaction (PCR) amplification of blaCTX-M alleles was carried out with primers CTX-MU1 (5′-ATG TGC AGY ACC AGT AAR GT-3′) and CTX-MU2 (5′-TGG GTR AAR TAR GTS ACC AGA-3′). These PCR primers allow the amplification of a 593 bp fragment present in different types of CTX-M, including blaCTX-M-1 to blaCTX-M-30.
Whole-genome sequencing analysis
An isolate with an ESBL phenotype was subjected to whole-genome sequencing (WGS). WGS was performed using Illumina next-generation sequencing. Reads quality was checked with FastQC and genome assembly with Unicycler, 20 with assembly quality check using Quast 21 and annotation with Prokka. 22 The Serotype was determined using Short Read Sequencing Typing (SRST2) tool 23 against EcOH database from Holt lab. Clermont Phylogroup was determined using the In Silico Clermont Phylotyper tool. Antimicrobial Resistance Identification By Assembly (ARIBA) tool 24 was used for multilocus sequence typing (MLST) analysis according to the Achtman scheme and with the ResFinder tool 25 reference database to identify resistance genes. Point mutation resistance was investigated using AMRFinderPlus 26 and plasmid detection and typing using PlasmidFinder. 27
Results
The culture of the tapir's fecal sample yielded one multidrug-resistant E. coli isolate. This E. coli isolate was resistant to cefotaxime, cefepime, ampicillin, ampicillin/sulbactam, and tetracycline (Table 1).
Features of E. coli Isolate Harboring Extended-Spectrum Beta-Lactamase and Tetracycline Resistance
AMP, ampicillin; CTX, cefotaxime; FEP, cefepime; MLST, multilocus sequence typing; SAM, ampicillin/sulbactam; TET, tetracycline.
Owing to the resistance against third-generation cephalosporins and a positive ESBL test (using the combination disk test), we performed a PCR toward blaCTX-M genes. The presence of a blaCTX-M gene was detected (Fig. 1). This multidrug-resistant isolate, harboring the gene encoding the production of the ESBL, was subjected to WGS to determine the serotype, the MLST sequence type, the Clermont phylogroup and the presence or absence of resistance genes, resistance point mutations, and plasmids (Table 1).
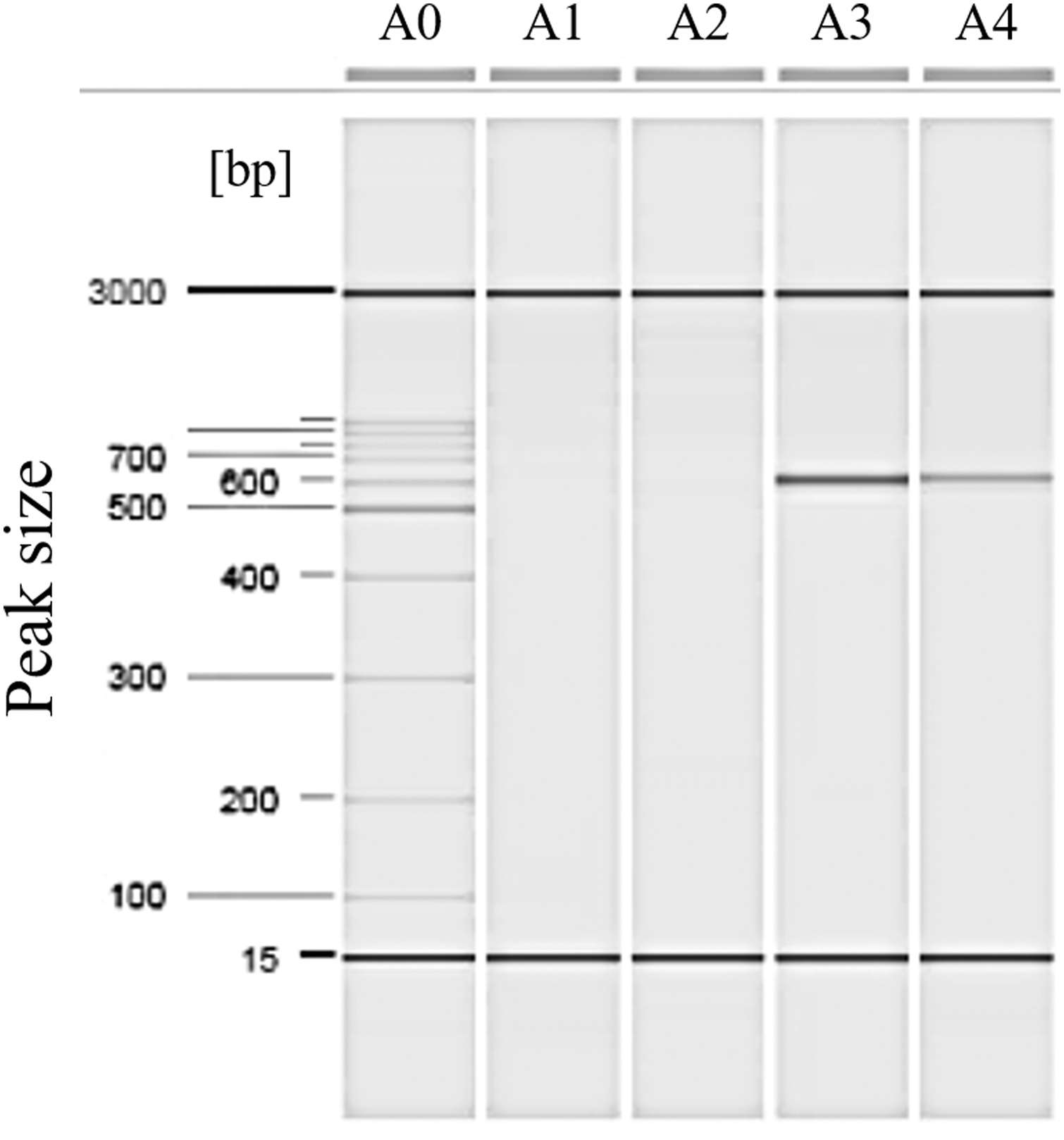
Detection of ESBL type CTX-M. A0: molecular weight marker, A1: reaction mixture control, A2: negative control (E. coli ATCC 25922), A3: positive control (Shigella flexneri OPS 187), A4: E. coli isolate H024B. ESBL, extended-spectrum beta-lactamase.
The detection of resistance genes blaCTX-M-1, blaTEM-1b, mdfA, tet(B) (using the ResFinder tool) confirmed the phenotypic resistance to cefotaxime, cefepime, ampicillin, ampicillin/sulbactam, tetracycline, and the ESBL phenotype. We also detected additional resistance genes to the aminoglycosides group [aph(3″)-Ia, aph(6)-id] (Fig. 1). The aminoglycosides gentamicin and amikacin were classified as susceptible by phenotypic methods. This susceptibility is expected since [aph(3″)-Ia ot aph(6)-id] do not confer resistance to these two antibiotics. 28 No resistance-related point mutations were found for any antibiotic class.
The serotype and MLST sequence type determined by WGS showed that this E. coli isolate belonged to the sequence type 58 (ST58) and the serotype O08:H21, respectively (Table 1). The conjugative plasmids Col156 and IncN were also detected (Table 1).
Discussion
In this study, we report a multidrug-resistant ST58 E. coli harboring an ESBL of the CTX-M-1 class in a fecal sample of a captive Baird's tapir (T. bairdii) in Costa Rica. E. coli harboring ESBL has been increasingly reported in several wildlife species in the past two decades in wild and captive conditions, including reports in >30 species shedding ESBL in diverse geographical landscapes.10,29 Although resistant bacteria seem to correlate with anthropogenic activity, some studies report the prevalence of ESBL in pristine habitats.30–32 In addition, some of the driving factors for the occurrence of resistant bacteria in wildlife are not fully understood; current evidence suggests that species' biology and ecology, including feeding and nesting near human-dominated landscapes, are among the key factors.29,33
In contrast, captive animals are in close contact with humans. Food manipulation, runoff, disinfectant use, and frequent antibiotic therapy are suggested to be the main drivers for resistant bacteria transmission (i.e., mainly through horizontal gene transfer mechanisms) in these facilities.29,31,34
Similar to human and veterinarian medicine, the most detected type of ESBL in wild animals corresponds to the blaCTX-M gene-family. Most of the studies conducted have identified blaCTX-M-1, blaCTX-M-2, blaCTX-M-9, blaCTX-M-13, blaCTX-M-14, blaCTX-M-15, blaCTX-M-32, blaCTX-M-55, and blaCTX-M-65.10,12,16,17 In addition to the blaCTX-M gene-family also the blaSHV-12 and blaTEM-52 were identified. 10 The most commonly reported non-ESBL phenotypes showed resistance to ampicillin, streptomycin, and tetracycline. 10
E. coli phylogenetic analysis was formerly based on a triplex PCR determining four main groups (A, B1, B2, and D). 35 The expanding body of multilocus sequence data and genome data for E. coli has contributed significantly to the better understanding of phylogroups for this bacteria into eight groups (A, B1, B2, C, D, E, F, belonging to E. coli sensu stricto; and Escherichia cryptic clade I). 36 Specifically, phylogenetic group B1 is often detected in E. coli isolates from captive animals, including Giant anteater (Myrmecophaga tridactyla), Geoffroy's cat (Leopardus geoffroyi), ocelot (Leopardus pardalis), and South American tapir (T. terrestris).17,37
ST58 has been reported as one of the main ESBL-producing E. coli circulating at the human–animal-environment 38 ; however, there are very few reports of ST58 in wildlife. 10 ST58 was previously reported in a fatal case of a captive critically endangered Brazilian merganser (Mergus octosetaceus) in several organs 34 and in peri-urban South American coatis (Nasua nasua) in Brazil. 38
IncN has been associated with disseminating several blaCTX-M variants, such as blaCTX-M-1, blaCTX-M-3, blaCTX-M-15, blaCTX-M-32, blaCTX-M-40, and blaCTX-M-65. 39 In several European countries, IncN plasmids largely contributed to the spread of blaCTX-M-1 among humans, animals, and the environment. 40 To our knowledge, the only reports related to IncN plasmids in wild animals were performed in European free-ranging waterbirds,41,42 in which they detected IncN and IncI1 harboring blaCTX-M-1 and IncI1 and IncFIA-FIB harboring blaCTX-M-15. In both surveys, they also detected the genes blaTEM-1B and tet(B). The broad-host-range IncN plasmids might contribute to the spillover of isolates producing CTX-M-1-like enzymes among other members of Enterobacteriaceae. 41
The detection of an IncN plasmid in the fecal microbiota of a wild animal held in captivity is considered a public health concern. 10 This individual may act as a reservoir, contributing to this resistance determinant's potential dissemination throughout the facilities and the environment.9,11 The possible source for the acquisition of these resistance determinants in this tapir is unknown. It is relevant to note that, in addition to the frequent contact with the staff that takes care of the animals in the facilities, its enclosure locates near several species' enclosures. This species included coyotes (C. latrans), red fox (U. cinereoargenteus), collared peccaries (T. tajacu), puma (P. concolor), mantled howler monkey (A. palliata), and spider monkey (A. geoffroyi), where water runoff occurs during rain or cleaning of the enclosures. Besides, many wild bird species settled or moved around the facilities that might function as vectors or reservoirs of resistance determinants. Therefore, close contact between humans and animals nearby occurs regularly and can disseminate antibiotic resistance genes. The isolation of a multidrug-resistant E. coli harboring ESBL contrasts with our previous results in free-ranging Baird's tapirs, in which 98% of E. coli isolates obtained from fecal samples were pansusceptible. 14
To monitor the animals' health status, we propose monitoring the fecal resistome to prevent resistance determinants spillover in the facilities or in animal candidates to be released. Our findings concern veterinarians to be aware of possible complications in treating captive animals infected with resistant bacteria.9,10
Conclusion
Our results provide evidence that hygienic practices, appropriate and responsible use of antibiotics in the rehabilitation of wild animals is essential when releasing them into the wild to prevent the dissemination of resistant determinants into the environment. 9 These findings support the notion that places with a high density of animals in a relatively small area promote high dissemination of resistance determinants, mainly through water runoff or direct or indirect contact among humans and animals.
Ethical Approval
The study was performed under the approval of the National Environment Ministry (MINAE), and the National System of Conservation Areas (SINAC), under the research permit (SINAC-SE-CUS-PI-R-021-2017), with the corresponding scientific passport (N° SINAC-SE-023-2017). Sampling processing was performed under the approval of the National Commission for Biodiversity Management (CONAGEBIO) under the research permit Project ID 431.
Footnotes
Acknowledgments
The authors thank Marcela Suárez, Tomás Poklepovich, and Josefina Campos for their technical support during WGS. We also want to thank the staff from the captive facilities where the tapir was held; Paloma Alcázar; the staff from Costa Rica Wildlife Foundation/Nai Conservation; and the staff from the Bacteriology Laboratory of the Universidad Nacional de Costa Rica (Heredia, Costa Rica).
Disclosure Statement
No competing financial interests exist.
Funding Information
The study was funded by the Bacteriology Laboratory of the School of Veterinary Medicine and by the Fondo para el Fortalecimiento de las Capacidades Estudiantiles (FOCAES-UNA-VI-OFIC-212-2017) of the Universidad Nacional de Costa Rica.