
Abstract
Background:
Amidst the era of widespread resistance, there has been a renewed interest in older antibiotics such as fosfomycin, owing to its activity against certain resistant Gram-negative pathogens, including multidrug-resistant variants expressing extended spectrum β-lactamases or carbapenemases. The goal of the study was to investigate pharmacokinetic/pharmacodynamic (PK/PD) index and PK/PD targets of fosfomycin in murine thigh and kidney infection models, employing clinical isolates of Escherichia coli (E. coli) and Klebsiella pneumoniae (K. pneumoniae).
Methods:
Seven isolates of E. coli (one wild-type and six clinical isolates) and five isolates of K. pneumoniae (one wild-type and four clinical isolates) were utilized for in vivo PK/PD studies. Single-dose plasma PK studies were conducted in infected mice by subcutaneous route. PD index was determined from exposure-response analysis employing 24-hr dose fractionation studies in neutropenic murine thigh infection model, while pharmacodynamic targets (PDTs) were derived from both thigh and kidney infection models.
Results:
Dose fractionation studies demonstrated that in vivo efficacy of fosfomycin best correlated with AUC/MIC for E. coli (R2 = 0.9227) and K. pneumoniae (R2 = 0.8693). The median AUC/MIC linked to 1 log10 kill effects were 346.2 and 745.2 in thigh infection model and 244.1 and 425.4 in kidney infection model for E. coli and K. pneumoniae, respectively. The mice plasma protein binding of fosfomycin was estimated to be 5.4%.
Conclusions:
The in vivo efficacy of fosfomycin against Enterobacterales was best described by AUC/MIC. The PDTs derived from this study may help define the coverage potential of fosfomycin at the clinical doses approved.
Introduction
The significant rise in resistance to even the last-resort antibiotics such as carbapenems among Gram-negative organisms coupled with limited availability or access to novel antibiotics compel clinicians to deploy older, underutilized antibiotics such as polymyxins, tetracyclines, aminoglycosides, and fosfomycin.1–5 The resistance scenario in certain parts of the world is such that the treatment of even urinary tract infections (UTI) caused by MDR pathogens is challenging to treat, often necessitating rescue therapies. 6 In such situations, polymyxins and fosfomycin could act as life-saving therapies, particularly for infections caused by metallo β-lactamases or dual carbapenemase producers for which newer treatment options are not effective. 7
Fosfomycin offers the advantage of availability of an oral dosage form, which is generally preferred in community settings such as UTI. Fosfomycin is available as a 3 g single oral dose (fosfomycin tromethamine and fosfomycin calcium) and was approved for the indication of uncomplicated UTIs many years ago. The parenteral formulation as fosfomycin disodium is available in some countries. 8 In light of nonavailability of MDR pathogens and active oral therapy options, as well as dearth of newer intravenous antibiotics, there is a need to undertake systematic assessments of pharmacokinetic/pharmacodynamic (PK/PD) features of fosfomycin with an objective of defining its therapeutic role against contemporary pathogens.
It is pertinent to note that, the doses and breakpoints of fosfomycin were derived years ago based on the knowledge of PK/PD available then. However, in recent years, significant progress in the area of PK/PD has been witnessed, which mandates re-assessment of fosfomycin from this point of view.
The objectives of our studies were to (A) evaluate the murine PK profile of fosfomycin in a neutropenic, thigh- and kidney-infected mice, (B) determine pharmacodynamic index (PDI) employing 24-hour dose fractionation studies, and (C) determine the magnitude of pharmacodynamic target (PDTs) of fosfomycin linked to stasis, and 1 log10 and 2 log10 kill, using neutropenic murine thigh and kidney infection models. The identified PDTs of fosfomycin may help in defining the coverage profile of fosfomycin at the approved doses. In addition, the integration of these PDTs along with available human PKs will also be useful in assessing the current susceptibility breakpoints of fosfomycin.
Materials and Methods
Antibiotic, bacterial isolates, and media
Fosfomycin disodium salt and Glucose 6-phosphate were purchased from Sigma–Aldrich. Aqueous solutions of fosfomycin were prepared freshly in sterile Milli Q water (Millipore) for administration in mice. Solutions were freshly prepared before each experiment, sterilized by passage through a 0.45 μm pore-size syringe filter, and kept at 2–8°C before use. Comparator antibiotics (Piperacillin-Tazobactam [PIP-TAZ], Meropenem [MEM], and Ceftazidime [CAZ]) were recovered from their respective commercial preparations and purity was ascertained by HPLC analyses conducted at Wockhardt Research Centre. Seven bacterial strains of Escherichia coli [six clinical isolates (harboring various β-lactamase phenotypes) and one wild type (not harboring β-lactamase phenotype)] and five K. pneumoniae (four clinical isolates and one wild type) strains were utilized in this study.
In vitro susceptibility testing
The minimum inhibitory concentrations (MICs) were determined in triplicate on three separate days using agar dilution in cation-adjusted Mueller-Hinton medium (HiMedia) supplemented with 25 mg/L glucose 6-phosphate as per CLSI guidelines. 9 MICs were also determined for comparator antibiotics such as PIP-TAZ, CAZ, and MEM by per CLSI-recommended agar dilution method. The MIC distributions along with resistance mechanism of these strains are presented in Table 1. The MIC of replicate assays is reported as mode value and utilized in PK/PD analyses. Strains harboring various resistance mechanisms are depicted in Table 1. Phenotypic resistance mechanisms were identified as per CLSI guidelines.10,11 The clinical isolates employed in current PD studies are part of Wockhardt strain repository and were collected during surveillance program conducted across tertiary Indian care hospitals. 10
In Vitro Activity of Fosfomycin and Comparator Antibiotics Against Escherichia coli and Klebsiella pneumoniae Strains Employed in This Study
FOS, fosfomycin; PIP-TAZ, piperacillin-tazobactam; MEM, meropenem; CAZ, ceftazidime; ESBL, extended spectrum β-lactamases; MBL, metallo β-lactamases; KPC, Klebsiella pneumoniae carbapenemase; MIC, minimum inhibitory concentration;
Susceptible, isolates not harboring beta-lactamase phenotype.
Murine thigh and kidney infection model
All PK and in vivo efficacy studies were carried out in 6-week-old outbred male Swiss albino mice, weighing about 25–30 g, procured from laboratory animal resources, Wockhardt Research Centre. Mice were kept in polyethylene cages with sawdust bedding maintaining 12-h day/12-h light cycle, room temperature 18°C–24°C, and relative humidity 40–70%. Standard rodent diet (Nutrivet life science, Pune) and drinking water were provided ad libitum. Animals were acclimatized for 3–4 days in laboratory condition before start of experiments. All experiments used in this study were approved by Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), an Institutional Animal Ethics Committee (Protocol approval Number: Form B/05/2020-21).
Neutropenia was rendered to mice following intraperitoneal administration of cyclophosphamide on 4 days (150 mg/kg) and 1 day (100 mg/kg) before thigh or kidney infection. 12 The overnight grown cultures were diluted (1:10) in cation-adjusted Muller Hinton Broth (CaMHB) and subjected for logarithmic phase (2 hr) before infection. The ODs of respective culture were adjusted at 580 nm using UV spectrophotometer (Shimadzu UV-1800). Following appropriate dilutions of initial inoculum (1–5 × 109) in fresh CaMHB, bacterial counts of the inoculum (1–5 × 106) were used for infection. On the day of infection, kidney infections with each of the strains were produced by administration of 50 μL of inoculum into each kidney 13 of isoflurane-anesthetized mice, while thigh infections were produced by injecting 100 μL of inoculum in right thigh of mice. 14 Two hours after induction of infection, treatment with fosfomycin was initiated.
Murine PKs of fosfomycin
Single-dose plasma PK studies of fosfomycin were conducted in kidney- and thigh-infected group of mice. The dosing solution of fosfomycin was freshly prepared in MilliQ water and administered as 10 mL/kg of dose volume. Fosfomycin was administered at 50, 100, 250, 500, 1,000, and 2,000 mg/kg by subcutaneous route at the base of a fold of loose skin in the posterior neck region of mice. Group of four mice per time point (divided in two sets; each set comprising of two mice, total: 200 mice for PK studies) were sampled for each dose at 0.25, 0.5, 1, 2, 4, 6, 8, and 10 hr postadministration of drug. Plasma was separated from blood and stored at −70°C until analysis.
Concentrations of fosfomycin in plasma samples were quantified by LC-MS/MS method. The calibration curve for fosfomycin was linear (R 2 > 0.96) over the range of 0.5–200 μg/mL with coefficient of variation not exceeding 5%. Lower limit of quantitation (LLOQ) was 0.50 μg/mL. The accuracy and precision (relative standard deviation, % RSD) were 102.3% and 4.3%. Bioanalytical data were acquired using Analyst v1.6.1 software (API4500; AB Sciex).
PK parameters (means ± standard deviations) like maximum plasma concentration (Cmax), area under the plasma concentration-time curve (AUC), clearance (Cl), and half-life (t1/2) were estimated by performing noncompartmental analysis using Phoenix WinNonlin 8.0 (Certara). The best-fit PK model (1 Compartment, extravascular with Lag time) developed from initial estimates of actual PK studies were used to simulate PK profiles of fosfomycin (Supplementary Table S3A). The predicted PK data were well in agreement with observed data. The simulated PK data were employed for all PK-PD analysis.
Determination of PK/PDI
The PK/PDI determination studies were conducted in neutropenic mice infected with E. coli ATCC 25922 and K. pneumoniae NCTC 13368 using thigh infection models. Total daily doses of fosfomycin between 12.5, 50, 200, 800, 1,600, and 3,200 mg/kg were administered subcutaneously fractionated into one (q24h), three (q8h), or six (q4h) equally divided doses to determine PK/PD indices (n = 3 mice/dose group, total: 120 mice for PDI studies). The mice were sacrificed 24 hr after initiation of drug treatment, and the thighs were removed aseptically and processed for colony-forming unit (CFU) determination. Vehicle control (MilliQ water)-treated mice (n = 3/strain) were sacrificed just before start of treatment and 24 hr after start of treatment.
The correlations between efficacy and the PK/PD indices (Cmax/MIC ratio, AUC0–24/MIC ratio and % T>MIC) were determined by nonlinear regression analysis using GraphPad Prism (GraphPad Prism; Version 7). The results of these studies were analyzed using the sigmoid Emax model derived from the Hill equation, E = E0 + (Emax × DN)/(EDN50 + DN), wherein, E is the effect (change in log10 CFU per thigh between treated and untreated mice after the 24 hr period of study; E0 is the baseline effect; Emax is the maximum effect; D is the total dose administered over 24 hr; ED50 is the dose required to achieve 50% of Emax; and N is the Hills slope of the dose-effect curve. 15
Estimation of PK/PDT for efficacy
In vivo PDT assessment studies were performed in murine thigh and kidney infection models for each isolate. Five to six doses of fosfomycin (twofold and fourfold increasing doses from 2.08 to 4,266.6 mg/kg in q4h regimen based on strain) were administered in q4h regimen to neutropenic thigh- and kidney-infected mice (n = 3 mice per dose group, total: 480 mice for PDT studies). The total daily doses of fosfomycin varying from 12.5 to 25,600 mg/kg/24 hr based on the MIC of strains were employed in the study. Untreated control animals were included for each strain. Drug treatment was initiated 2 hr postinfection. Animals were humanly sacrificed at 24 hr after start of treatment, and the thighs/kidneys were aseptically removed for cfu count determination.
The doses and PK/PD index target required to produce a net static effect (static dose), 1 log10 kill and a 2 log-kill, compared to the start of therapy were calculated wherever applicable for every isolate. The associated mean/median 24-h AUC/MIC targets were calculated for fosfomycin.
Results
Minimum inhibitory concentrations
MICs of fosfomycin and comparators (PIP-TAZ, CAZ, and MEM) against the strains used in the in vivo studies are presented in Table 1. Fosfomycin MICs ranged between 1 to 128 mg/L and 1 to 32 mg/L for E. coli and K. pneumoniae, respectively. The MICs of PIP-TAZ, MEM, and CAZ ranged between 0.25–256, 0.015–32, and 0.015–256, respectively, against these strains (Table 1).
Fosfomycin murine PKs
Subcutaneously administered fosfomycin demonstrated a linear and dose-proportional PK profile in healthy and thigh/kidney-infected mice (Fig. 1 and Supplementary Figs. S1, S2). The mean PK parameter estimates of fosfomycin are presented in Table 2. Across the range of doses evaluated, overall, plasma exposure of fosfomycin was comparable between healthy and infected (thigh or kidney) mice. The mean Cmax ranged from 117.7 to 999.4 μg/mL, while the mean exposure ranged from 84.4 to 807.1 μg.h/mL. The mean terminal elimination half-life was in the range of 0.6–0.7 hr. The % T>MIC was also calculated at all the evaluated doses and summarized for few selected doses in Supplementary Table S1 of Supplementary Data. The PK estimates for doses not evaluated in PK studies and employed in PD studies were simulated from best-fit PK parameters estimated from current PK study (Supplementary Table S3).

Mean plasma concentrations versus time profile (Mean ± SD) for fosfomycin following subcutaneous administration of single doses in neutropenic thigh- and kidney-infected mice.
Mean Plasma Pharmacokinetic Parameters (Mean ± SD) of Fosfomycin Following Subcutaneous Single-Dose Administration in Neutropenic Thigh/Kidney-Infected Mice
Determination of PK/PDI
PK/PD index of fosfomycin, which closely follows efficacy, was evaluated against E. coli ATCC 25922 and K. pneumoniae NCTC 13368 using a 24-hr dose fractionation study comprising doses equally divided over 24 hr, using q4h, q8h, and q24h dosing regimens. The relationships between AUC/MIC ratio, Cmax/MIC ratio, and % Time>MIC and PD responses for E. coli ATCC 25922 and K. pneumoniae NCTC 13368 are depicted in Fig. 2A–F, respectively. Nonlinear regression analysis using the sigmoidal Emax model demonstrated AUC/MIC as PK/PD index that best predicted the efficacy (R 2 = 0.9227 & R2 = 0.8693) followed by Cmax/MIC (R 2 = 0.7429 & R2 = 0.7607) and % T>MIC (R 2 = 0.4861 & R2 = 0.4293) for both the strains (Table 3).
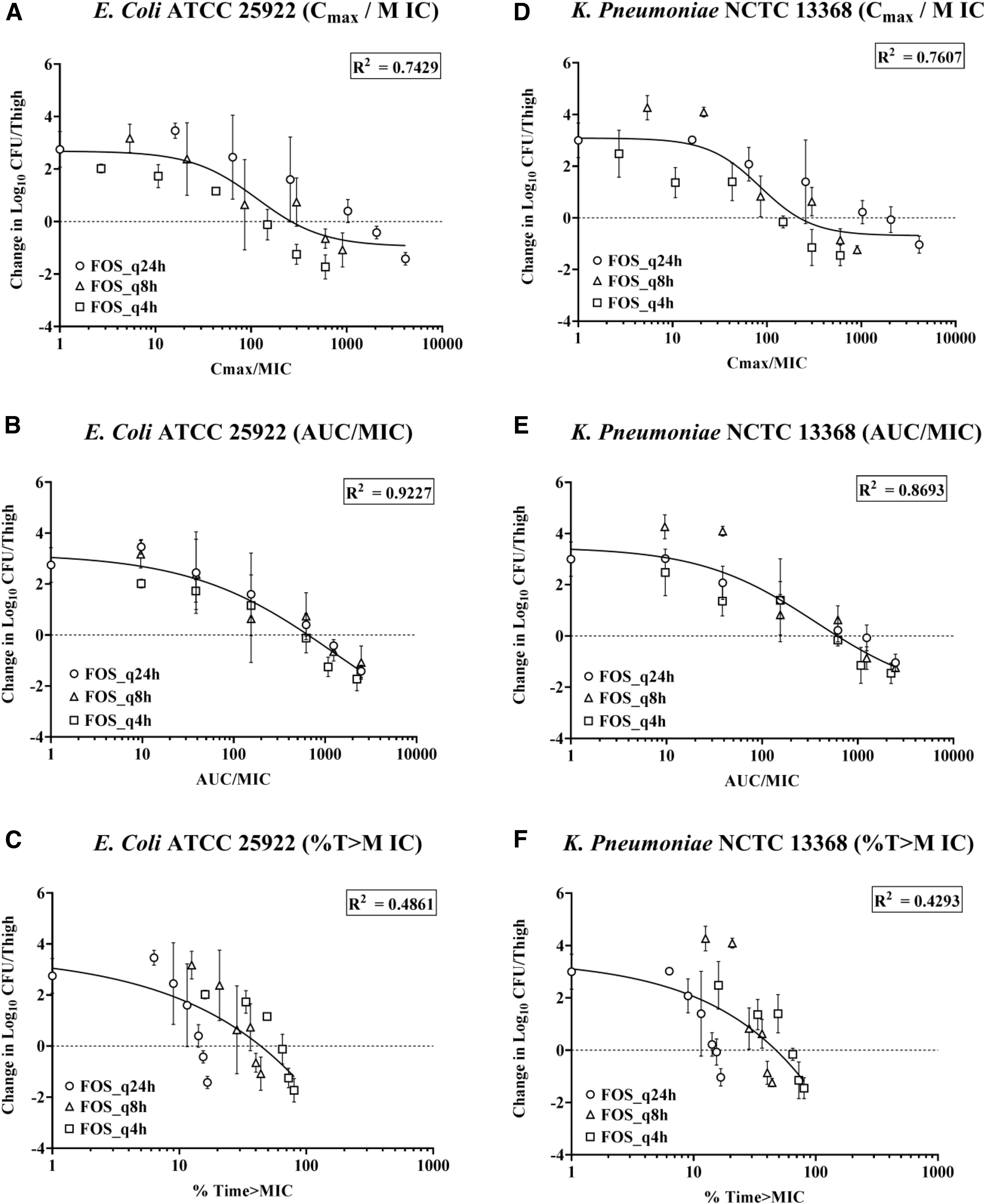
Relationships between PK/PD parameters and efficacy of fosfomycin against Escherichia coli ATCC 25922 (Left Panel)
Estimated Parameters of Sigmoid Emax Model for E. coli ATCC 25922 and K. pneumoniae NCTC 13368
E0, baseline effect; Emax, maximum effect; EC50, PD index associated with 50% of the Emax; R2, coefficient of regression;
Determination of PK/PDTs
Thigh infection model
The dose–response curves for all seven E. coli and five K. pneumoniae strains in thigh infection model are shown in Fig. 3 (Supplementary Figs. S3, S5A and Supplementary Tables S4, S6A). The thigh bacterial burden at the start of treatment (2 hr postinfection) ranged from 5.75 log10 to 7.27 log10 CFU/thigh, which increased at least 1 log10 CFU/thigh at the time of sacrifice (24 hr after start of treatment) in untreated group of mice.
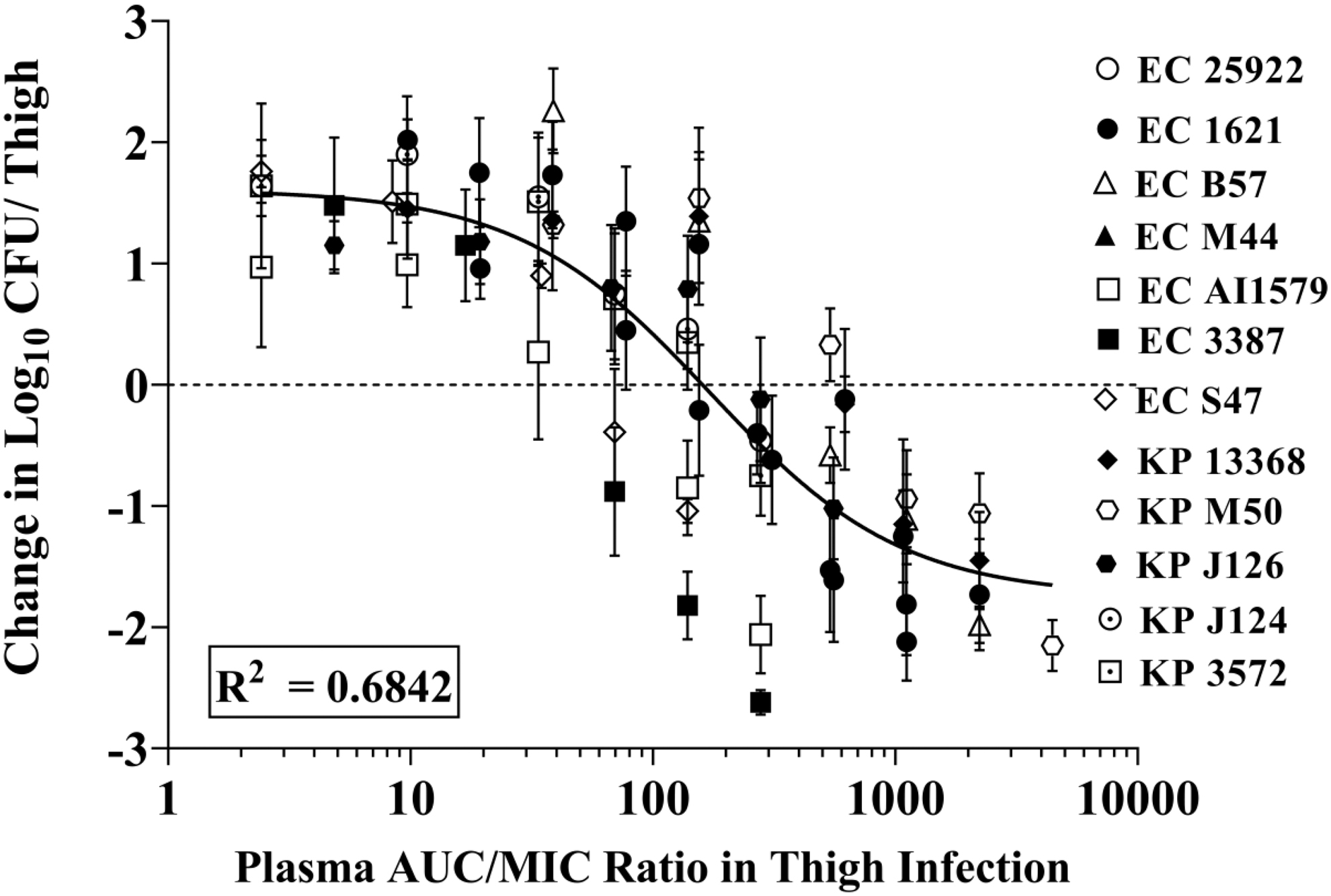
Co-modeled dose–response curves in relation to treatment effect against E. coli and K. pneumoniae strains using a neutropenic murine thigh infection model.
Maximal effect varied from near stasis to >2 log10 kill compared to the initial load at the time of treatment (2 hr). The plasma protein binding of fosfomycin was determined to be <10% in mice plasma (Supplementary Table S2); hence, protein binding was not considered for calculating AUC/MIC ratios. The median AUC/MIC values for stasis and 1 log10 kill effects were 171.5 and 346.2 for E. coli and 263.2 and 745.3 for K. pneumoniae, respectively. The co-modeled (E. coli + K. pneumoniae) 24-h AUC/MIC targets for stasis and 1 log10 kill were 159.9 and 541.6, respectively (Table 4). The 2 log kill PD effect was observed for most E. coli strains, except E. coli S 47 strain, while 2 log kill PD effect was not observed for Klebsiella strains.
Summary of Fosfomycin Pharmacodynamic Target-Associated (AUC/MIC) Requirement for Stasis, 1 log10 Kill, 2 log10 Kill and Calculated Stasis, and 1 log10 Kill Dose (mg/kg/day) Requirement in Murine Thigh Infection Model
NC, not calculated.
Kidney infection model
The dose–response curves for all seven E. coli and five K. pneumoniae strains are shown in Fig. 4 (Supplementary Figs. S4, S5B and Supplementary Tables S5, S6B). The mean kidney bacterial load at the start of treatment (2 hr postinfections) ranged from 5.75 log10 to 6.10 log10 CFU/kidney in untreated group of mice. At 24 hr after start of treatment, the increase in mean bacterial load in the kidney was >1 log10 CFU 24 hr in untreated groups.

Co-modeled dose–response curves in relation to treatment effect against E. coli and K. pneumoniae strains using a neutropenic murine kidney infection model.
The observed PD effect was in the range of 1 to 2 log kill with respect to bacterial load at the start of therapy across the strains. The median AUC/MIC value was for stasis and log10 kill effects was 132.0 and 244.1 for E. coli and 204.2 and 425.4 for K. pneumoniae, respectively. The co-modeled (E. coli + K. pneumoniae) 24-h AUC/MIC targets for stasis and 1 log kill were 102.4 and 380.4, respectively (Table 5). The 2 log kill PD effect was observed for all the strains, except K. pneumoniae J124 and 3572. Overall, PK/PD index-associated targets in kidney were lower compared with thigh-derived targets; this could be due to availability of higher fosfomycin concentrations in the well-perfused organ like kidney.
Summary of Fosfomycin Pharmacodynamic Target Associated (AUC/MIC) Requirement for Stasis, 1log10 kill, 2log10 kill and Calculated Stasis, and 1log10 Kill Dose (mg/kg/day) Requirement in Murine Kidney Infection Model
NC, not calculated.
Discussion
In light of an increase in resistance to currently available antibiotics, and the drought of new treatment options for MDR Gram-negative pathogens, a re-look at older antibacterial agents with the objective to maximizing their therapeutic scope is desirable. 16 In this context, significant efforts in re-assessing the PK/PD attributes of colistin have helped rationalize its dosing regimen to maximize efficacy and minimize toxicity. 17 Such analyses are based on integrating the potency of older agents against contemporary pathogens with systematic PK/PD analyses. Redefining the coverage potential of older antibiotics by assessing their PK/PD may help in rationalizing their clinical use. 18 Fosfomycin is an attractive candidate for such PK/PD analyses, as it is also available in the form of oral treatment option, a useful feature for the treatment of community infections.
Fosfomycin was introduced into the market decades ago when the science of PK/PD was relatively at an early stage, and with renewed interest in its usage, thorough investigations employing contemporary PK/PD information are required. 19 Currently, an intravenous formulation of fosfomycin is undergoing clinical development for the treatment of complicated UTI and pyelonephritis in the United States. 20 The existing study was designed to add to existing information on PK/PD of fosfomycin. Examination of studies aimed at identifying the PK/PD index of fosfomycin points toward discordance with some describing fosfomycin's driver of efficacy being time dependent, while others reporting it as a concentration-dependent drug.21–25
For instance, the bactericidal activity of fosfomycin identified by Parker et al mentions a time-dependent pattern, 22 while Perdigao-Neto et al were not able to conclude whether PD activity of fosfomycin is concentration or time dependent. 26 Roussos et al reported that differential time- or concentration-dependent PD activity of fosfomycin is organism specific. 27 The time dependent activity reported in these studies was determined by employing only one strain (Staphylococcus aureus) using static in vitro time-kill kinetic assay, likewise using same assay employing P. aeruginosa, ATCC 27583 Macleod et al reported time dependent activity of fosfomycin. 28 In contrast to it, concentration-dependent activity was reported by Mazzei et al using the same methodology employing E. coli and Proteus mirabilis strains. 29
These results suggest confusion among scientific community as to whether fosfomycin shows time- or concentration-dependent activity. Hence, re-assessment of robust in vivo activity as well as PK/PD properties of fosfomycin required to develop an effective dose and dosing regimen that complies with current standards is utmost important. This study was undertaken to further characterize PK/PD of fosfomycin against Enterobacterales employing nonclinical infection models. Generally, nonclinical infection models are used to identify efficacious dose and dosing regimens. The purpose of employing thigh as well as kidney infection model in this study was to validate the PDTs identified using plasma as surrogate marker and validate the identified efficacy driving parameters for UTI infections employing kidney infection model.
To increase the predictive robustness of PK/PD index magnitude, inclusion of diverse and multiple strains with wide variability is an important aspect of PK/PD studies. The existing study included seven E. coli strains and five K. pneumoniae strains with a wide range of MICs (1–128 μg/mL) linked to diverse resistance mechanisms. Overall, fosfomycin demonstrated dose-dependent in vivo bactericidal activity against all strains evaluated.
In a neutropenic mice thigh infection model, 24-h dose fractionation studies revealed AUC/MIC to be the PK/PD index most closely linked to efficacy followed by Cmax/MIC and least by T>MIC. The PDTs (24-h AUC/MIC) associated with 1 log kill ranged from 77.4–1,427.2 to 23.5–1,108.4 for Enterobacterales as determined in thigh and kidney infection models, respectively, while the median dose associated with 1 log kill was 3,882.2 and 2,801.9, respectively. In a previous study, PDT (24-h AUC/MIC) associated with 1 log kill was in the range of 12.8–78.0 for Enterobacterales using thigh infection model. 25
The PDTs determined using the thigh infection model in this study are higher, which could be attributed to robust PK data generated by collecting important early PK time points in mice, particularly within the 1st hour of dosing in our study. These key early PK time points were missing in Lepak et al's study along with overall lower fosfomycin exposures observed in mice PK studies, which lead to underestimation of PK parameters (Cmax, AUC and T>MICs) responsible for estimation of PDTs.
A previous study had shown that fosfomycin demonstrates concentration-independent activity, with minimal enhancement in antibacterial activity at concentrations above MICs, thus identifying T>MIC as the PK/PD index of fosfomycin. Most studies identifying fosfomycin PD index were based on in vitro PD studies employing time-kill kinetic assays, wherein application of dose fractionation studies generating variable Cmax, exposure, and time above certain concentrations, which differentiate interdependence of various PK/PD drivers, was not considered. Lepak et al using neutropenic thigh infection model and Hope and colleague employing in vitro dose fractionation studies using hollow fiber reported AUC/MIC as PD/PD index of fosfomycin.25,30
In addition, Zykov et al also conducted the PK/PD study of fosfomycin using mice UTI model employing multiple dosage regimens, 31 wherein plasma exposure reported for fosfomycin in mice were in line with exposure observed in this study, thus re-confirming PK profile and validating the PDTs determined in this study are robust and reliable. The PDTs identified using both the models are comparable, irrespective of the infection model suggesting plasma as surrogate maker can be reliably used for kidney infection models for identification of PDTs for drugs.
Based on the PDTs identified in this investigation and currently approved breakpoint of fosfomycin (64/32 mg/L), single oral dose of 3 g fosfomycin, which generates 24-h AUC of 165 μg.h/mL, 32 may not meet these PDTs. Even fosfomycin administered through intravenous route at 8 g single dose generating plasma exposure of 1,056 μg.h/mL 32 would not be able to achieve these PDTs. Higher PDT requirement of fosfomycin identified in our study is indirectly supported by one of recently published fosfomycin PD data, wherein higher doses (8 g every 8 hr, 1 hr, or 4 hr infusion) of intravenous fosfomycin have been recommended to achieve such higher PDTs. 33
Owing to highly resistant strains employed in this investigation, fosfomycin demonstrated moderate activities in in vivo studies, which suggest that therapeutic utility of fosfomycin could be limited to uncomplicated UTIs due to availability of high-enough concentrations of fosfomycin in urine to cover sensitive strains causing uUTIs.
However, coverage of fosfomycin could be expanded in complicated UTIs caused by highly resistant pathogens either by employing very high doses of fosfomycin, which would generate higher plasma, as well as urinary exposure of standalone fosfomycin or use of combination therapies of fosfomycin with other synergistic antibiotics. Hence, it shall be interesting to perform in vivo efficacy studies of fosfomycin in combination with other compatible antibiotics.
In conclusion, robust PK and PD data employing 12 Enterobacterales in thigh and kidney infection models revealed AUC/MIC as the PK/PD index that best correlates with bacterial killing for fosfomycin. The median AUC/MIC target values associated with 1 log10 kill was 378.7 and 333.3 using thigh and kidney infection model, respectively, for Enterobacterales.
Fosfomycin at current dose and dosing regimen may be appropriate for the treatment of uncomplicated UTIs due to availability of sufficient urinary exposure, but could limit its clinical utility against complicated UTIs caused by highly resistant strains, which would require higher plasma, tissue, as well as urinary concentrations of fosfomycin. However, therapeutic coverage of fosfomycin in cUTI caused by highly resistant strains could be improved by combination therapy of fosfomycin with other antibiotics, which needs to be confirmed by undertaking in vivo efficacy studies using combination drugs. The PD index and PDTs identified in this investigation could be useful or guide for defining the therapeutic scope of fosfomycin for the treatment of uncomplicated as well as complicated UTIs caused by MDR Gram-negative pathogens.
Footnotes
Acknowledgments
We thank the Wockhardt Research Centre, India, and Dr. Mahesh Patel for granting permission to conduct these studies. We also acknowledge the excellent technical assistance from Mr. Vineet Zope, Mr. Nilesh Chavan, Mr. Kushal Umarkar, Mr. Pavan Tayde, Mr. Prashant Joshi, Mr. Sukantakumar Naik, Dr. Kiran Patil, and Mr. Sanjay Shingare.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was self-funded.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.