
Abstract
Objective:
The objective of this study was to characterize ICEAplChn2, a novel SXT/R391-related integration and conjugation element (ICE) carrying 19 drug resistance genes, in a clinical isolate of Actinobacillus pleuropneumoniae from swine.
Methods:
Whole genome sequencing (WGS) of A. pleuropneumoniae CP063424 strain was completed using a combination of third-generation PacBio and second-generation Illumina. The putative ICE was predicted by the online tool ICEfinder. ICEAplChn2 was analyzed by PCR, conjugation experiments, and bioinformatics tools.
Results:
A. pleuropneumoniae CP063424 strain exhibited high minimum inhibitory concentrations of clindamycin (1,024 mg/L). The WGS data revealed that ICEAplChn2, with a length of 167,870 bp and encoding 151 genes, including multiple antibiotic resistance genes such as erm(42), VanE, LpxC, dfrA1, golS, aadA3, EreA, dfrA32, tetR(C), tet(C), sul2, aph(3)″-lb, aph(6)-l, floR, dfrA, ANT(3″)-IIa, catB11, and VanRE, was found to be related to the SXT/R391 family on the chromosome of A. pleuronipneumoniae CP063424. The circular intermediate of ICEAplChn2 was detected by PCR, but conjugation experiments showed that it was not self-transmissible.
Conclusions:
To our knowledge, ICEAplChn2 is the longest member with the most resistance genes in the SXT/R391 family. Meanwhile, ATP-binding cassette superfamily was found to be inserted in the ICEAplChn2 and possessed a new insertion region, which is the first description in the SXT/R391 family.
Introduction
Actinobacillus pleuropneumoniae is the causative agent of porcine contagious pleuropneumonia (PCP), a highly contagious respiratory disease causing significant economic losses to the swine industry in the world.1,2 To date, 19 serotypes of A. pleuropneumoniae have been identified on the basis of capsular antigen genes. 3 However, there were no effective vaccines developed to prevent and control PCP. Antibiotics are widely used to control this disease in the global swine industry, consequently, an increase in the resistance of A. pleuropneumoniae to the commonly used antibiotics has been reported worldwide in recent years.4–7
Integration and conjugation elements (ICEs) are not only an important mediator for horizontal gene transfer among bacteria, but also the most abundant conjugation elements found in prokaryotes, which can enable the host to obtain some new characteristics and improve its adaptability to the environment.8,9 The ICEs cannot independently replicate in bacteria, but it can be integrated into chromosomes of the hosts with integration and excision genes. After integration, ICEs can be excised from the chromosome and form circular molecules as intermediate products of ICE transferring.10,11
The SXT/R391 family is the most numerous and richest family of ICEs known to date. 12 Recent studies have shown that Proteus mirabilis is one of the most important hosts of SXT/R391 ICEs and SXT/R391 elements have also been identified from Pseudomonas mirabilis of human, gulls, and pigs.13,14 Recently, two types of SXT/R391 ICEs have been identified from A. pleuropneumoniae, including ICEApl2 and ICEAplChn1.15,16 Here we describe ICEAplChn2, a novel ICE related to SXT/R391 and carrying multiple antibiotics resistance genes, identified from A. pleuropneumoniae CP063424 strain.
Materials and Methods
Bacterial isolation and antimicrobial susceptibility testing
The pathogen was isolated from the lung of a pig farm in eastern China. The bacteria were inoculated in Tryptic Soy Agar (TSA) medium (containing 5% serum, 20 μg/mL nicotinamide adenine dinucleotide [NAD]). The isolates were identified and subjected to antimicrobial susceptibility testing. The antimicrobial susceptibility test was performed by microdilution of broth according to the previous study. 17 The tested strain was A. pleuropneumoniae CP063424, and the test drugs were clindamycin and florfenicol.
Identification and analysis of ICEAplChn2
Genomic DNA of A. pleuropneumoniae CP063424 was extracted using the TIANGEN Bacteria DNA Kit (TIANGEN, Beijing, China) according to the manufacturer's instructions. Whole genome sequencing (WGS) was completed using a combination of third-generation PacBio and second-generation Illumina. The putative ICE was predicted by the online tool ICEfinder. 18 Further sequence analysis of the ICEAplChn2 was done using BLASTn and BLASTx. Comparison analysis was generated by using Mauve, default parameters were used for all programs. 19
Phylogenetic analysis of ICEAplChn2
To explore the evolutionary origin of ICEAplChn2, we blasted its sequence in NCBI database and selected some high homology ICEs to construct the phylogenetic tree. In addition, the phylogenetic trees were constructed again for the core backbone genes of ICEs with high homology. Phylogenetic trees were performed with MEGA7 software by the maximum-likelihood method. 20 Bootstrap values were calculated with 1,000 replications.
Confirmation of the circular extrachromosomal form of the ICEAplChn2
ICEs existed in line or circular forms in the bacteria. To detect different forms of ICEAplChn2, four specific primers were designed and used for PCR amplification (Table 3).
Conjugal transfer of ICEAplChn2
Genomic DNA of A. pleuropneumoniae CP063424 was extracted using the TIANGEN Bacteria DNA Kit (TIANGEN) according to the manufacturer's instructions. The transformants were generated by electroporation using extracted genomic DNA according to a previous study. 21 A. pleuropneumoniae serotype 8 strain was cultured to an OD600 = 0.5 and washed three times with equal volume of precooled sucrose/glycerol (SG) buffer (15% glycerol, 272 mM sucrose), and finally resuspended using 80 μL SG buffer.
Eight microliters of DNA of A. pleuropneumoniae CP063424 was added into the competent cells and incubated in ice water for 30 min. After electroporation (2.5 kV), 1 mL of Tryptone Soy Broth (supplemented with 1/15 serum, 1/30 0.1% NAD, and 0.03 M sucrose) was added and incubated at 37℃ for 1 h, then the cells were plated into TSA supplemented with 256 μg/mL clindamycin to screen the positive transconjugants.
In this experiment, ICEHpsaHPS conjugal transferred to sHPS7 served as a positive control.
Results and Discussion
Antimicrobial susceptibility testing
The antimicrobial sensitivity test showed that A. pleuropneumoniae CP063424 was highly resistant to clindamycin with a minimum inhibitory concentration of 1,024 μg/mL, and was not resistant to florfenicol (Table 1). To find the reason for the high clindamycin resistance, we performed WGS and found that ICE contains the clindamycin resistance gene erm(42), which has been shown to be associated with clindamycin resistance in Pasteurella multocida from bovine respiratory disease. 22 Although A. pleuropneumoniae CP063424 was not resistant to florfenicol, we still found the flor gene in the ICE, which had the same results as ICEApl2. 15
Antimicrobial Susceptibility Testing of Actinobacillus pleuropneumoniae CP063424
MIC, minimum inhibitory concentration.
Identification and characterization ICEAplChn2
Through WGS, we found a novel ICE in the chromosome of A. pleuropneumoniae CP063424. The ICE integrates into the prfC gene through integrase, which are typical features of the SXT/R391 family. According to the relevant information, this novel ICE was named ICEAplChn2. The SXT/R391 ICE family is one of the largest ICE families, with the largest members among Gram-negative bacteria. 12 To date, at least 155 ICEs of the SXT/R391 family have been identified, but most of them are found in Vibrio cholerae, which is the etiological agent of the diarrhea disease cholera.23,24
In recent years, three ICEs have been identified from A. pleuropneumoniae, two of which belong to the SXT/R391 family and one belongs to the ICEHin1056 family. The results of Mauve sequence alignment showed that ICEAplChn2 is similar to the other two ICEs of SXT/R391 family identified from A. pleuropneumoniae, but many different variations were also observed (Fig. 1).

Mauve alignments of ICEAplChn2, ICEApl2, and ICEAplChn1. ICE, integration and conjugation element.
The chromosome of A. pleuropneumoniae CP063424 is 2,379,826 bp long with a guanine and cytosine (GC) content of 41.64%. ICEAplChn2, a 167,870 bp ICE, contains 151 open reading frames with 46.57% GC content, which inserted into the A. pleuropneumoniae CP063424 prfC gene, from 9,537 to 177,406 bp (Table 2). Blastn analysis of the complete sequence of ICEAplChn2 showed high homology with some SXT/R391 ICEs, including ICEs from Pr. mirabilis, V. Cholerae, and A. pleuropneumoniae.
Genomic Information of Actinobacillus pleuropneumoniae CP063424 and ICEAplChn2
GC, guanine and cytosine; ICE, integration and conjugation element; ORF, open reading frames.
Based on phylogenetic analysis, we found that ICEAplChn2 was most closely related to ICEchCHAHV1003 and ICEVchCHN2255, both of which were identified from V. cholerae. However, compared with ICEApl2 and ICEAplChn1 identified from A. pleuropneumoniae, ICEAplChn2 was located in different clusters, which may indicate that ICEAplChn2 and the other two ICEs are undergoing different evolution separately (Fig. 2).

Phylogenetic tree of ICEAplChn2 and 12 other SXT/R391 ICEs. The tree was constructed by the maximum-likelihood method with MEGA7. Bootstrap values were calculated with 1,000 replications.
As genetic map showed that attL and attR (accgcttatttttttg) were found at the end of element (Fig. 3). We also identified 18 multidrug resistance genes, including VanE, LpxC, dfrA1, golS, aadA3, EreA, dfrA32, tetR(C), tet(C), sul2, aph(3)″-lb, aph(6)-l, floR, dfrA, ANT(3″)-IIa, catB11, erm(42), and VanRE. Most of them are located in VRIII and HS4 (Fig. 4). However, vanE, LpxC, and IsaC genes are located at a novel insertion site after prfC gene, which has not been reported in the SXT/R391 family.
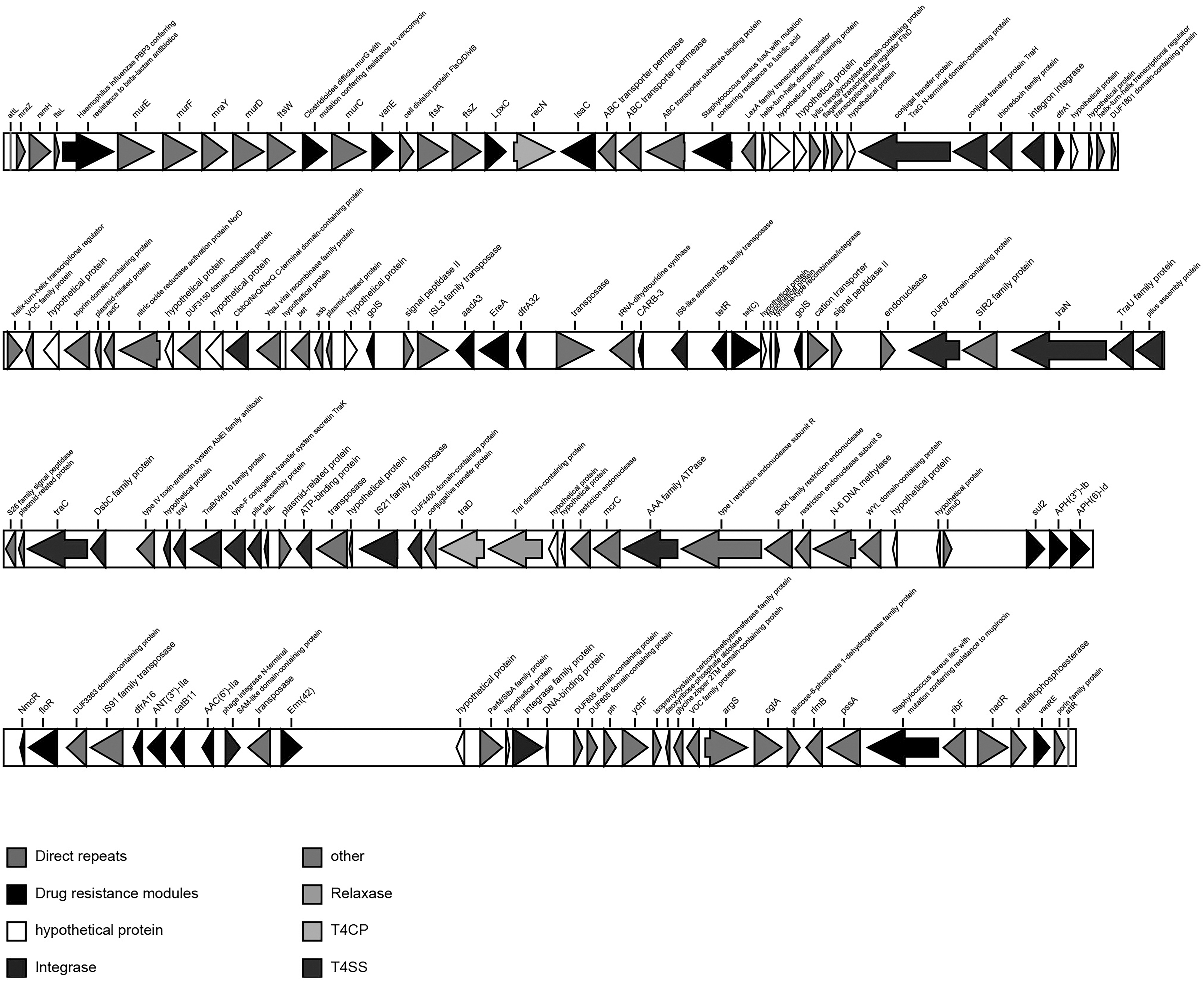
Genetic map of the ICEAplChn2. The direction of the arrow indicates the direction of transcription. Direct repeats, attL and attR are located at the terminals of the 167-kb ICE. Genes are shown in different colors (as shown in the notes).
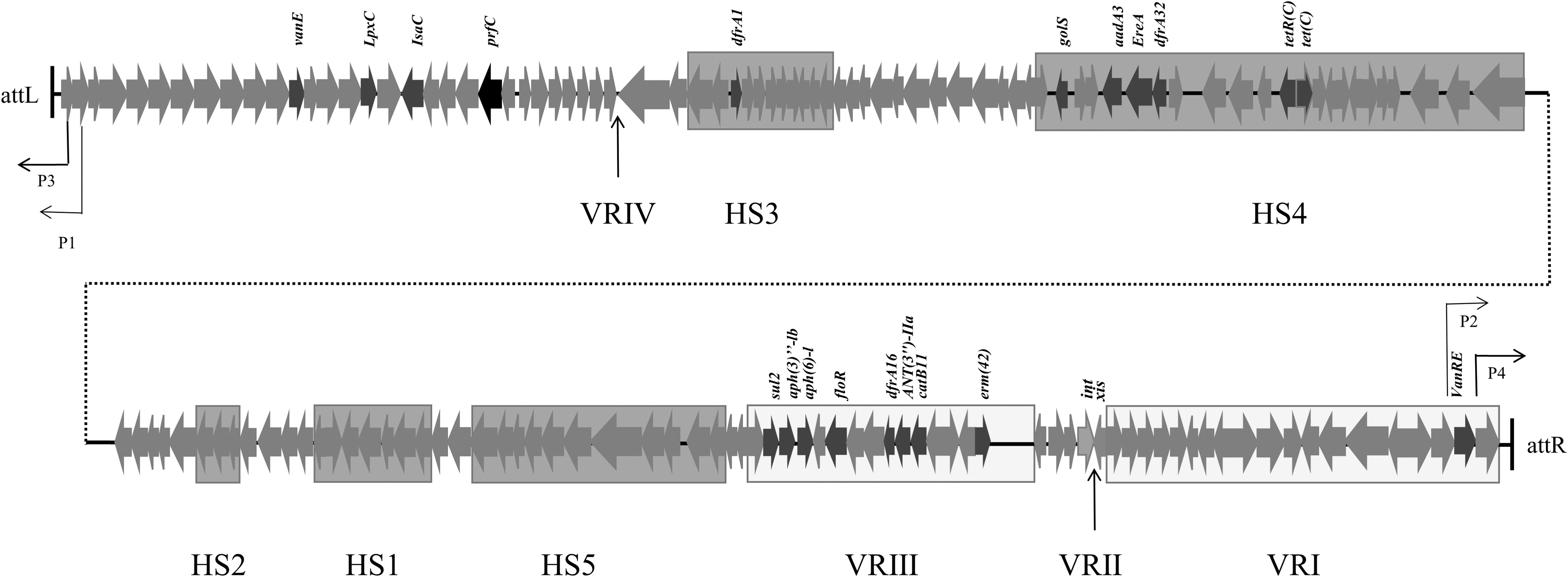
A schematic map of the organization of ICEAplChn2. The transcriptional directions and relative size of the ORFs are indicated by the arrows. A dotted line connects the different parts of the ICEAplChn2. Resistance genes are indicated in italics above. The hotspots (HS) and variable regions (VR) of ICEAplChn2 are shown in different rectangles, respectively. ORFs, open reading frames.
In addition, we also found that HS1-5 and VRI in ICEAplChn2 are different from that of ICEApl2 and ICEAplChn1, the other two identified SXT/R391 family ICEs in A. pleuropneumoniae. In terms of composition of the variable regions, ICEAplChn1 was composed of VRII, VRIII, and VRVI, and ICEApl2 was composed of VRII and VRIII, whereas ICEAplChn2 was composed of VRI, VRIII, and a new insertion region. The results also showed that these three kinds of ICEs represent different antibiotic resistant genes transfer processes.
According to the Blastn alignment, VRI, VRIII, and HS1-5 showed different results, respectively. Among them, VRI of ICEAplChn2 shared 99% nucleotide sequence identity with partial genome of app6 strain (accession number CP026009), but it was not in the sequence of ICEAplChn1. The VRIII region of ICEAplChn2 shared 99.89% identity with both ICEVchind4 and ICEApl2 at 66% query cover. Moreover, HS1, HS2, and HS5 were most similar to that of ICEchCHAHV1003, which was identified from V. cholerae. In addition, HS3 and HS4 of ICEAplChn2 showed high homology with ICEPmiFra1 and ICEVchN1, which also belong to the SXT/R391 family, respectively.
To sum up, ICEAplChn2 showed a high degree of rearrangement in the above regions within the SXT/R391 family. Interestingly, between attL and prfC gene, ATP-binding cassette (ABC) transporter substrate-binding protein and ABC transporter permease are identified, which is the first report in SXT/R391 ICEs. ABC superfamily is widely found in mammals, bacteria, fungi, protozoa, and so on. It carries out the transmembrane transport of a variety of molecules, including proteins, small peptides, amino acids, ions, and antibiotics in an active transport mode. 25 Up to date, ABC superfamily has been found to be associated with multidrug resistance in a variety of bacteria.26,27
In summary, ICEAplChn2 is the largest ICE in the SXT/R391 family and carries the most resistance genes. Compared with the same family of ICEs that were identified from A. pleuropneumoniae, ICEAplChn2 showed a different structural composition. For the variable regions and hotspot regions, ICEAplChn2 also showed a high degree of recombination. Beside of that, A. pleuropneumoniae growth-associated proteins and enzymes including FtsA, FtsW, RecN, and FtsI were also identified in ICEAplChn2. We speculate that this may relate to the adaptation of the ICE to the host bacteria, or to the adaptation of the host bacteria to the environmental edge.
In A. pleuropneumoniae, plasmids have been shown to be important vectors carrying a variety of resistance genes.28–30 However, with the discovery and study of ICEApl1, ICEApl2, ICEAplChn1, and ICEAplChn2, ICEs are also an important carrier of antibiotics resistance genes and play an important role in the horizontal transfer of resistance genes.
Detection of the circular extrachromosomal form of ICEAplChn2
Using A. pleuropneumoniae genomic DNA as a template, four specific primers (Table 3) were used to detect the circular form of ICEAplChn2 (Fig. 5A). The results showed that ICEAplChn2 exists in A. pleuropneumoniae CP063424 in both integrated and cyclized forms, but the amount of ICEAplChn2 presented in the cyclized form is less than that in the integrated form (Fig. 5B).

Detection of the circular form of ICEAplChn2.
PCR Primers Used in This Study
Conjugal transfer of ICEAplChn2
We used the clinical isolate A. pleuropneumoniae serotype 8 as the recipient strain to identify the transferability of ICEAplChn2 by electrotransformation, but we did not obtain transconjugants. For the positive control, ICEHpsaHPS was successfully conjugal transferred to sHPS7 as previous study. 21 Meanwhile, we used another way to test the transferability of ICEAplChn2 as previously described, but we obtained consistent result. 16 The same phenomenon also occurred in ICEApl2 and ICEAplChn1, which were also identified from A. pleuropneumoniae. Amount of A. pleuropneumoniae-related proteins in ICEAplChn2 may affect its ability of conjugation and transfer. Further studies are needed to be performed to reveal specific mechanisms associated with inability of transconjugation.
Conclusion
In summary, a novel SXT/R391-related ICE, named ICEAplChn2, was identified in a clinical isolate of A. pleuropneumoniae CP063424. To our knowledge, ICEAplChn2 is the longest member with the most resistance genes in the SXT/R391 family. Meanwhile, ABC superfamily was found to be inserted into the ICEAplChn2 and possessed a new insertion region, which is the first description in the SXT/R391 family.
Footnotes
Authors' Contributions
J.C. contributed to methodology, collecting data, and writing the original draft. Y.G. and B.Z. were in charge of methodology. Y.L. took care of review and editing.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Key Research and Development Program of China (grant 2022YFD1800902), National Natural Science Foundation (grant 31672565), and the Priority Academic Program for Development at Jiangsu Higher Education Institutions (PAPD).