
Abstract
Bronchiectasis, which is an abnormal and irreversible dilation of one or several bronchial segments, causes significant morbidity and impaired quality of life to patients, mainly as the result of recurrent and chronic respiratory infections. Staphylococcus aureus is a microorganism known for its high infectious potential related to the production of molecules with great pathogenic power, such as enzymes, toxins, adhesins, and biofilm, which determine the degree of severity of systemic symptoms and can induce exacerbated immune response. This review highlighted the clinical significance of S. aureus colonization/infection in bronchiectasis patients, since little is known about it, despite its increasing frequency of isolation and potential serious morbidity.
Introduction
Bronchiectasis is the dilation and destruction of large bronchi caused by chronic infection and inflammation. Common causes are cystic fibrosis (CF), immune defects, and recurrent infections, although, in some cases, it may have unknown causes. 1
Continuous inflammation alters the anatomy of the airways and causes chronic cough and purulent sputum, whereas some patients may also experience fever and dyspnea. 2 Treatment and prevention of acute exacerbations include bronchial dilators, elimination of secretions, and antibiotics therapy in case of complications resulting from lung injuries due to resistant or opportunistic infections. 3
Selective predilection of Staphylococcus aureus in respiratory infections is still unclear. However, it is known that S. aureus adheres more intensely to hair cells, squamous nasal and oral epithelial cells. 4 Thus, an increase in viscosity of airway secretions and change in mucociliary function in bronchiectasis patients may be conditions that lead to S. aureus colonization. 5 Biofilm production is one of the strategies that results in the generation of isolates protected by a dense layer of extracellular matrix, which protects bacterial cells against the activity of both the immune system and antimicrobials. 6
In addition to adherence, S. aureus exhibits several other virulence factors, such as the production of leukocidins, hemolysins, catalase, coagulase, protein A, and many other toxins that contribute to its pathogenicity and enable it to escape from the host's immune system. 7
Regarding toxins, this study highlights superantigens (SAgs), which are exotoxins resulting from products of bacterial metabolism. They damage respiratory mucous membranes and, over time, also contribute to respiratory disease due to the host's exacerbated inflammatory response. 8
It has recently been observed that intense antibiotic therapy against bacterial infections of the airways is one of the procedures that most dramatically increase patients' life expectancy. 9 However, frequent use of antibiotics by bronchiectasis patients leads to an increase in bacterial resistance. Regarding S. aureus, the emergence of methicillin-resistant S. aureus (MRSA) isolates encoded by the mecA gene stands out, since they lead to resistance to all beta-lactam antibiotics. The mecA gene is in a mobile genetic element (Staphylococcal Cassette Chromosome mec, SCCmec) that can carry resistance genes to other classes of drugs, configuring the multidrug-resistant microorganism. 10
Despite its increasing frequency of isolation and potential serious morbidity, the clinical significance of S. aureus colonization/infection in bronchiectasis patients has been poorly studied. Thus, this review was triggered by the need to know more about the importance of S. aureus in this pathology.
Bronchiectasis
Bronchiectasis is one of the main chronic lung diseases characterized by inflammation and irreversible bronchial dilation. Interaction among chronic infection, exacerbations, and inflammation leads to a vicious cycle and results in lung damage to bronchi and parenchyma. 11
A severe inflammatory reaction in bronchi favors accumulation of viscous secretions to the point that they obstruct the airways completely and, consequently, dilate obstructed bronchi. Thus, the combination of obstructive conditions and high mucus production, associated with inflammatory response, results in accumulation of large amounts of secretion and interferes with the mucociliary function, a fact that prevents the adequate expiratory air flow that enables individuals to cough effectively and therefore remove residues. 12
The disease may be related to both congenital conditions, such as CF and anatomical defects of the bronchial tree, and acquired ones, whose cause may not have been identified. Therefore, the hypothesis is that this clinical picture has arisen as the consequence of environmental aggressors and susceptible hosts, which are conducive to impaired pulmonary defense mechanisms, in addition to being associated with concomitant injury to small and large airways. 13 Bronchiectasis is characterized by exacerbation and recurrent pulmonary infection with high morbidity and poor quality of life, and its prevalence has been associated with geographic, ethnic, and age factors.14,15
The first international guideline on the diagnosis and treatment of non-CF bronchiectasis was published in 2010. 16 However, in Brazil, the first systematic review on the topic was only published in 2019. It aimed at systematizing knowledge that had been accumulated on the subject. 17
A few years ago, bronchiectasis not related to CF was considered an “orphan disease” because it represented an unusual clinical condition, which remained under diagnosed due to lack of knowledge about its pathophysiological and etiopathogenic mechanisms. An increase in knowledge of mechanisms and characteristics of the disease (airway dilation and recurrent infections) has currently enabled to develop an efficient diagnostic and therapeutic approach, especially due to the dissemination of chest tomography, which has substantially contributed to the most accurate diagnosis of the disease. 17
Microbiome in Non-CF Bronchiectasis Patients
Colonization of airways in suppurative lung diseases, such as bronchiectasis, plays a fundamental role in the evolution of their clinical manifestations. Therefore, understanding the role of the microbiome is fundamental to understanding their pathophysiology. 18
Most bacteria that cause lung inflammation in bronchiectasis are found as commensals in the upper respiratory tract microbiome. When these bacteria move into the lower respiratory tract, they can be treated as potentially pathogenic microorganisms (PPMs), since they use several strategies to prevent immune defense, such as inhibition of mucociliary clearance and changes in quorum sensing (QS) and, consequently, in the production of virulence factors, such as the formation of biofilms.19–21
Characteristics of the lower airway microbiota in bronchiectasis patients without CF correlate significantly with clinical markers of disease severity. 22 It happens because PPMs can be harmful to bronchiectasis patients. These microorganisms pose potential risk to lung infections and can secrete several inflammatory mediators that cause progressive tissue damage and airway obstruction. 23
Chronic bacterial infections play an important role in disease progression in bronchiectasis patients. Thus, bronchiectasis patients' quality of life may be related not only to their clinical condition but also to the level of infection and the microorganism. 24 As a result, identification of PPMs in patients' airways is of great importance, considering that chronic bronchial infections affect them at some point in the evolution of bronchiectasis. 25
Pathophysiology of this disease predisposes the persistence of colonizing microorganisms, which have often been associated with accelerated decline in lung function and a large number of exacerbations, thus worsening quality of life and leading to high mortality.26–28
Until recently, the lower respiratory tract was believed to be a sterile niche; however, recent studies have pointed out that it holds microorganism colonization. 15 There also appears to be a strong relationship between the host's microbiome and the emergence of chronic lung diseases. 29
In bronchiectasis patients, an increase in the proportion of potentially pathogenic species is observed as the severity of the disease increases, and, consequently, there is a reduction in the abundance and variety of bacteria, which compose the lower respiratory tract microbiome. Lung infections, induction of cytokine production, tissue damage, and progressive chronic obstruction impact the clinical course of the disease negatively.18,30 Several species of Gram-negative and -positive bacteria, mycobacteria, and fungi can directly harm the pulmonary system,31,32 mainly Pseudomonas aeruginosa, Haemophilus influenzae, Streptococcus pneumoniae, S. aureus, and Moraxella catarrhalis. 16 Non-tuberculous mycobacteria (NTM) can also be considered; even though the disease has been mainly reported in women, bronchiectasis has been diagnosed at old age and associated with the gastroesophageal reflux disease. 33
In addition, studies have shown that bronchiectasis patients who developed NTM in respiratory samples had milder disease and better lung function than patients who had P. aeruginosa. 34
The association of PPMs with persistence (chronicity) of the disease has been well documented in patients with chronic bronchial infection, mainly caused by P. aeruginosa and H. influenzae. 35
Other bacteria, such as S. aureus, have emerged in this condition and can cause accelerated clinical deterioration; however, the real impact of these infections has yet to be fully determined. 29
Experts agree that P. aeruginosa infection must be eradicated, especially in cases of CF, since they have well-characterized protocols, unlike cases of non-fibrocystic bronchiectasis, in which there is more extensive and diffuse involvement of the lungs. It suggests that there may be progressive functional decline and/or more severe exacerbations when there is chronic infection caused by other pathogenic bacteria, such as Burkholderia cepacia, Achromobacter xylosoxidans, Stenotrophomonas maltophilia, and sensitive MRSA. 36
Staphylococcus aureus
S. aureus belongs to the Staphylococcaceae family, which currently comprises 54 species and 28 subspecies. 37 In smears of biological material stained by the Gram method, staphylococci are found as gram-positive cocci—approximately one micrometer in diameter—which can be isolated, in pairs, but in most cases aggregated, forming clustered structures. 38
S. aureus is a bacterium that is often found on healthy people's skin and in nasal passages. Therefore, it can maintain symbiotic behavior with its host without causing any disease. S. aureus is capable of producing infections in various tissues of the human body and is associated with high rates of morbidity and mortality. It is one of the main causes of infections acquired both in the community and in the hospital environment. 39
It is the major cause of respiratory, bone, joint, and endovascular diseases. Most infections occur in people with multiple risk factors for infection,39,40 such as bronchiectasis patients. 41
Although it is not the most common microorganism that has been isolated, S. aureus has already been identified in non-fibrocystic bronchiectasis patients, with variation in frequency reports.42–44 In addition, it should be highlighted that its detection may not be entirely correct, as occasionally there may be cases of undiagnosed CF and allergic bronchopulmonary aspergillosis, 45 which reveal shortage of epidemiological data on S. aureus infection in non-fibrocystic bronchiectasis. 44
In cases of CF, S. aureus is the initial pathogen in the colonization of most patients' respiratory tracts. It is evident in the fact that, in the first clinical investigation of CF in 1938, when average survival was 2 years, all patients who survived beyond a week ended up dying from pulmonary complications in which S. aureus was frequently implied. 46 In fact, before 1950, S. aureus infection was the main cause of respiratory failure in a population that rarely reached 10 years of age. 47
The aforementioned reports show that the species in the respiratory tract has been known for a long time. In 1938, Dorothy Andersen associated the pulmonary clinical picture of bronchitis, bronchiectasis, pulmonary abscesses, and lobar pneumonia found in patients who had CF with S. aureus colonization. 48
S. aureus is usually the first microorganism detected in respiratory cultures, since it is established very early in patients. 49 Lung infection can be either chronic or intermittent. 50 Its impact on patient survival has been controversial. Some studies do not associate S. aureus with worsening clinical condition, 51 whereas others correlate S. aureus to a faster decline in lung function in cases of chronic colonization or colonization by beta-lactam resistant strains.51,52
S. aureus can persist for years and, in general, undergoes adaptations that will favor its maintenance in the pulmonary environment. The most common one is the formation of small-colony variants (SCV). 53 The development of the SCV phenotype provides a survival strategy for S. aureus, which remains within the host's cells, without provoking a strong response, and acts as a reservoir for persistent infection. 54
The SCVs induce a less intense host response, by comparison with the wild type of S. aureus after infection and invasion of eukaryotic cells. The SCV intracellular cells elicit a significant response, however attenuated by the host with a lower release of chemokines, such as CCL5 (RANTES), CXCL10 (IP-10), CXCL11 (ITAC), and molecules of intracellular adhesion ICAM-1 after endothelial cell infection, by comparison with the wild type. 55 Thus, S. aureus has several advantages in the SCV state in relation to the host's immune response. It represents an important avoidance strategy, due to low induction of chemokines and lack of HIF-1 factor activation, which seems to be essential for the bactericidal activity of phagocytes. 54
Persistence of SCVs is also related to low production of α-toxin. Reduction in selective production results in two populations of organisms, that is, an aggressive subpopulation (producing α toxin), which causes acute destruction of host tissues, and a subpopulation of latency that can persist in the protective environment of host cells and cause recurrent disease when the electron transport chain is reconstituted. 56
Most studies of SCVs focus on their intracellular persistence, but the role of biofilms in chronic infections should not be overlooked, especially in the context of pulmonary infections in CF patients. Mechanisms involved in the formation of biofilms by SCVs are not well understood, although some results suggest that adhesin, an intercellular polysaccharide, composes part of the biofilm matrix. 58
Adhesins and biofilm
Adhesion of pathogens to the host tissues is the initial and essential event in the pathogenesis of most infections and in the colonization and invasion of the host organism. It is considered a virulence factor. Thus, many pathogens, such as S. aureus, express specific surface proteins, called adhesins, that adhere to molecules of both host cells and extracellular matrix components, such as collagen, elastin, fibrinogen, and laminin, which are responsible for fixation of bacteria to tissues and for the interaction between host cell and tissue receptors.14,57
The S. aureus genome encodes more than twenty adhesins, 59 which are crucial for the development of microbial infections. They are expressed on cell membrane surfaces of microorganisms and are known as MSCRAMMs (acronym for microbial surface components recognizing adhesive matrix molecules). 57
S. aureus has numerous MSCRAMMs, such as homologous fibronectin binding proteins, FnbA and FnbB, that mediate adherence to host tissues. These proteins are encoded by two genes closely linked to both fnbA and fnbB genes. The latter has much of the sequence, which is homologous to the former. Both (FnbA and FnbB) are very important in the initial phase of adhesion in infectious processes by S. aureus and bind to fibronectin and elastin. FnbA can also bind to fibrinogen.14,60,61
Both clfA and clfB genes, which are not allelic variants, but different genes, encode ClfA and ClfB adhesins. The former is found in all growth stages of S. aureus, whereas the latter is found in the exponential growth phase of the bacterium under anaerobic conditions. ClfA and ClfB bind to fibrinogen and promote platelet aggregation,62,63 whereas ClfB also has the ability to bind to cytokeratin 1062,63 and cytokeratin 864 (Fig. 1). Many molecules that participate in adherence are also involved in different mechanisms of immune avoidance. ClfA binds to the regulatory factor of complement 1 (C1), increases the conversion of C3b to an inactivated form (iC3b), and impairs the phagocytosis of S. aureus. 65
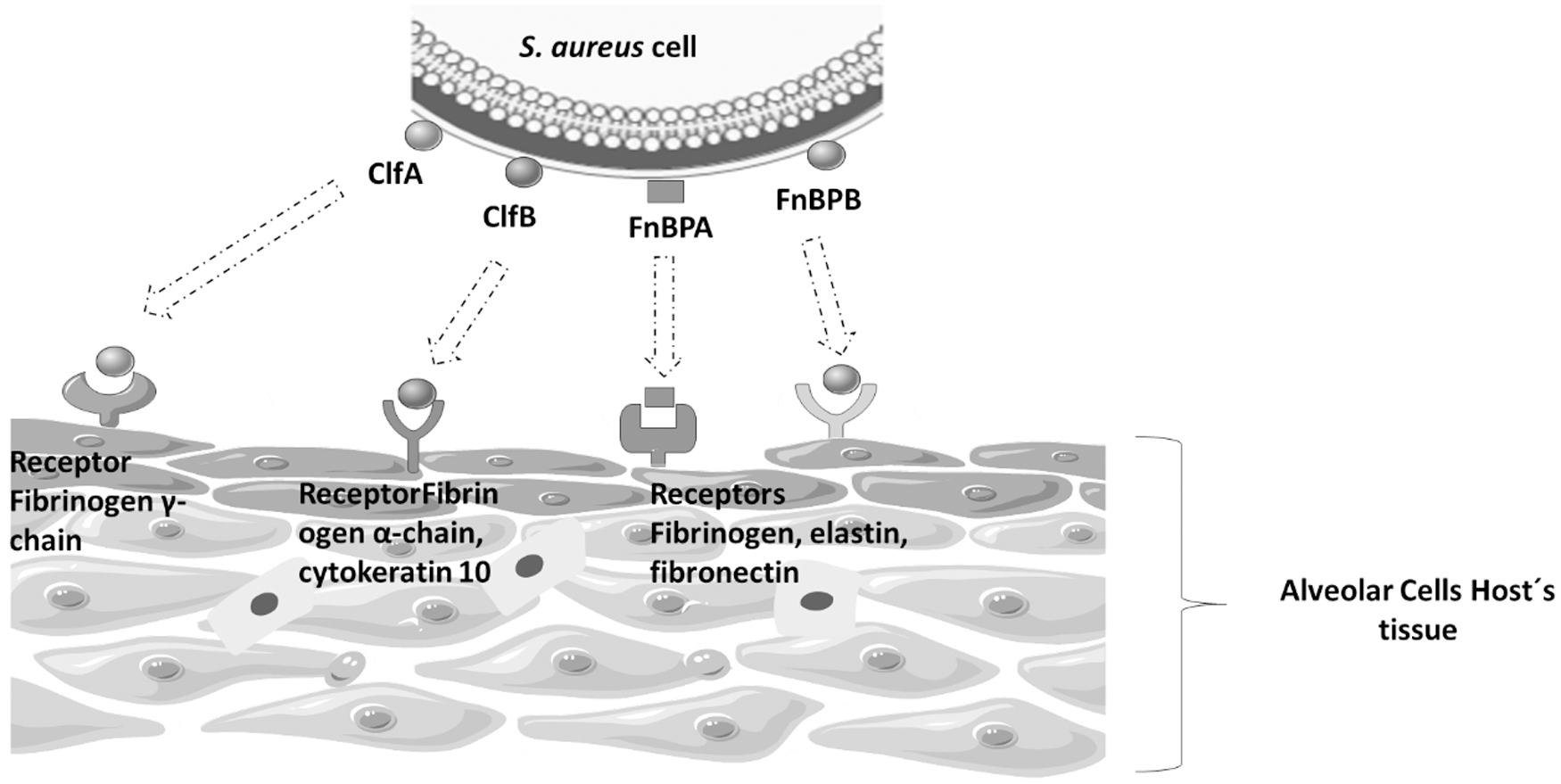
Staphylococcus aureus mechanisms of adherence to alveolar cells. S. aureus is well equipped to bind multiple host matrix proteins during infections. It may adhere to cells via interactions among the adhesins expressed on the bacterial cell surface as ClfA, CflB, FnBPA, and FnBPB, which are anchored in specific receptors located in the alveolar cells host's tissue.
After initial adhesion to the surface, intercellular adhesion and biofilm formation can occur. The biofilm consists of an aggregate of microorganisms surrounded by an extracellular matrix of polysaccharide or protein origin that they produce. Bacteria remain adhered to abiotic or biotic surfaces,66,67 forming “sessile” bacterial communities characterized by cells attached to the substrate and to each other, which are surrounded by a polymeric matrix called “slime.” Biofilm bacteria exhibit an altered phenotype with respect to the rate of growth and transcription of genes. 68
The development of bacterial biofilm has some peculiarities, depending on differences in metabolism of each microorganism, but, in general, it is divided into three main stages: adhesion, colonization, and maturation. 69 The S. aureus species was deeply studied regarding its biofilm formation, and five stages were observed: adhesion, multiplication, exodus, maturation, and dispersion. 72
Adhesion and multiplication
In the adhesion phase, the microorganism binds to receptors in the host through adhesins.70,71 In this phase, S. aureus cells begin to divide to accumulate and produce several extracellular proteins and promote intercellular binding that helps to prevent cells from detaching from the surface. During the multiplication phase, part of the bacterial population dies and lysis releases DNA and cytoplasmic proteins in the extracellular environment, involving the existing living cells. 72 In Staphylococcus, the most important component in the adhesion among bacterial cells is polysaccharide intercellular adhesin (PIA).
PIA synthesis is mediated by the icaADBC operon locus. 73 This operon is composed of the icaR gene, which is involved in the regulation of ica structural genes74–76 and four icaADBC genes responsible for the biosynthesis of this polysaccharide. 71 Together with other polymers, such as theoretical acids and proteins, they form the extracellular matrix called “slime.” 77
IcaA, IcaD, and IcaC proteins, located in the cytoplasmic membrane, participate in the extension of the polysaccharide chain. The IcaA protein represents the catalytic N-acetylglycosaminatransferase activity, forming the first oligosaccharides. Alone, it has low activity, which increases when coexpressed with IcaD, leading to the synthesis of larger β 1,6-N-acetylglycosamine residues, with 10 to 20 oligomers.66,75 However, long oligomers of up to 130 residues are formed only in the presence of IcaC. This protein may still act in the transmembrane transport of the formed polymer. The icaB gene encodes a protein that catalyzes deacetylation reactions during PIA biosynthesis.74–76
Deacetylated PIA seems to be essential for biofilm formation, immune evasion, and adhesion to epithelial cells. PIA deacetylation significantly affects its biophysical properties, which is crucial for the fixation of PIA on the bacterial cell surface and increases solubility in an aqueous environment, in addition to being indispensable for the formation of efficient biofilm in the process of colonization of abiotic surfaces and epithelial cells. Finally, PIA deacetylation plays an important role in immune evasion processes. 76
Exodus and maturation
In the next stage, called “exodus,” part of the cell population produces nuclease that degrades DNA within the biofilm matrix, detaching it from most accumulated population. A microcolony forms with the remaining cells and results in restructuring of the biofilm that seems to be an important stage for the formation of the secondary structure in the next stage. 72
A mature biofilm is characterized by microorganisms within the matrix surrounded by the “slime,” forming several layers. In this structure, there are microcolonies separated by channels where nutrients and oxygen reach cells and waste and toxic products of cellular metabolism are eliminated. 65
Regulation of biofilm formation
Biofilm formation in S. aureus is a process controlled by global virulence regulators, such as SigB, LuxS, SarA (SarA, staphylococcal accessory regulator), and agr QS (agr, accessory gene regulator). 78 The agr locus is a grouping of genes with QS activity, consisting of two divergent operons, controlled by P2 and P3 promoters, which regulate the expression of various virulence factors. The operon whose promoter is P2 encodes four proteins (AgrA, AgrB, AgrC, and AgrD), whereas the operon with P3 promoter encodes the effector molecule of the agr system (RNAIII). Products of the agrB and agrD genes, which are AgrB and AgrD proteins, respectively, form the self-inducing peptides (AIP). AgrC is a transmembrane protein with an AIP receptor. When critical bacterial cell density is reached, high concentrations of AIP bind to the AgrC receptor, resulting in its activation and subsequent phosphorylation of AgrA. 63
Phosphorylated AgrA induces P2 and P3 promoters, activating the transcription of previously described proteins and, particularly, of RNAIII. It acts as a transcription factor for several virulence genes and appears to have two functions: the negative regulation of genes that encode proteins associated with the colonization process and the positive regulation of genes that encode bacterial exoproteins associated with host cell damage. Negative and positive regulation of genes involved in the described processes promotes establishment and development of infections by S. aureus. 64
The role of bacterial biofilm in persistent infections
In general, the host's immune responses generated during biofilm infection are largely ineffective, which leads to chronic disease. It occurs through several mechanisms, which include direct death of leukocytes—macrophages (MФs), myeloid-derived suppressor cells, and neutrophils—and reprogramming of the immune response. Death can be caused by toxins, such as α-hemolysin and leukocidin AB, which are produced in large numbers during biofilm growth and can directly kill MФs, neutrophils, and other leukocytes to inhibit immune recognition and microbicidal activity. In reprogramming of the immune response, biofilm tilts the infiltrating macrophages of M1 (which induce a medium of pro-inflammatory cytokines, release antimicrobial products and phagocytosis bacteria) toward the M2 subtype (which induces anti-inflammatory response), avoiding effective phagocytosis and bacterial death. 83 Besides, biofilm bacteria are 10–1,000-fold more resistant to antibiotics than genetically identical planktonic ones and may be associated with their high tolerance to antibiotics. 17
Therefore, in general, infections related to biofilms act as aggravators of patients' initial clinical conditions and can further weaken them, making them susceptible to the development of recurrent infections, as is the case of bronchiectasis.
Accumulation of mucus in the airways of bronchiectasis patients predisposes them to the growth of microorganisms and formation of biofilms that keep them partially protected from the action of antibiotics and the immune system. Biofilm formation also contributes to the persistence of infections and the risk of acute exacerbation, because encasement of bacteria in a polymer-based matrix reduces efficacy of host defenses and antibiotic activity. Antibodies and macrophages poorly penetrate the biofilm structure, and pathogens incorporated into biofilms have low susceptibility.80,81
It should be highlighted that the prevalence of these phenotypic changes in a large number of S. aureus bronchiectasis clinical isolates has not been well-documented. There are a few studies that evaluated the capacity of biofilm production in clinical isolates of the respiratory tract, such as the one carried out by Cakir Aktas et al., who analyzed 88 S. aureus clinical isolate samples recovered from respiratory tract specimens; and 31 of them were from CF patients. Biofilm production was detected in 96.8% of CF isolates; in this group, 32.3% exhibited a strong positive phenotype whereas in 47.4% of non-CF isolates, a strong positive phenotype was not observed (p < 0.05). 82
As the aforementioned study showed, S. aureus isolated from the respiratory tract can produce biofilm; however, further research needs to be carried out to affirm that there is a relationship between chronic infection and production of biofilm by S. aureus. It occurs with the P. aeruginosa species, whose relationship between chronic infection and biofilm is well understood. Among the characteristics of adaptation, the mucoid phenotype (MUC) stands out. It results from the overproduction of a polysaccharide called alginate, a polymer of D-manuronic acid and L-glucuronic acid that is the biofilm of the species. Once infection with the MUC phenotype is established, eradication is practically impossible.48,49
Just as the production of P. aeruginosa biofilm can lead to an infection that is difficult to eliminate, the same can occur with biofilm produced by S. aureus. Although the composition of staphylococcal biofilm is different from that of P. aeruginosa biofilm, the function is the same. It was observed by Schwartbec et al., who compared the persistence of mucoid and non-mucoid S. aureus isolates from the airways of CF patients. Their competition experiments assessed fitness loss and survival under starvation conditions to determine survival advantages, whereas phagocytic assays analyzed protection against neutrophilic killing. 84
The authors reported that mucoid isolates can persist in the airways for long periods since they possess a survival advantage, by comparison with the non-mucoid phenotype, that is, protection against neutrophil killing under aerobic conditions that may be optimized under nutrient limited conditions.
Therefore, further studies should be conducted with the aim of assessing whether the production of biofilm by S. aureus interferes with the survival of patients with recurrent lung infections, as in the case of bronchiectasis.
Superantigens
The high infectious potential of S. aureus is restricted not only to its fast multiplication and dissemination in tissues, but also to the production of molecules with great pathogenic power, which include enzymes and toxins. Studies have suggested that S. aureus colonization/infection may be a marker of severe disease; they have reported that its presence in bronchiectasis is not associated with CF, in which there are some suggestions of a negative association with lung health.44,85 S. aureus produces several extracellular toxins, such as enterotoxins (EEs) and toxic shock syndrome toxins 1 (TSST-1).86,87 Staphylococcal EEs and TSST-1 belong to the family of SAgs originally identified in S. aureus. Staphylococcal toxins were first described in 1959 by Bergdoll et al., 86 and 23 serologically distinct EEs are currently known.87,88
Common characteristics of EEs are high stability and resistance to most proteolytic enzymes, such as pepsin or trypsin, a fact that allows them to maintain their activity in the gastrointestinal tract and their thermostability. 89 The TSST-1 has many properties in common with EEs, such as the ability to stimulate nonspecific proliferation of T cells and induce release of interleukin (IL)-1 and tumor necrosis factor alpha (TNF-α), and is recognized as the main cause of the toxic shock syndrome, characterized by fever, hypotension, and shock in humans.78,79,90
All these toxins have superantigen activity, which, unlike conventional antigens, does not need to be processed by antigen presenting cells (APCs). The classic way of activating T cells of the immune system is through the binding of T cell receptors (TCRs) to processed antigens and shown by the major histocompatibility complex (MHC) of APCs. If the T cell recognizes the antigen as non-self, tyrosine kinase is produced, which stimulates proliferation and differentiation of T cells. Response is highly regulated to limit harmful effects. 91
Activation of T cells mediated by EEs is different due to their characteristics and ability to activate T lymphocytes in the β domain of the variable chain (Vß) of the TCR. Thus, a large number of T cells can be stimulated under exposure to EE. 84 SAgs bind to the lateral surfaces of both TCRs and MHC class II molecules, outside the antigen-binding site, distorting the normal interaction of TCR-MHC-II.91,92
This interaction results in mass activation and proliferation of nonspecific T cells.89,93 It is estimated that picogram concentrations of these SAgs may be sufficient to nonspecifically stimulate up to 50% of all T cells in the human body, 94 thus, producing pro-inflammatory cytokines, such as IL-1, IL-2, gamma interferon, and TNF-α. Production of large amounts of cytokines causes intense inflammatory reaction and damages host tissues95,96 (Fig. 2).
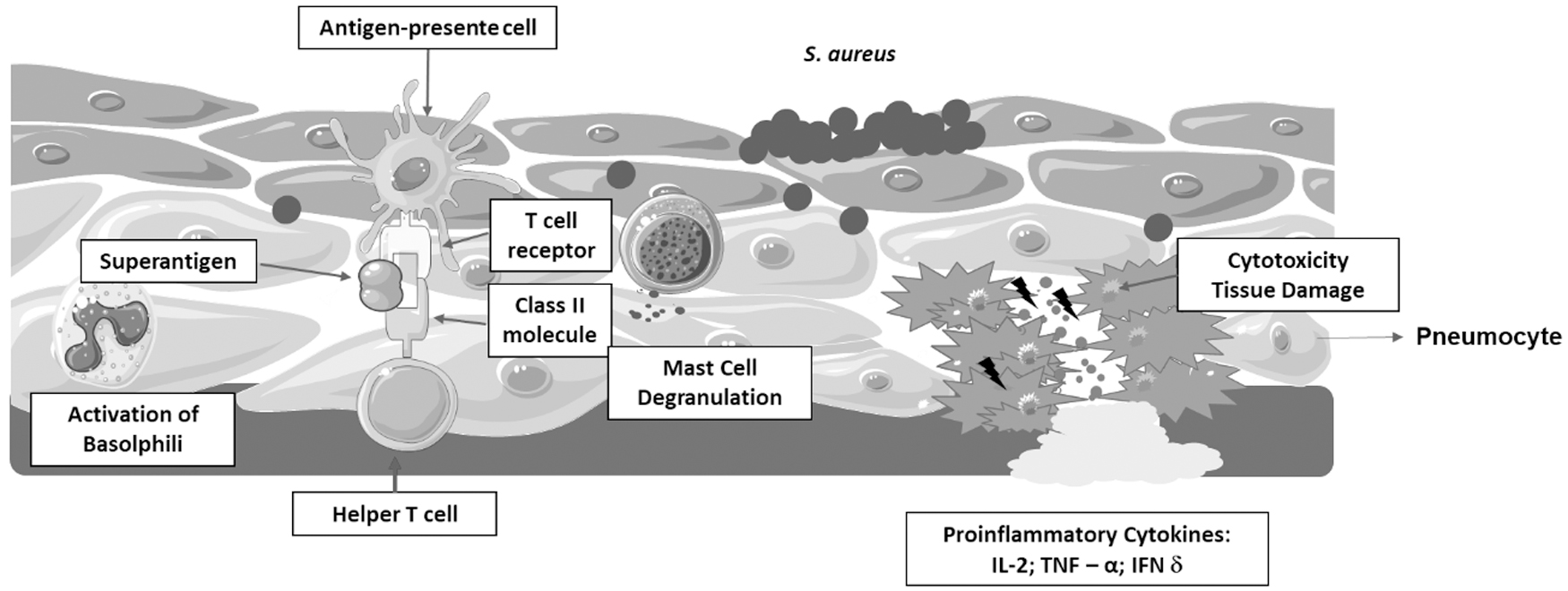
SAgs host receptor interactions: SAgs are produced intracellularly by S. aureus and are released on infection as extracellular mature toxins. SAgs are a class of antigens that result in excessive activation of the immune system. Specifically, it causes nonspecific activation of T cells, resulting in polyclonal T cell activation and massive cytokine. Some of the major products are cytokines such as IL-2, TNF-α, and IFN-γ. This excessive uncoordinated release of cytokines damages tissue and the organ system. IFN-γ, gamma interferon; IL-2, interleukin 2; SAgs, superantigens; TNF-α, tumor necrosis factor alpha.
SAgs can cause staphylococcal food poisoning and toxic shock syndrome and contribute to staphylococcal infection. Studies suggest that staphylococcal EEs have a broad role in the manifestation of several other human illnesses, including those associated with the respiratory tract.97–99
In rabbits, systemic enterotoxin type B (EEB) administration produces pulmonary inflammation characterized by leukocyte infiltration, damage to the endothelium, and high vascular permeability. 100 In addition, pulmonary inflammatory response characterized by an influx of lymphocytes, eosinophils, and neutrophils was also observed after intranasal EEB administration in mice. 101 DeSouza et al. showed that intratracheal administration of EEA in rats induces pulmonary inflammatory response characterized by predominant influx of neutrophils, whose maximum response is observed 4 hr after EEA administration. 102
Although the in vivo mechanism of action of EEs has not yet been clearly elucidated, detection of pro-inflammatory cytokines, such as IL-1β, IL-8, and the tumor necrosis alpha factor, and interactions between chemokine and cytokines in bronchial cells lead to the hypothesis that such interactions may result in the recruitment and activation of certain inflammatory cells that modulate the inflammatory process. This process triggered by EES by the antigen–antibody interaction can mediate the lesion of the bronchial wall, leading to plasma exudation at inflammatory sites, and can help to perpetuate inflammatory response in bronchiectasis.
Panton-Valentine leukocidin
Panton-Valentine leukocidin (PVL) is an exotoxin whose importance has been highlighted due to increasing isolation of S. aureus samples that produce it. It consists of a heptameric protein complex formed by two main proteins, known as components S and F, encoded by contiguous genes lukS-PV and lukF-PV, located in a prophage, because the genes that encode it are inserted in bacteriophages that are capable of integrating the S. aureus chromosome.103–105
Depending on the concentration, PVL forms pores in the cell or mitochondrial membrane of neutrophils and macrophages. When it is found at low concentrations, this leukocidin binds and forms pores in the mitochondrial membrane of polymorphonucleated cells, causing apoptosis. However, at high concentrations, PVL forms pores in the cytoplasmic membrane of these cells, leading to cell lysis and, consequently, releasing pro-inflammatory molecules. 106
Although the role of this leukocidin in the pathogenesis is not yet fully understood, epidemiologically, PVL-producing strains have been linked to necrotic lesions and severe necrotizing pneumonia.104,107
In vitro data support the role of PVL as a virulence factor in necrotizing pneumonia through recruitment and lysis of polymorphonuclear leukocytes and the release of cytotoxic granules, causing inflammation and direct lung injury. However, the required concentration of PVL toxin is not seen consistently in vivo; thus, extreme virulence found in some strains of S. aureus cannot be attributed to this leukocidin. 108 Even so, some cases of children with necrotizing pneumonia caused by positive S. aureus PVL were reported in the United States, Europe, and Asia; in some cases, the infection led to death.107,109–113
PVL has been highlighted as an important virulence factor, since it is constantly found in S. aureus samples that carry the mecA gene that confers resistance to all drugs of the beta-lactam class; as a result, they are called MRSA. 114
Methicillin Resistance
Antimicrobials that belong to the beta-lactam class act by inhibiting bacterial enzymes that are responsible for catalyzing the transpeptidation reaction, which is needed to join peptideoglycan chains, constituents of the cell wall. These enzymes are called penicillin binding protein (PBP), because they are the target-binding site and have high affinity with penicillin, structural analogues of its substrate. Consequently, methicillin resistance confers cross-resistance to all beta-lactam compounds, except for the fifth generation cephalosporin, ceftaroline, and ceftobiprole. 115
Production of PBP is related to the acquisition of the mecA gene, part of a mobile genetic element called SCCmec. SCCmec, also known as genomic island or antimicrobial resistance island, integrates the bacterial chromosome in a specific sequence—bacterial chromosomal attachment site (attBSCC)—located close to the origin of replication of staphylococci, at the 3′ end of an open reading frame (ORF)—of unknown function, called orfX.116–118 This element is a well-adapted vehicle for genetic exchange among staphylococcal species and appears to play an important role in the survival of bacterial cells in “stressful” environments. 119
MRSA are the cause of community and hospital infections.120,121 MRSA used to be a pathogen found exclusively in hospitals or hospital-related infections, called HA-MRSA, but in the mid-1990s, community-based cases of MRSA called community-acquired (CA)-MRSA began to emerge. Healthy individuals, without any hospital history or risk factors for the development of MRSA infections, got diseases related to this pathogen. 122
HA-MRSA is usually associated with risk factors, such as previous use of antibiotics, presence of a central vascular catheter, surgical procedures, and prolonged hospitalization, whereas CA-MRSA occur in patients who were neither hospitalized in a 1-year period nor underwent medical procedures. 123
The clinical picture of infections by CA-MRSA, in about 90% of cases, consists of skin and soft tissue infections, but the pathogen is also responsible for severe and potentially fatal clinical pictures, such as pneumonia necrotizing. These conditions may have a fulminating nature, leading to death in the first 6 hr of disease evolution. 124
The most severe conditions in CA-MRSA infections may be related to the fact that CA-MRSA isolates have more virulence factors, whereas HA-MRSA have more resistance determinants, since the former need to overcome natural barriers and healthy hosts' immune systems, whereas the latter finds patients who are more susceptible to infection (such as the elderly and the immunocompromised), but suffers high pressure from drugs used in hospitals. 125
With the emergence of β-lactam-resistant strains and the expression of multidrug resistance, treating infections in patients affected by MRSA has become a public health challenge, since methicillin-resistant strains have been associated with bad prognosis of several diseases, as reported by Ren et al. They carried out a large observational study, which collected data from patients in North America and showed that only MRSA in respiratory tract cultures is associated with more severe airflow obstruction, by comparison with those patients who only had methicillin-sensitive S. aureus. This association was observed in both young and old patients. 126
Multiresistance of MRSA strains is worrisome, because antibiotic therapy is one of the most potent weapons in the treatment of bronchiectasis. Since most patients have bronchial colonization, the use of antibiotics stands out mainly in the treatment of exacerbations.127,128 In addition, a characteristic of the disease is the high susceptibility to recurrent pneumonia.
Therefore, bronchiectasis patients are almost “addicted” to antibiotics, as they may require their constant use in a rotating regimen, and MRSA have been associated with worsening patients' clinical outcomes, 43 since antibiotic therapy often ends up being ineffective against this multi-resistant pathogen.
Therefore, bacterium resistance to antibiotics in respiratory infections of bronchiectasis patients is of increasing concern and its monitoring is important to patients' treatments.
Final Considerations
Chronic bronchopulmonary infection, the major cause of lung damage, can lead to inflammation of the respiratory mucosa and progressive lung destruction. Quality of life of bronchiectasis patients may be related not only to the patient's clinical condition but also to the virulence and resistance of the microorganism that infects or colonizes the lower airways.
The fact that S. aureus has recently stood out as an agent of chronic infection is worrying, since the high infectious potential of S. aureus has been well described in the literature. It is restricted not only to its ease of multiplication and dissemination in tissues, but also to the production of molecules with great pathogenic power, such as enzymes, toxins, adhesins, and biofilm that can induce an exacerbated immune response in the host. Besides, it allows strong adhesion to cells and components of the extracellular matrix of host tissues and protects against the activity of immune system components and antimicrobial agents (Table 1).
Summary Point
CA-MRSA, community-acquired methicillin-resistant Staphylococcus aureus; EEs, enterotoxins; PIA, polysaccharide intercellular adhesin; PVL, Panton-Valentine leukocidin.
In addition to virulence factors, infections caused by this pathogen are a clinical challenge, since they have high capacity for resistance development due to greater adaptation under selective pressure of intense use of antimicrobials. It is worrying in the case of bronchiectasis patients as they are frequently submitted to antimicrobial therapy, since the disease causes recurrent infections that need to be treated with antibiotics.
In the case of bronchiectasis, mainly non-fibrocystic ones, there are no studies worldwide. There are a few that are not conclusive with regard to lung injuries caused by S. aureus colonization/infection. Therefore, studies are necessary and must be carried out to allow more representative and accurate estimate of the importance of toxins and staphylococcal biofilm as an aggravating factor of this disease. Given the scarcity of data on the relevance of S. aureus in non-fibrocystic bronchiectasis, investigating the epidemiology and results of S. aureus infection in bronchiectasis is valuable.
Research on virulence and resistance of S. aureus may guide health professionals so that they can conduct effective treatment and avoid recurrence and persistence of pulmonary infections caused by this microorganism in bronchiectasis patients.
Future Perspective
S. aureus is an important human pathogen that frequently causes lung infection, a fact that has been attributed to its several virulence factors. In bronchiectasis, virulence factors, such as adhesins, biofilms, and toxins, must account for the complex interplay between this microorganism and its effect on the clinical disease. Even though some studies suggest a negative association with pulmonary health, data are still scarce and further research is needed.
Further studies of the role of virulence factors of S aureus in bronchiectasis could lead the way to regulate inflammatory reactions of the epithelium and to develop effective strategies to treat S. aureus-induced airway diseases.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Council for Technological and Scientific Development (CNPq) and Coordination for the Improvement of Higher Education Personnel (CAPES).