
Abstract
Candidiasis is a common fungal infection caused by Candida species, with Candida albicans being the most prevalent. Resistance to azole drugs, commonly used to treat Candida infections, poses a significant challenge. Transcriptional activator candidate 1 (TAC1) gene has emerged as a key player in regulating drug resistance in C. albicans. This review explores the structure and function of the TAC1 gene and its role in azole resistance. This gene encodes a transcription factor that controls the expression of genes involved in drug resistance, such as efflux pump genes (CDR1, CDR2, and MDR1) and ERG11. Mutations in TAC1 can increase these genes’ expression and confer resistance to azoles. Various TAC1 gene mutations, mostly gain-of-function mutations, have been identified, which upregulate CDR1 and CDR2 expression, resulting in azole resistance. Understanding the mechanisms of azole resistance mediated by the TAC1 gene is crucial for the strategies in the effective antifungal development pipeline.
Introduction
Candidiasis is a fungal infection caused by various species of Candida, either as a primary or secondary infection. 1 Although noninvasive candidiasis is localized to regions such as the mouth, digestive tract, vagina, nails, and lungs, individuals with underlying disorders or weakened immune systems are susceptible to invasive and potentially fatal infections in visceral organs.2,3 Candida albicans is the most frequently isolated species in Candida infections. However, a steady decrease in its frequency and a progressive increase in non-albicans Candida infections have recently been reported.4,5 Numerous mechanisms contribute to the pathogenicity and drug resistance in Candida infections, influenced by both host and environmental factors and regulated by multiple genes. 6 Despite extensive global efforts to elucidate the functions of genes involved in the mechanisms driving heightened pathogenicity and increased azole resistance, the function of approximately 70% of these genes remains unknown.7,8
Azole resistance in C. albicans primarily involves genes encoding enzymes associated with ergosterol biosynthesis, a crucial component of the fungal cell membrane. Several key genes, including ERG11, CDR1, CDR2, and MDR1, are implicated in C. albicans azoles resistance, particularly fluconazole. 9 An increase in the expression of these genes, often through substitution mutations or alterations in their regulatory genes like MRR1, Transcriptional Activator Candidate 1 (TAC1), and UPC2, can confer azole-resistant phenotypes. 10 The TAC1 gene has recently gained attention because of its pivotal role in controlling the expression of other genes in response to environmental conditions, such as pH, temperature, and nutrient availability. TAC1 plays a crucial role in regulating the production of diverse proteins and enzymes that facilitate yeast’s rapid adaptation to the surrounding environment, metabolism, drug transport, and the regulation of efflux pump genes in C. albicans. Furthermore, it assists in maintaining yeast’s membrane composition to optimize organism efficiency under various environmental conditions.11–13
The TAC1 gene encodes a transcription factor responsible for regulating the expression of multiple genes involved in drug resistance, including efflux pumps (CDR1, CDR2, and MDR1) and ERG11 genes. Mutations in TAC1 can lead to increased expression of these genes and subsequently result in enhanced resistance to azole drugs and a decline in drug effectiveness. 12 Considering the existing knowledge gaps in this field, in this review, we aim to describe various aspects of the TAC1 gene, including its structure and function, and its role in azole resistance.
Data Collection
This mini-review study was conducted using the keywords TAC1 gene, C. albicans, azole resistance, and resistance mechanism, and searching in authoritative databases such as Scopus, PubMed, and Web of Science. Studies not documented in English were excluded from the research. The search period was from May 1, to May 30, 2023, and all studies published from 1996 to May 30, 2023, were reviewed.
Structure of TAC1 Gene
TAC1 is located on chromosome 5 of the C. albicans genome. The structure of the TAC1 gene consists of 1 exon, which spans a genomic region of approximately 2.9 kb. This gene encodes a 981-amino-acids protein with a predicted molecular weight of 105 kDa. Using the SWISS-MODEL server (https://swissmodel.expasy.org), we found that this protein contains several functional domains, including a basic helix-loop-helix (bHLH) domain, a leucine zipper (LZip) domain, and a C-terminal activation domain (Fig. 1). These domains are important for TAC1 binding to DNA and activation of target genes.14,15
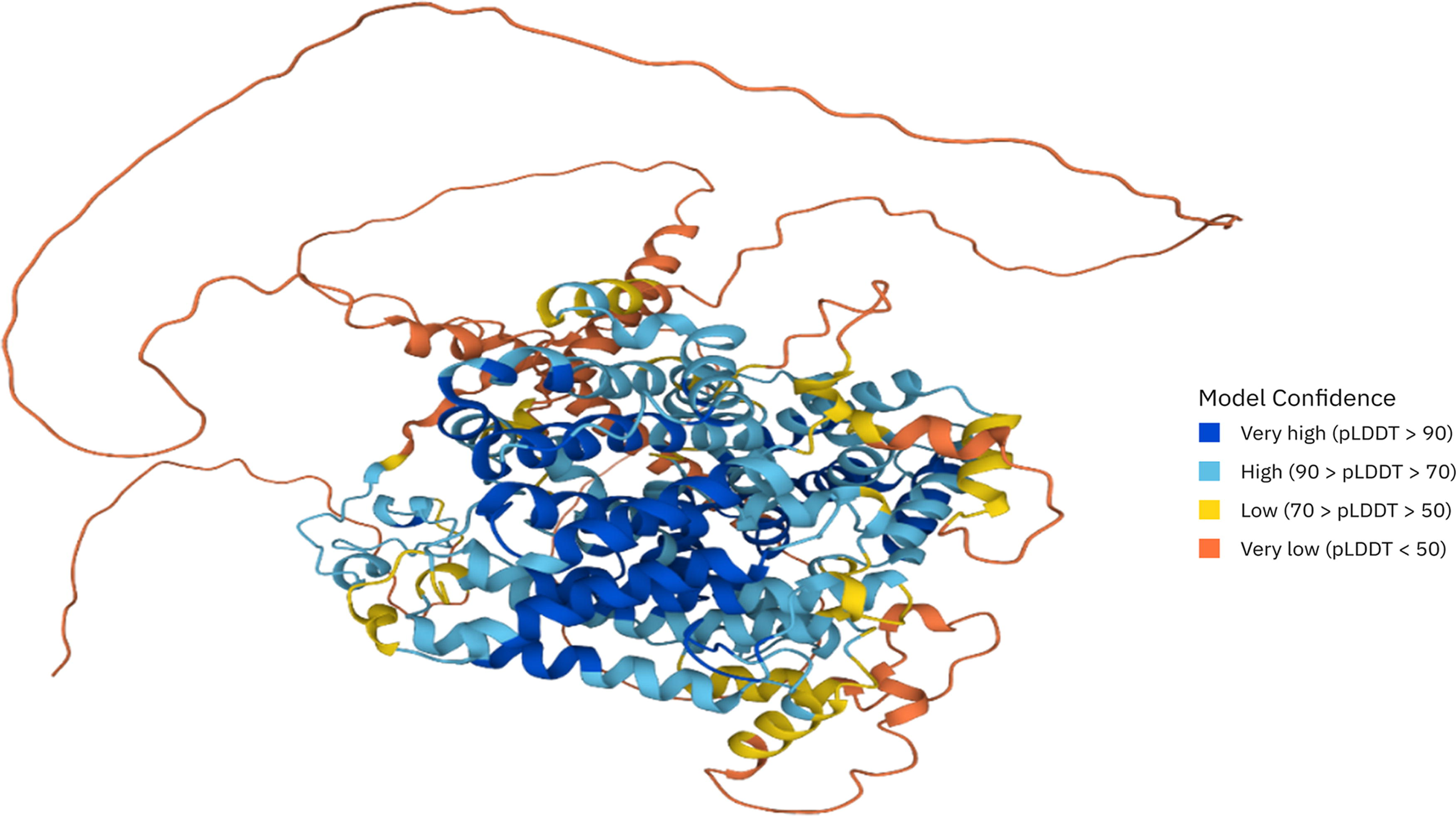
The three-dimensional structure of the TAC1 protein shows different stereochemical regions that have been investigated using the LDDT. The LDDT is a superposition-free score that evaluates local distance differences of all atoms in a model, including validation of stereochemical plausibility. The reference can be a single structure or an ensemble of equivalent structures. LDDT, local distance difference test.
The TAC1 gene exhibits a complex structure encompassing three domains, including an N-terminal DNA-binding domain, a middle homology region, and a C-terminal acid activation domain. 13 The DNA-binding domain facilitates the protein’s ability to bind to specific DNA sequences, whereas the trans-activating domain is responsible for initiating the expression of downstream target genes. In addition, the regulatory domain interacts with various proteins and signaling pathways to modulate the functionality of the TAC1 protein. Consequently, mutations in the TAC1 gene can induce alterations in the regulation of pivotal pathogenic factors, such as drug resistance, biofilm formation, and filament formation in C. albicans, thereby underscoring the significance of this gene.11,16,17
The TAC1 gene of C. albicans encodes a vital transcription factor protein (Tac1p) responsible for regulating multiple genes involved in azole antifungal resistance. When C. albicans is exposed to azole drugs, Tac1p specifically binds to its binding sites located within the promoter regions of target genes, triggering a cascade of events.18,19 This binding facilitates the recruitment of additional transcription factors and co-activators, thereby initiating the transcription of these genes. Consequently, the expression of the ERG11 gene, which encodes the enzyme lanosterol 14α-demethylase, increases within the cell, intensifying its resistance to azole drugs. Furthermore, the augmented production of efflux pumps leads to the expulsion of azole drugs from the cell, thereby reducing their efficacy.20,21
Tac1p interacts with several other proteins to regulate the expression of genes associated with azole resistance. Notably, it interacts with the Mediator complex, which aids in recruiting RNA polymerase II to the target gene promoter regions. In addition, Tac1p interacts with the SWI/SNF complex, a chromatin remodeler that modifies the chromatin structure, rendering the promoter regions more accessible to transcription factors. 22 Moreover, Tac1p cooperates with other transcription factors like Mrr1p and Upc2p to regulate gene expression linked to azole resistance. Mrr1p, a transcription factor, binds to the promoter regions of CDR1 and CDR2 genes, subsequently activating their transcription. Upc2p, on the other hand, regulates genes involved in ergosterol biosynthesis (Fig. 2). It is important to note that mutations in the TAC1 gene, as well as changes in the expression levels and functions of associated genes and transcription factors, can occur, resulting in alterations in azole resistance. 23
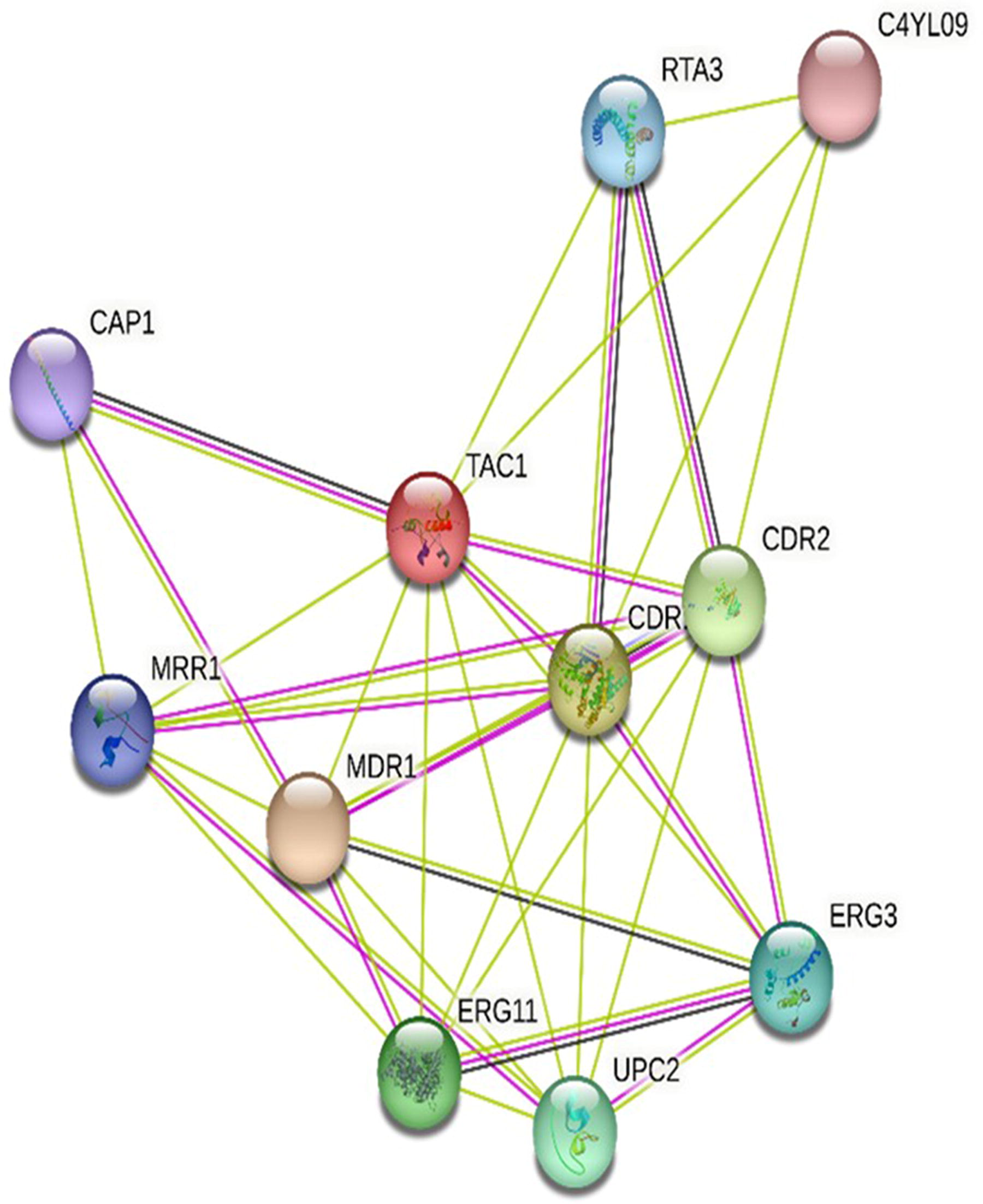
A protein–protein association network between TAC1 and other effective gene products in C. albicans. The interaction network was constructed by using the STRING database version 11.0 (https://string-db.org). The confidence score threshold was set to >0.7, whereas the species was “C. albicans”. Cytoscape version 3.7.1 (www.cytoscape.org), along with the “Molecular Complex Detection Plugin” (MCODE), was used to identify critical modules from the PPI (protein–protein interaction) network. Modules with MCODE score >4.5 were considered significant.
The primary mechanism of azole resistance in C. albicans revolves around the overexpression of multidrug transporters such as CaMDR1, CDR1, and CDR2 genes. CaMDR1 expression is controlled by at least two regulatory cis promoter regions, whereas the regulation of CDR1 and CDR2 includes a basal response element (BRE), a drug response element (DRE), a negative regulatory element (NRE), and steroid response elements. Both CDR1 and CDR2 genes are regulated by a cis-acting element known as the DRE, featuring a consensus sequence of 5′-CGGAWATCGGATAATTTTTTT-3′, which binds to the transcription factor Tac1p. The binding of Tac1p to the DRE leads to the upregulation of CDR1 and CDR2 genes in response to inducers like azole drugs.24,25
Mutations in the TAC1 gene can give rise to gain-of-function (GOF) alleles, resulting in the constitutive overexpression of CDR1 and CDR2 and subsequently enhancing resistance to azole drugs, particularly fluconazole. Table 1 lists various types of DNA mutations investigated in different studies that occur within this gene.
A List of Studies That Have Investigated the Different Types of Mutations That Occurred in the TAC1 Gene
TAC1, transcriptional activator candidate 1.
GOF alleles eventually lead to the constitutive overexpression of CDR1 and CDR2, resulting in increased resistance to azole drugs, notably fluconazole. In the next section, we will discuss the various types of mutations investigated in different studies that have occurred in this gene.
Discussion and Conclusion
Resistance of C. albicans to azole drugs, particularly fluconazole, can occur through diverse mechanisms. One of the primary mechanisms involves the upregulation of genes such as CDR1, CDR2 (encoding ATP binding cassette [ABC] efflux pumps in the cell membrane), and MDR1 (encoding major facilitator superfamily [MFS24] efflux pumps in the cell membrane). This upregulation leads to an increase in the number of ABC and MFS efflux pumps, resulting in the extrusion of azole drugs from the cell. Consequently, the drug concentration inside the cell decreases, giving rise to a resistant phenotype.39,40 The augmented expression of these genes predominantly arises from point mutations occurring in their regulatory factors, including TAC1, UPC2, and MRR2 genes.41,42 Given the crucial role of TAC1 as a significant component of these regulatory factors, the present study aimed to investigate the reported mutations of the TAC1 gene in various studies, as well as its structural and functional characteristics. The pioneering work by Coste and collaborators in 2007 first reported several mutations in the TAC1 gene. 27 These mutations, predominantly GOF mutations, opened up new avenues for exploring the involvement of TAC1 in the development of resistant phenotypes in C. albicans. In 2006, Coste et al. 26 identified a point mutation (N977D) in the TAC1 gene associated with the upregulation of CDR1 and CDR2, resulting in elevated levels of drug resistance. Hyperactive alleles carrying this mutation exhibited dominance over wild-type alleles, and only homozygous strains for the hyperactive alleles displayed high expression levels of CDR1 and CDR2.12,26 In another study, conducted by Coste et al. in 2007, two deletion mutations, ΔM677 and ΔL962-N969, were identified, leading to mitotic recombination and single-gene correction. 27 Another mutation, A736V, was introduced in Coste’s study, causing homozygosity for the GOF mutation and giving rise to two distinct hyperactive TAC1 alleles with the same GOF mutation in these resistant isolates. 27 Furthermore, in 2009, Coste et al. 30 identified and characterized four GOF mutations, namely, E461K, G980E, E841G, and N972D, in the TAC1 gene, indicating that GOF mutations can occur at specific positions in Tac1p. These findings shed light on the structural and functional implications of TAC1 mutations in C. albicans drug resistance.17,28,29,31–35,37,38
In contrast to the aforementioned studies, Dhamgaye et al. 43 conducted a study using RNA sequencing (RNA-seq) to analyze the global expression levels of fluconazole resistance genes in a resistant C. albicans strain and its drug-sensitive counterpart. Surprisingly, they found that resistance in these isolates was associated with the overexpression of CZF1, a gene encoding a transcription factor that inhibits β-synthesis and negatively regulates cell wall integrity. Their RNA-seq analysis also revealed an unknown transcribed region upstream of the TAC1 gene, which serves as the primary regulator of CDR transcription. 43
Moreover, our analysis of gene expression profiles of 12 resistant C. albicans isolates compared with 12 sensitive isolates, based on the array deposited in the NCBI Gene Expression Omnibus (GEO) database with accession number GSE8727, revealed elevated expression levels of CDR1 (log2FC = 1.203, p value = 8.05e-07), CDR2 (log2FC = 4.44, p value = 7.37e-17), IFU5 (log2FC = 1.868, p value = 6.75e-08), and ERG2 (log2FC = 1.076, p value = 4.31e-07).44,45 Furthermore, analyzing the data from 12 resistant isolates compared with that from 3 isolates with a knocked-out TAC1 gene revealed strong suppression of CDR1 and CDR2 expression in the knocked-out strains, even compared with the sensitive strains. This evidence suggests a functional interaction between the ERG2 and IFU5 genes and the TAC1 gene, as well as the CDR1 and CDR2 genes, which are known to be influenced by TAC1 expression.
Given the escalating exposure of patients to various drugs, comprehensive investigation of antifungal resistance, using extensive genomic approaches, is imperative.46,47 Recent advances in high-throughput DNA sequencing, such as next-generation sequencing (NGS), have revolutionized our understanding of fungal biodiversity. NGS enables the study and identification of mixed populations of mutant and wild-type Candida yeasts, providing a large number of sequences reads rapidly. Furthermore, it facilitates the identification of not only the most dominant community but also low abundance or rare species.48,49 Garnaud et al. 36 conducted a study employing NGS to comprehensively evaluate six genes commonly associated with antifungal resistance (ERG11, ERG3, TAC1, CgPDR1, FKS1, and FKS2) in 40 Candida isolates (18 C. albicans, 15 Candida glabrata, and 7 Candida parapsilosis). Through this method, they identified two nucleotide substitutions (C2804T and T2822C) in the TAC1 gene but, unlike FKS1, ERG3, and FKS2 genes, in which multiple resistance–associated mutations were observed, these two novel nucleotide mutations were not associated with resistance in C. albicans species. 36 Most of the studies reviewed did not quantify TAC1 gene expression. Nevertheless, the study by Zare-Bidaki et al. showed that the overexpression of the TAC1 gene was the highest in the resistant group (96.8%) compared with the susceptible group (44.4%), and the mean fold change in the resistant isolates was about 3.1 compared with the susceptible isolates. 18 In another parallel study by Liu et al., the expression of TAC1 gene between different resistant and susceptible isolates was investigated and the fold change was measured between 1.1 and 3.4. 45 However, the TAC1 gene stands out as a key player in generating azole-resistant phenotypes, particularly against fluconazole. Mutations occurring at different regions of Tac1p, as reported in various studies spanning from 1997 to 2023, mostly lead to increased expression of genes involved in controlling Candida cell efflux pumps (CDR1, CDR2), as well as the ERG2 and IFU5 genes. This indicates that mutations in this gene can be linked to diverse mechanisms of C. albicans resistance to azole drugs. Besides conferring resistance in C. albicans, the presence of a mutation in TAC1 gene can have also other effects. For example, it has been shown that homozygous null mutations in this gene can cause different phenotypes such as reduced hyphal growth, decreased resistance to oxidative stress, reduced stimulation to steroid hormones, and reduced filamentous growth. 50
To comprehend these resistance mechanisms fully, scientific advancements and the utilization of new sequencing techniques, such as NGS, are crucial in elucidating effective strategies to counter this resistance phenomenon.
Declarations
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
Footnotes
Authors’ Contributions
A.H.M., A.H., I.H., and O.R. conceived and designed the analysis; A.H.M., M.GH., M.A., I.H. collected the data; A.H.HM., A.H. contributed data or analysis tools; S.S.B., R.G., L.G., O.R. performed the analysis; A.H.M., A.H., M.GH., M.J.B., M.A., R.G., L.G., I.H., O.R. wrote the article; I.H., O.R. review and editing. All authors have reviewed and approved the final version of the article before submission.
Disclosure Statement
The authors declare no commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.