
Abstract
Streptococcus agalactiae (Group B Streptococcus , GBS) is a major agent of perinatal infections. Biofilms have been associated with GBS colonization and disease, as well as with infection persistence and recurrence. Although GBS remains susceptible to beta-lactams, it is still unknown how sessile cells respond to these antibiotics. Here, we evaluated the effect of different concentrations of penicillin (3–48 mg/L) on in vitro biofilm formation by four GBS strains belonging to serotype Ia/clonal complexes23 that were recovered from the oropharynx or urine of pregnant women and were previously characterized as strong biofilm producers. All four GBS strains were fully susceptible to penicillin (minimum inhibitory concentration = 0.023 mg/L), but penicillin was not able to fully prevent biofilm formation by these GBS strains. Biofilms formed in the presence of penicillin had reduced biomasses and thickness, but they were still classified as strong. Penicillin significantly reduced the density of live cells, but higher penicillin concentrations did not lead to improved prevention of biofilm formation. Biofilms formed in the presence of penicillin had no channels or long cocci chains observed in penicillin-free biofilms. Overall, results highlight the concerning possible impacts of biofilm formation in penicillin-based treatment and preventive strategies of GBS infections, even when the bacterial strain involved is fully antibiotic-susceptible.
Introduction
S
Although the genitourinary and/or gastrointestinal tracts are considered the main reservoir of GBS, the oropharynx can act as an alternative reservoir for horizontal transmission from mothers to their infants.3,4 Nevertheless, studies focusing on GBS strains recovered from the human oropharynx are still scarce.
The main virulence factor of GBS is the polysaccharide capsule that allows the recognition of 10 different capsular types (Ia, Ib, II-IX). 6 The distribution of capsular types varies according to geographical area and clinical presentation, but, in general, serotype Ia is among the most frequently found in both carriage and disease. 7 The relevance of serotype Ia is further ratified by its inclusion in GBS vaccine proposals currently being developed and evaluated. 8 Moreover, GBS strains recovered from human sources usually belong to one of four major clonal complexes (CC) as established by multilocus sequence typing (MLST), including CC1, CC17, CC19, and CC23.9,10
CC23 is mainly represented by serotype Ia strains, and it is one of the versatile GBS lineages that lack host tropism or clinical specificity, being recovered from carriage and diseases in humans, as well as in other animals.11,12
Colonization of host tissues is a critical step in the pathogenesis of GBS disease. In this regard, biofilm production is considered a key virulence factor.7,13 Persistence at and infection of certain anatomic sites is dependent on bacterial adhesion, which can be favored by biofilm production. 14 Moreover, when grown in biofilms, GBS can show increased antibiotic tolerance.15–17 Beta-lactam antibiotics, such as penicillin, are the drug of choice for treating and preventing GBS disease, especially because GBS strains showing reduced susceptibility to penicillin in conventional antimicrobial susceptibility tests are still rarely reported.18,19
However, mature GBS biofilms can tolerate penicillin concentrations up to 4,000 times higher the minimum inhibitory concentration (MIC) of planktonic counterparts. 16 The initial stages of biofilm formation are the most critical for their successful establishment; thus, elimination or reduction of initial planktonic cell density can be useful to prevent biofilm formation. 20 Nevertheless, the potential of penicillin to prevent biofilm formation by GBS when added at early stages of biofilm development has not yet been evaluated.
Recently, we have shown that most GBS strains recovered from different clinical sources in Brazil are strong biofilm producers, particularly those recovered from oropharynx and urine. 21 Here, we assessed the impact of different penicillin concentrations in the prevention of in vitro biofilm formation by four GBS strains belonging to the versatile serotype Ia/CC23 lineage and recovered from oropharynx and urine specimens. These strains stood out in a previous study 21 among the strongest biofilm producer GBS strains in our collection.
Materials and Methods
Bacterial strains
Four GBS strains belonging to serotype Ia and to sequence types (ST) included in CC23 were analyzed in the study (Table 1). All of them were previously characterized as strong biofilm producers. 21 Isolates were obtained during previous carriage surveillance studies performed as approved by the ethics committees of the institutions involved or recovered as part of the standard patient care procedures, so specific ethical approval was not needed.
Characteristics of Group B Streptococcus Strains Included in the Study
ST, sequence type.
Determination of susceptibility to penicillin
Penicillin MIC of planktonic-growing cells was determined according to CLSI guidelines and breakpoints 22 by using antibiotic gradient strips (Liofilchem® MIC Test Strips).
Assessment of biofilm prevention by penicillin
Biofilm formation assays were carried out by using the microtiter plate method as previously described 21 with minor modifications. Briefly, GBS strains were grown overnight in Todd-Hewitt broth (Oxoid) supplemented with 1% glucose. Culture suspensions were then adjusted to the turbidity of a 0.5 McFarland standard and inoculated into wells of 96-well polystyrene microtiter plates (NEST Scientific). For each GBS strain, control (without penicillin) and five test wells containing five different penicillin concentrations were evaluated.
All wells were inoculated with a total of 200 μL. In control wells, 180 μL of bacterial inoculum at 0.5 McFarland plus 20 μL of distilled water were added. In test wells, 180 μL of bacterial inoculum at 0.5 McFarland plus 20 μL of penicillin (Sigma) solution were added. Test wells consisted of five different penicillin concentrations: 3, 6, 12, 24, and 48 mg/L. Plates were then incubated at 37°C for 20 hours in a 5% CO2 incubator (Series II Water Jacket CO2 Incubator; Thermo Scientific) without shaking. After incubation, unattached bacteria were removed by washing with distilled water. Adherent bacteria were stained for 5 minutes with 0.1% (w/v) crystal violet solution, and bound crystal violet was released from stained cells by using ethanol.
Biomass was quantified by measuring absorbance values at 570 nm (optical density, OD570) using a microplate reader (SpectraMax Plus 384 Absorbance MicroplateReader; Molecular Devices). Each strain was analyzed in triplicate in two different assays. According to previously established criteria, 21 GBS isolates presenting biomasses with OD570 values ≥0.5 can be classified as strong/moderate biofilm producers, whereas those presenting absorbance values below 0.5 can be classified as weak or non-biofilm producers.
Confocal laser scanning microscopy of biofilms
Biofilms formed by strain Sag 010 in the absence (control wells) and presence of 48 mg/L of penicillin (test wells) were also analyzed by confocal laser scanning microscopy (CLSM) after staining with C™ BacLight™ Bacterial Viability Kit (Invitrogen) according to the manufacturer's instructions. Biofilms were formed in CELLview® slides (Greiner), where control and test wells were analyzed in duplicate, and visualized by using a Leica TCS-SPE confocal microscope at the excitation fluorescence wavelength of 485 nm and emission at 530 nm (green) and 630 nm (red). Data were analyzed by using ImageJ 23 and Comstat2.24,25 Sag 010 was chosen for CLSM analysis as this was the only representative of ST23 in the study, which is the most frequently detected ST and the probable common ancestor within CC23.11,12
Statistical analyses
Statistical analyses were performed by using the one-way analysis of variance (ANOVA) followed by Tukey's multiple-comparisons test or the two-way ANOVA with Bonferroni posttests, depending on the type of data. Tests were carried out by using the software GraphPad Prism v5.0, and p-values of <0.05 were considered significant.
Results and Discussion
GBS strains were susceptible to penicillin and confirmed to be strong biofilm producers
All four GBS strains presented penicillin MIC of 0.023 mg/L in the antimicrobial susceptibility assay, showing that planktonic-growing GBS are fully susceptible to penicillin according to CLSI (2020) breakpoints, which indicate susceptibility for MIC ≤0.12 mg/L. Likewise, all four strains were confirmed to be strong biofilm producers by the microtiter plate method, as previously reported, 21 presenting mean OD570 values in the control wells (without penicillin) between 1.337 (for Sag 083) and 2.981 (for Sag 029) (Fig. 1).
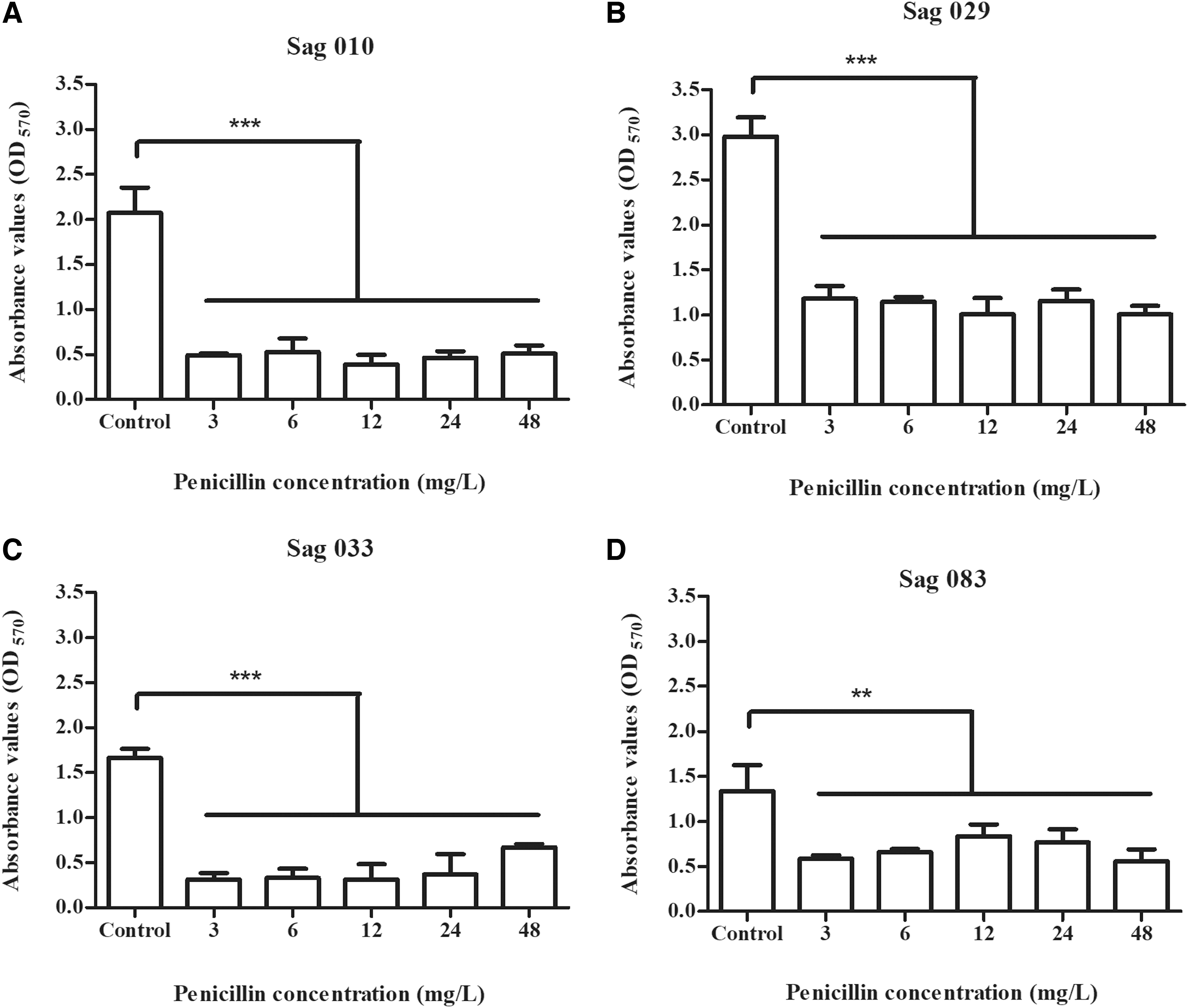
Mean absorbance values (as measured by the crystal-violet staining method at 570 nm) of biofilms formed in 96-well microtiter plates by four GBS isolates:
Penicillin reduced but did not fully prevent biofilm formation by GBS strains
Despite the full susceptibility in the antimicrobial susceptibility assay, penicillin was not able to fully prevent biofilm formation by the four GBS strains tested. Although biomasses were significantly reduced (p < 0.05) in all test wells (regardless of the penicillin concentration added) when compared with control wells (Fig. 1), mean OD570 values of test wells still represented nearly 20% (for Sag 010 and Sag 033), 30% (for Sag 029), or 40% (for Sag 083) of the absorbance values detected in control wells. These results indicate that biofilms were still formed by all four strains despite the addition of different penicillin concentrations.
Moreover, biomasses in test wells were, in general, represented by absorbance values higher than 0.5 (Fig. 1). For strains Sag 029 (Fig. 1B) and Sag 083 (Fig. 1D), mean OD570 values higher than 0.5 were observed in all test wells (at all penicillin concentrations tested). Thus, our results show that, in general, biofilms formed by GBS strains in the presence of different concentrations of penicillin are significantly reduced when compared with control but can still be classified as strong or moderate in most of the cases.
Higher penicillin concentrations did not lead to improved prevention of biofilm formation
In general, the lowest biomass OD570 values were detected at wells containing 12 mg/L of penicillin (Fig. 1), except for strain Sag 083 that had the lowest absorbance values at 48 mg/L (Fig. 1D). Nevertheless, there was no significant difference (p > 0.05) in biomass OD570 values among the different test wells for the four GBS strains included in the study (Fig. 1). This shows that increasing penicillin concentration did not necessarily lead to improved prevention of biofilm formation.
Of note, penicillin concentrations used in biofilm assays were nearly 100 (3 mg/L) to 2,000 (48 mg/L) times higher than the MIC obtained for planktonic-growing cells (0.023 mg/L). Moreover, the highest penicillin concentration adopted in the study (48 mg/L) corresponds approximately to the human serum concentration achieved after intravenous application of 5 million IU of penicillin G, 26 which is the initial penicillin dose given to pregnant women colonized by GBS as part of the intrapartum antibiotic prophylaxis of neonatal disease. 27 Overall, these results suggest that the ability to form biofilms may be associated with failure in penicillin-based treatment or prevention of GBS infections, even when bacterial strains seem to be fully susceptible to penicillin in the planktonic form.
Very few studies have analyzed antibiotic tolerance in GBS biofilms to date. Ruppen et al. 16 showed that penicillin concentrations of 2,000–4,000 times higher the MIC of planktonic-growing cells are required to eradicate a mature biofilm in vitro. They also observed that higher penicillin concentrations do not lead to more effective killing of sessile GBS in vitro, and they suggested that a prolonged penicillin treatment would be necessary to have an effect on biofilm-producing bacteria in vivo.
Penicillin changed the arrangement and reduced live cell density, biomass, and thickness of GBS biofilms
Although evaluating biofilm formation by the microtiter method using 96-well polystyrene plates presents many advantages, such as easiness of the procedure, quick turn-around time, and high reproducibility, it does not necessarily reflect a relationship between biomass absorbance value and bacterial cell viability. 28 In fact, OD values measured after crystal-violet staining include live cells, dead cells, and extracellular matrix. Thus, to assess the impact of adding penicillin in viability, biofilms formed by a representative strain (Sag 010) in control (without penicillin) and test (with 48 mg/L of penicillin) wells were also evaluated by CLSM analysis after staining with LIVE/DEAD™ BacLight Bacterial Viability Kit (Invitrogen).
The CLSM analysis of biofilms formed by Sag 010 in control wells (without penicillin) revealed the predominant presence of live (green) cells, which represented around 85% of the total biomass (Fig. 2A) and thickness (Fig. 2B) measurements. Moreover, CLSM images revealed the presence of long chains of cocci, as well as channels in the biofilm (Fig. 3A), which are known to be present in many bacterial biofilms to facilitate the circulation of water, nutrients, and oxygen throughout the community.29,30

Biomass

Biofilms formed by the GBS strain Sag 010 visualized by CLSM after staining with LIVE/DEAD BacLight Bacterial Viability Kit (Invitrogen) in control (without penicillin) wells
In turn, CLSM analysis of biofilms formed in test wells (with 48 mg/L of penicillin) showed that, in accordance with microtiter plate results, total biomass and thickness were significantly reduced (p < 0.05) compared with control wells (Fig. 2). Moreover, no chains of cocci or channels were observed (Fig. 3B), showing that penicillin also changed the arrangement of the structure.
However, live cells were still detected, despite a significant reduction (p < 0.05) in their density when compared with penicillin-free biofilms (Fig. 2). On the other hand, no difference (p > 0.05) was observed in dead cell density (Fig. 2) when compared with control, suggesting that penicillin had little or no effect in the last. In fact, certain antibiotics, including penicillin, are known to be effective only in actively growing cells 31 ; thus, little or no effect is expected in dead cells.
Further, spatial distribution of live and dead cells was different between biofilms formed in the presence and in the absence of penicillin. In biofilms formed in control wells (without penicillin), live (green) cells were characteristically concentrated at the upper parts of the structure whereas dead (red) cells were at the inner parts (Fig. 4A; Supplementary Video S1). In turn, biofilms formed in test wells (with 48 mg/L of penicillin) showed an inverse distribution of live and dead cell components. Live (green) cells were concentrated at the bottom, whereas dead (red) cells were concentrated at the top of the structure (Fig. 4B; Supplementary Video S2).
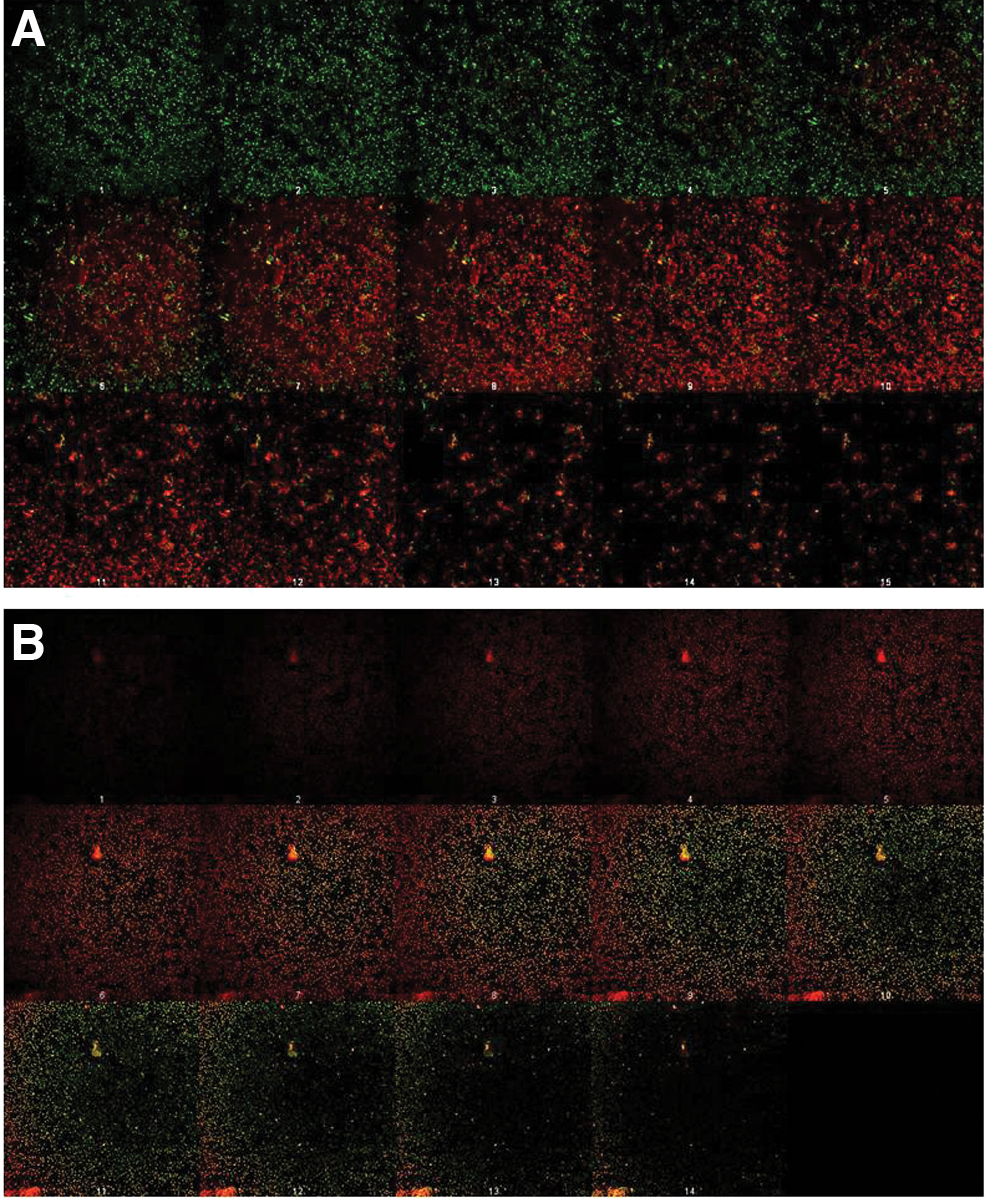
Biofilms formed by the GBS strain Sag 010 visualized by CLSM after staining with LIVE/DEAD BacLight Bacterial Viability Kit (Invitrogen) in control (without penicillin) wells
The distribution of live and dead cells within the biofilm usually reflects the availability of nutrients and oxygen throughout the community. 32 External layers of biofilms have easier access to nutrients and oxygen and usually become aerobic and metabolically active, whereas inner layers are generally anaerobic and nutrient-deficient leading to a reduced growth rate or even death of bacterial cells in this portion. However, this metabolic heterogeneity of biofilms is also known to contribute to the antibiotic tolerance of the whole community. Bacterial cells with slow or even interrupted metabolism, characterizing a state of dormancy, can represent up to 1% of live cells found within a biofilm.33–35
Multiple factors appear to contribute to antibiotic tolerance in biofilms, including decreased penetration through the extracellular matrix, overexpression of resistance mechanisms such as efflux pumps, and the presence of dormant or metabolically inactive cells.33–35 This state of dormancy is reversible and can be induced by antibiotic exposure.35–37 It has been reported that this phenotypic adaptation toward dormancy in Pseudomonas aeruginosa can start after only 1 hour of antibiotic exposure. 34 These dormant cells can more easily tolerate adverse conditions, and they are not readily killed by penicillin. Hence, they could survive exposure to penicillin and become metabolically active when conditions are favorable, 38 being then able to establish a later mature biofilm.
Our results suggest that the addition of penicillin in the initial process of biofilm formation first led to bacterial killing of most live cells, but simultaneously might have activated response mechanisms in the community that contributed to the switch of a subpopulation toward a dormant state. These live but dormant cells that might have survived exposure to penicillin can be represented among green-colored cells in biofilms formed in the presence of penicillin. Therefore, despite significantly reducing the density of metabolically active cells and changing biofilm architecture, penicillin, even in high concentrations, apparently failed to fully prevent biofilm formation.
Concluding Remarks
The GBS strains belonging to serotype Ia and CC23 represent a versatile lineage, with frequent occurrence in both carriage and invasive disease worldwide1,7,10,12,39 and, as such, are worth investigation. The four GBS strains evaluated in this study belong to this lineage, and besides ST23, considered the most common ST within CC23 and the ancestor of the group, they represent STs that have been detected across the world. According to PubMLST database (https://pubmlst.org/organisms/streptococcus-agalactiae; accessed on April 14th 2021), reports of ST88 have been made in Australia, Europe, the United States and Japan; whereas ST427 was detected in England.
Our results show that even in concentrations 2,000 times higher than the MIC of planktonic cells, penicillin was not able to completely prevent biofilm formation by GBS belonging to serotype Ia/CC23. Although penicillin significantly reduced the biomass, thickness, and density of live cells and changed biofilm architecture, live cells were still detected in penicillin-exposed biofilms. Such cells might have survived due to a reversible dormant state and might as well be the source for establishing a mature biofilm if and when conditions are favorable.
Limitations of the study include the small number of strains analyzed and the in vitro model chosen, and thus, further analyses are required to address important issues raised by the results presented here that could lead to the better understanding and controlling of biofilm-associated GBS infections. Nevertheless, overall results of the present study highlight the concerning possible impacts of biofilm formation in the penicillin-based treatment and preventive strategies of GBS infections, and they may help to explain, at least in part, certain reports 40 of treatment failure, recurrent infections, and intermittent colonization, even when appropriate antibiotics are administered and the bacterial strain involved is fully drug-susceptible.
Footnotes
Authors' Contributions
D.C.S.S.A. and L.C.S. performed determination of susceptibility to penicillin. D.C.S.S.A. and L.M.A.O. performed assessment of biofilm prevention by penicillin. D.C.S.S.A. and N.S.C. conducted CLSM of biofilms. D.C.S.S.A., L.C.S., N.S.C., and L.M.A.O. performed confirmation of bacterial identification and analysis of results. S.E.L.F., L.M.T., T.C.A.P., and R.B.R.F. supervised the experiments. D.C.S.S.A., L.C.S., N.C.S., L.M.A.O., L.M.T., and T.C.A.P. wrote the article. T.C.A.P. provided finance and infrastructure. All authors read and approved the final version.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported in part by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ); Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq); and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES)—Finance Code 001.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.