
Abstract
Aims:
WQ-3810 has strong inhibitory activity against Salmonella and other fluoroquinolone-resistant pathogens. The unique potentiality of this is attributed to 6-amino-3,5-difluoropyridine-2-yl at R1 group. The aim of this study was to examine WQ-3810 and its derivatives WQ-3334 and WQ-4065 as the new drug candidate for wild-type Salmonella and that carrying QnrB19.
Materials and Methods:
The half maximal inhibitory concentrations (IC50s) of WQ-3810, WQ-3334 (Br atom in place of methyl group at R8), and WQ-4065 (6-ethylamino-3,5-difluoropyridine-2-yl in place of 6-amino-3,5-difluoropyridine-2-yl group at R1) in the presence or absence of QnrB19 were assessed by in vitro DNA supercoiling assay utilizing recombinant DNA gyrase and QnrB19.
Results:
IC50s of WQ-3810, WQ-3334, and WQ-4065 against Salmonella DNA gyrase were 0.031 ± 0.003, 0.068 ± 0.016, and 0.72 ± 0.39 μg/mL, respectively, while QnrB19 increased IC50s of WQ-3810, WQ-3334, and WQ-4065 to 0.44 ± 0.05, 0.92 ± 0.34, and 9.16 ± 2.21 μg/mL, respectively.
Conclusion:
WQ-3810 and WQ-3334 showed stronger inhibitory activity against Salmonella Typhimurium DNA gyrases than WQ-4065 even in the presence of QnrB19. The results suggest that 6-amino-3,5-difluoropyridine-2-yl group at R1 is playing an important role and WQ-3810 and WQ-3334 to be good candidates for Salmonella carrying QnrB19.
Introduction
Substantial evidence states that nontyphoidal Salmonella (NTS) causes about 153 million gastroenteritis and 57,000 deaths in humans worldwide. 1 Salmonella enterica subsp. enterica serovar Enteritidis and Typhimurium are the major serotypes that cause foodborne diseases. 2 Even though NTS infections are self-limiting, in severe cases and immune compromised patients, the use of antimicrobials is advised.2–4 In such cases, traditional first-line antimicrobials, for example, ampicillin, chloramphenicol, and trimethoprim/sulfamethoxazole, are used. Fluoroquinolones such as ciprofloxacin have also been drugs of choice for treating NTS infections in both humans and animals following resistance to first line drugs. 5
However, resistance to ciprofloxacin among many Salmonella serovars circulating in humans and animal foods was reported. For instance, NTS was reported to be resistant to ciprofloxacin with the high prevalence in patients from Taiwan (48.1%), Thailand (46.2%), Korea (36.5%), Singapore (24.5%), Philippines (14.9%), Hong Kong (7.1%), Sri Lanka (8.0%), China (5.5%), and the United States (2.7%).6–8 In addition, multidrug-resistant Salmonella Typhimurium with definitive phage type 104 (DT104) was detected to have been contaminating meat and meat products. 9 In Europe, Salmonella Enteritidis has been seen resistant to ciprofloxacin in 13.3% from clinical isolates and poultry products.10–12
In the awake of this, the World Health Organization declared fluoroquinolone-resistant Salmonella spp. as a high priority bacterial infection. 13 Hence, research and development of the effective antimicrobial agents to address the emerging microbial resistance to traditional antimicrobials are necessary. 13
The resistance mechanism of fluoroquinolones is often associated with chromosomal mutations conferring amino acid substitutions at the quinolone-resistant determining regions of DNA gyrase subunits (GyrA and GyrB) and the DNA topoisomerase IV subunits (ParC and ParE) in Gram-negative and Gram-positive bacteria, respectively. In addition, quinolone resistance is also associated with plasmid-mediated quinolone resistance (PMQR) mechanism.14,15 PMQR has emerged in Enterobacteriaceae, rapidly transmitted by horizontal gene transfer thereby posing a significant threat to use of fluoroquinolones. 15 Plasmid-encoded quinolone resistance protein Qnr has been reported in PMQR and comprises of seven families, namely QnrA, QnrB, QnrC, QnrD, QnrS, QnrE, and QnrVC pentapeptide repeat proteins. Plasmid-encoded quinolone resistance protein Qnr binds to DNA gyrases and decreases susceptibility to quinolones. 16 Furthermore, efflux pumps (OqxAB, QepA, and QaqBIII) and Aac(6’)-Ib-cr (an aminoglycoside-, ciprofloxacin-, and norfloxacin-inactivating acetyl transferase) have also been reported to mediate quinolone resistance.15,16
Recently, WQ-3810, a novel fluoroquinolone showed strong inhibitory activity against several pathogens. Kazamori et al. 17 reported stronger inhibitory activity of WQ-3810 against fluoroquinolone-resistant clinical isolates of Escherichia coli and Acinetobacter baumannii than other fluoroquinolones (ciprofloxacin, levofloxacin, moxifloxacin, and gemifloxacin) by minimum inhibitory concentration (MIC) and IC50 measurement. Low toxicity of WQ-3810 was also mentioned in the same report. Koide et al. 18 reported higher inhibitory activity of WQ-3810 than ciprofloxacin and nalidixic acid against Salmonella DNA gyrase with amino acid substitutions in GyrA. Similarly, WQ-3810 was shown to have higher inhibitory activity than levofloxacin and ofloxacin against Mycobacterium tuberculosis and Mycobacterium leprae DNA gyrases with fluoroquinolone resistance conferring amino acid substitutions.19,20 These findings highlighted WQ-3810 as an effective novel antimicrobial agent against fluoroquinolone-resistant pathogens. The uniqueness is attributed to the R1 groups (6-amino-3,5-difluoropyridine-2-yl) and R7 groups (3-isopropylaminoazetizine-1-yl) enhancing the binding affinity to DNA gyrase.17,21 Recently synthesized fluoroquinolones, WQ-3334 and WQ-4065, with similar molecular structure as WQ-3810 at R1 (WQ-3334) and R8 (WQ-4065) positions would also be expected to have strong inhibitory effect on fluoroquinolone resistant bacteria.
In our previous study, the presence of QnrB19 was demonstrated to add quinolone resistance to wild-type Salmonella DNA gyrase. 22 However, there is no information on the inhibitory effect of novel fluoroquinolones with 6-amino-3,5-difluoropyridine-2-yl as R1 groups against QnrB19 protected Salmonella DNA gyrase supercoiling activity. Hence, the present study aimed to examine WQ-3810, WQ-3334, and WQ-4065 as new drug candidates for wild-type Salmonella and that carrying QnrB19.
Materials and Methods
Antibacterial agents
The antibacterial agents, WQ-3810, WQ-3334, and WQ-4065 (Fig. 1), were provided by Wakunaga Pharmaceutical Co., Ltd. (Osaka, Japan).
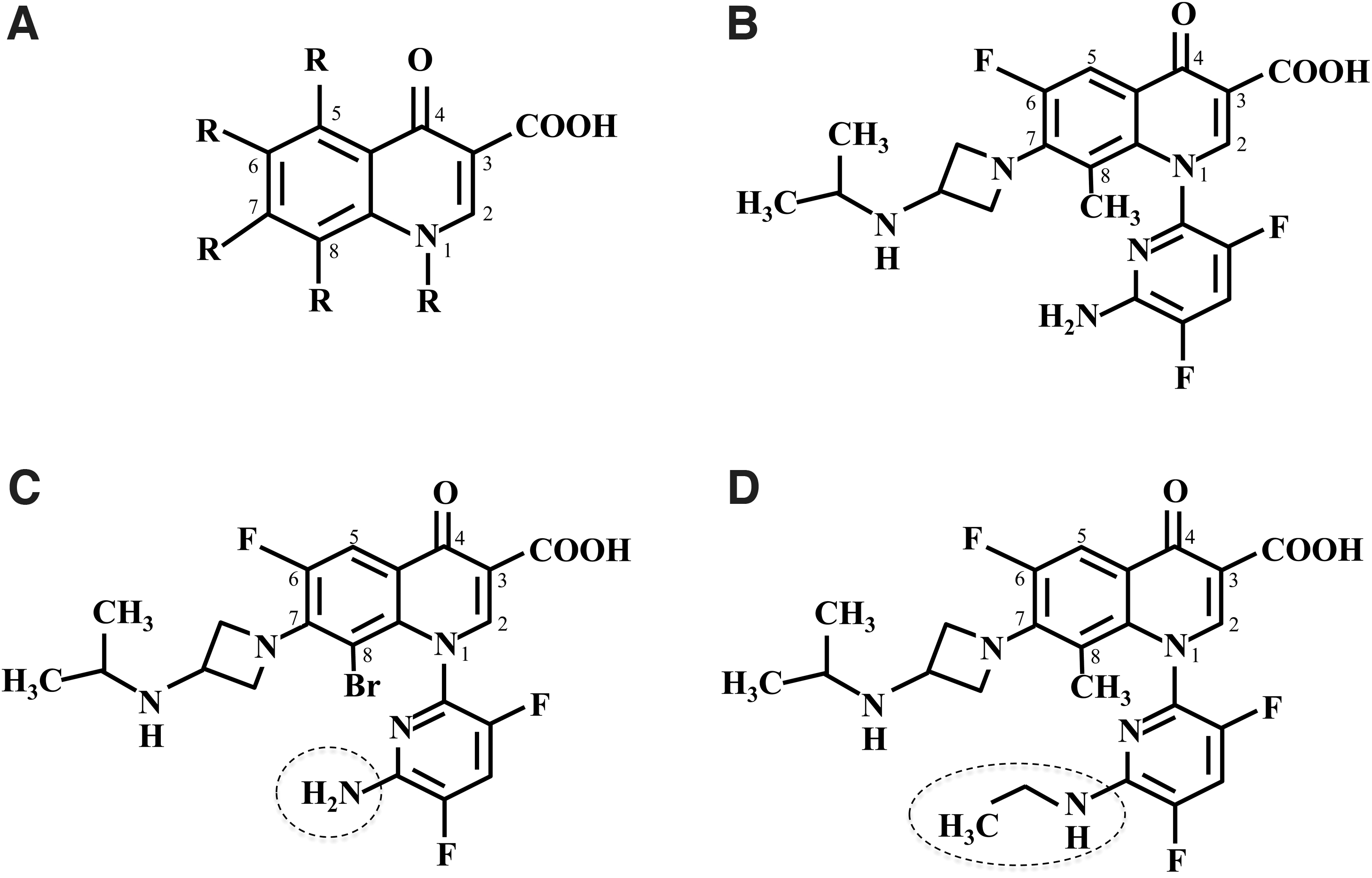
Chemical structures of fluoroquinolones used in the present study.
Expression and purification of Salmonella DNA gyrase subunits and QnrB19
Salmonella Typhimurium DNA gyrase subunits and QnrB19 used in the present work were prepared as in our previous work. 22 Briefly, Salmonella DNA gyrase subunits (GyrA and GyrB) were expressed and purified according to the procedures23–26 using Escherichia coli BL21 (DE3)/pLysS (Merck KGaA, Darmstadt, Germany) carrying gyrA or gyrB or qnrB19 in pET-28b(+) plasmid vector (Merck KGaA). The soluble proteins were loaded onto the column chromatography with Ni-NTA agarose resin (Thermo Fisher Scientific, Inc., Waltham, MA) to snare the His-tagged protein. The quality and quantity of all purified proteins were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) as shown in Fig. 2.

Purified recombinant DNA gyrase subunits of Salmonella and QnrB19 used in this study. Recombinant DNA gyrase of Salmonella and QnrB19 purities were examined by SDS-PAGE. Each protein sample (500 ng) was loaded on a SuperSep™ Ace 5–20% gradient gel. Lane M: protein size markers, lane 1: GyrA, lane 2: GyrB, lane 3: QnrB19. †Denotes GyrA with the MW of ∼97 kDa. ‡Denotes GyrB with the MW of ∼89 kDa. *Denotes recombinant QnrB19 with the MW of ∼25 kDa. MW, molecular weight; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis.
Inhibition of Salmonella DNA gyrase by quinolones and protection by QnrB19
The DNA supercoiling activity of Salmonella DNA gyrase was evaluated as previously described.22–26 Briefly, reaction mixture (30 μL) contained DNA gyrase assay buffer (35 mM Tris-HCl pH 7.5; 24 mM KCl; 4 mM MgCl2; 2 mM dithiothreitol; 1.8 mM spermidine; 6.5% glycerol; 0.1 mg/mL of bovine serum albumin), 1 mM adenosine triphosphate (ATP), 0.3 μg relaxed pBR322 DNA (Inspiralis Ltd., Norwich, UK), 18 nM DNA gyrase subunits (with/without fluoroquinolones), 0.16 μg/mL of WQ-3810, 0.64 μg/mL of WQ-3334, and 2.56 μg/mL of WQ-4065, and various concentrations of QnrB19 (6–4,374 nM; threefold serial increases). The reactions were incubated at 35°C for 60 minutes and stopped by adding 8 μL of 5 × dye mix (5% SDS, 25% glycerol, and 0.25 mg/mL of bromophenol blue). After incubation, the reaction mixtures were diluted 4 times with a 1 × DNA sample buffer (0.1% SDS; 0.05% Bromophenol Blue; 10% glycerol; and 100 mM Tris-HCl, pH 8.0) and loaded onto 1% agarose gel in Tris-borate-EDTA buffer for electrophoresis (96 minutes at 50 mA). The gel was stained with 0.5 μg/mL of GelRed (FUJIFILM Wako Pure Chemical Industries Ltd., Osaka, Japan). The amount of DNA supercoiling activity was measured by tracing the intensity of the bands with software ImageJ (http://rsbweb.nih.gov/ij). To confirm reproducibility, DNA supercoiling activity assay was carried out thrice.
Impact of QnrB19 on IC50 of fluoroquinolones
To examine the ability of QnrB19 to provide fluoroquinolone resistance to DNA gyrase, the DNA supercoiling activity of Salmonella DNA gyrase in the presence of QnrB19 and fluoroquinolones was investigated as previously described. 22 The reaction mixture was prepared in a final volume of 30 μL consisting of DNA gyrase assay buffer, 1 mM ATP, relaxed pBR322 DNA (0.3 μg) with 18 nM DNA gyrase subunit, fluoroquinolones (threefold serial concentration increases), 0.01–3.2 μg/mL of WQ-3810, 0.032–10 μg/mL of WQ-3334, and 0.32–100 μg/mL of WQ-4065, in the absence or presence of 18 nM QnrB19. Reactions, gel electrophoresis, and measuring DNA supercoiling activity were performed as explained in previous sections. Inhibitory effects of WQ-3810, WQ-3334, and WQ-4065 on DNA gyrase were assessed by determining the drug concentrations required to reduce half maximal inhibitory concentrations (IC50s), using the calculator at AAT Bioquest. To confirm reproducibility, the DNA supercoiling activity assay was carried out in triplicate.
Results
Inhibitory activities of WQ-3810, WQ-3334, and WQ-4065 against Salmonella DNA gyrase
Table 1 lists IC50s of WQ-3810, WQ-3334, and WQ-4065 as calculated by in vitro DNA gyrase assay with increasing concentration of each quinolone. Comparing to WQ-3810, WQ-3334 and WQ-4065 showed 2.2 and 23 times higher IC50s, respectively.
Impact of IC50s of Fluoroquinolones on WT Salmonella DNA Gyrase With/Without QnrB19
Position of substituents in the structures of fluoroquinolones.
IC50: Concentrations of fluoroquinolones to inhibit the supercoiling activity of DNA gyrase by 50%.
WT, wild type.
Impact of QnrB19 on Salmonella DNA gyrase supercoiling activity in the presence or absence of fluoroquinolones
Figure 3 shows the interaction among QnrB19, Salmonella DNA gyrase, and fluoroquinolones. At a concentration between 6 and 18 nM, QnrB19 showed little or less impact on DNA supercoiling activity of DNA gyrase. QnrB19 exhibited a concentration-dependent inhibitory activity against DNA gyrase at concentrations higher than 54 nM, while it completely inhibited the DNA gyrase activity at the highest concentration of 4,374 nM (Fig. 3A). WQ-3810 at the concentration of 0.16 μg/mL, WQ-3334 at 0.64 μg/mL, and WQ-4065 at 2.56 μg/mL reduced the DNA gyrase activity to 24.2, 11.1, and 19.3%, respectively (Fig. 3B–E), where QnrB19 at the concentration of 6–162 nM restored the DNA gyrase activity in dose-dependent manner to ∼4.45–4.28 times (WQ-3810), 5.78–6.80 times (WQ-3334), and 4.89–5.18 times (WQ-4065) (Fig. 3C).
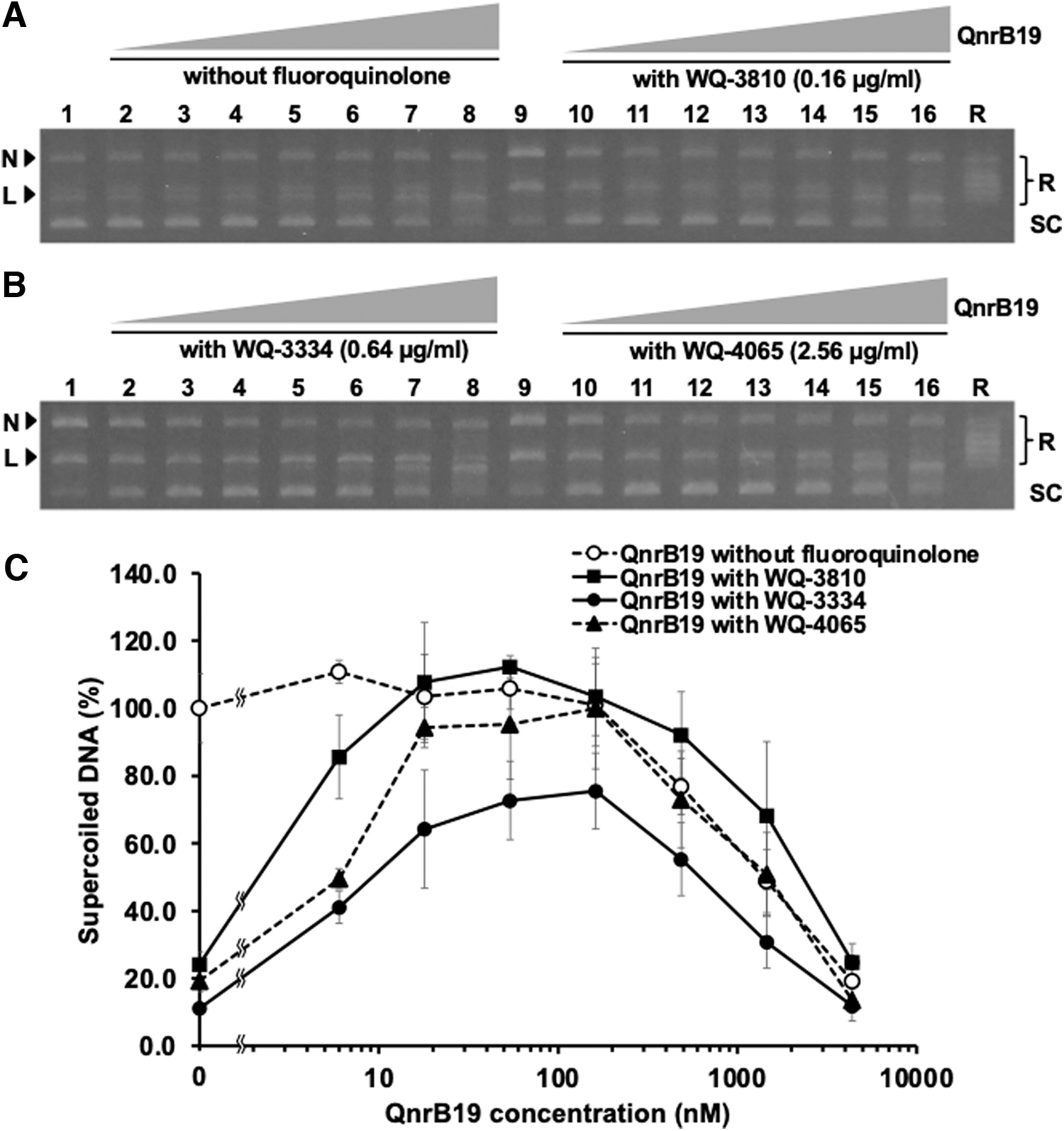
Inhibition of wild-type Salmonella DNA gyrase by fluoroquinolones and protection by QnrB19. Reaction mixtures were diluted 4 times with 1 × DNA sample buffer, and 10 μL of them were analyzed by electrophoresis in 1% agarose gels. Lanes 2–16: contained 0.3 μg relaxed plasmid pBR322, reaction buffer, and 18 nM DNA gyrase subunit.
Effect of QnrB19 on IC50s of fluoroquinolones
In a concentration-dependent manner, WQ-3810, WQ-3334, and WQ-4065 inhibited Salmonella DNA gyrase activity. Addition of 18 nM QnrB19, same concentration as DNA gyrase, diminished the inhibitory activities of all compounds examined in this study (WQ-3810; Fig. 4A, D, WQ-3334; Fig. 4B, E; and WQ-4065; Fig. 4C, F). Table 1 shows the effect of QnrB19 on IC50s of fluoroquinolones against Salmonella DNA gyrase. The IC50 of WQ-3810, WQ-3334, and WQ-4065 was increased by 14.2-, 13.5-, and 12.7-fold, respectively, with 18 nM QnrB19.

Effect of QnrB19 on IC50s of fluoroquinolones. Reaction mixtures were diluted 4 times with 1 × DNA sample buffer and 10 μL of them were analyzed by electrophoresis in 1% agarose gels. Lane 1–13: Reaction mixture contained 0.3 μg relaxed plasmid pBR322, reaction buffer, and 18 nM DNA gyrase subunit; Lanes 2–7: with 0.01–3.2 μg/mL WQ-3810
Discussion
Fluoroquinolones are drugs of choice for treating NTS infections; however, acquisition of resistance by Salmonella pathogens has limited the use of these drugs. WQ-3810 with unique substituents at R1 (6-amino-3,5-difluoropyridine-2-yl) and R7 (3-isopropylaminoazetizine-1-yl) has showed high activity against fluoroquinolone resistant pathogens. Two novel fluoroquinolones, WQ-3334 and WQ-4065, with similar structure to WQ-3810 have also been developed. However, there is limited information on the correlation between the structural features and inhibitory activities of these compounds against Salmonella DNA gyrase. In addition, there is no report on the impact of Qnr families against the activity of these compounds.
Park et al. 20 showed that interaction of R1 group in WQ-3810 with aspartic acid at position 464 in GyrB could enhance the inhibitory effect of this compound against DNA gyrase of M. leprae. The R1 group on WQ-3810 and WQ-3334 may contribute to high inhibitory activity of these quinolones (Table 1). A distinction at R8 between WQ-3810 [CH3] and WQ-3334 [Br] (Fig. 1) might explain the cause of 2.2-fold higher IC50 of the latter. In contrast, WQ-4065 with 6-ethylamino-3,5-difluoropyridine-2-yl at the R1 position showed 23.2-fold higher IC50 (Fig. 1). The addition of ethyl group at position 6 in R1 group strongly affected the DNA gyrase inhibitory activity. This explains the significance of the structure of R1 group in relation to the effectiveness of these novel fluoroquinolones.
At lower concentration (6–18 nM), QnrB19 had little effect on DNA supercoiling activity of both DNA gyrase (GyrA and GyrB) (Fig. 3C). However, increasing concentration of QnrB19 reduced the activity of DNA gyrase in concentration dependent manner evident by complete inhibition of DNA supercoiling activity at the highest concentration of 4,374 nM of QnrB19 (Fig. 3A). In this study, QnrB19 showed the ability to protect DNA gyrase from the effect of WQ-3810, WQ-3334, and WQ-4065 in a concentration dependent manner (Fig. 3). At the concentration of 18–162 nM, QnrB19 showed restoring ability on WQ-3810, WQ-3334, and WQ-4065 inhibiting DNA supercoiling activity of DNA gyrase by ∼4.3 to 4.5-, 5.8 to 6.8-, and 4.9 to 5.2-folds, respectively (Fig. 3C). This agreed with the results of our previous work, where, at the concentration of 54 nM, QnrB19 showed the maximum restoring ability on the nalidixic acid (50 μg/mL), norfloxacin (1 μg/mL), and ciprofloxacin (0.5 μg/mL) inhibiting DNA supercoiling activity of DNA gyrase by ∼1.8, 3.6, and 7.1-fold, respectively. 15 Comparing the results in this study with our previous study, QnrB19 showed restoring ability on the novel fluoroquinolones (WQ-3810, WQ-3334, and WQ-4065) inhibiting DNA gyrase activities similar to norfloxacin and ciprofloxacin, but higher than nalidixic acid. Thus, it is worth noting that QnrB19 protected DNA gyrase activity from old and new quinolone with distinct properties.
Under the existence of QnrB19 (18 nM), the IC50s of WQ-3810, WQ-3334, and WQ-4065 increased by14.2-, 13.5-, and 12.7-folds, respectively. In our previous work, 22 the presence of same molar concentration of QnrB19 caused 3.0- and 2.7-fold increase of IC50s of ciprofloxacin and norfloxacin, respectively, while there was no effect on the IC50 of nalidixic acid. These results suggested the structural differences, especially at R7 group (3-isopropylaminoazetizine-1-yl in WQ-compounds, piperazine in ciprofloxacin and norfloxacin, and none in nalidixic acid), to be the cause of distinct increase of IC50s by the addition of QnrB19. Conversely, better choice of R7 group may contribute to develop quinolones strong for the effect of Qnrs. This will be elucidated in our future study.
Elsewhere, IC50s obtained by DNA supercoiling assay targeting wild-type Salmonella DNA gyrase and MICs obtained by conventional culture-based drug susceptibility tests have been compared. Ciprofloxacin showed IC50 of 0.25 ± 0.05 μg/mL and MIC of 0.0078 μg/mL (32-fold lower than IC50). In contrast, nalidixic acid exhibited the IC50 of 65.1 ± 19.08 μg/mL and MIC of 4 μg/mL (16-fold lower than IC50). 23 In our previous study, MIC of WQ-3810 was 2.4-fold lower than IC50. 18 These facts show that the correlation between IC50s and MICs varies between quinolones. The Clinical and Laboratory Standards Institute 2019 defined MIC breakpoint for Salmonella spp. resistance to ciprofloxacin, norfloxacin, and nalidixic acid to be ≥1, ≥16, and ≥32 μg/mL, respectively. 27 Predicted MICs of WQ-3810, WQ-3334, and WQ-4065 against wild type Salmonella and those under the existence of QnrB19 were 0.013, 0.030, 0.30, 0.18, 0.38, and 3.8 μg/mL, respectively. WQ-3810 and WQ-3334 would be good candidate drugs for the treatment of salmonellosis caused by either wild type Salmonella or that carrying QnrB19.
In conclusion, novel fluoroquinolones, WQ-3810, WQ-3334, and WQ-4065, with unique difluoropyridine derivatives at R1 groups were assessed in this study to observe the inhibitory activity against wild type Salmonella DNA gyrase and interaction with plasmid-encoded quinolone resistance protein QnrB19. Our results show that WQ-3810 and WQ-3334 with 6-amino-3,5-difluoropyridine-2-yl at the R1 group showed stronger inhibitory activity against Salmonella DNA gyrases with a lower IC50s than WQ-4065 which have a 6-ethylamino-3,5-difluoropyridine-2-yl group at the R1 position either in the absence or presence of QnrB19. The results suggested that WQ-3810 and WQ-3334 could be good candidate drugs for wild type Salmonella and Salmonella carrying QnrB19.
Submission Declaration
This work has not been published previously. All authors meet the ICMJE authorship criteria.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported, in part, by a grant from the Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan, for the Joint Research Program of the Hokkaido University Research Center for Zoonosis Control to Y.S. and, in part, by Japan Agency for Medical Research and Development (AMED) under Grant Number JP20jk0210005, JP20jm0110021, and JP20wm0125008 to Y.S.