
Abstract
Integrative and conjugative elements (ICEs) are important vectors of lateral gene transfer and contribute to the evolution of bacterial pathogens. However, studies on the transfer among species and the physiological consequences of ICEs are rare. The objective of this study was to investigate the cross-species transferability of newly identified erm(B)-carried ICE in Streptococcus anginosus San95 and its physiological consequences after transfer. The erm(B)-carried ICE, characterized by a triple serine integrase module, integrated into hsdM genes, thus designated ICESan95_hsdM. Analysis of ICESan95_hsdM revealed 32 additional ICESan95-like ICEs in the available NCBI genome (n = 24) and sequence of clinical isolates (n = 8). Polymerase chain reaction (PCR) was used to evaluate the 467 clinical isolates, of which 84 were positive for core genes (integrase, relaxase, and T4SS genes) of ICESan95_hsdM. Cross-species transfer experiments demonstrated that ICESan95_hsdM could transfer from S. anginosus to different streptococcal and enterococcal recipients. Growth and competitive culture assays showed acquisition of ICESan95_hsdM incurred no fitness cost. Our work discovered a group of ICEs in Streptococci and Enterococci. For the first time, we demonstrated the broad cross-species transferability to different species or genera of ICEs with no fitness cost that enables commensal S. anginosus to deliver antimicrobial resistance genes to other streptococci and enterococci.
Introduction
Lateral gene transfer (LGT) is considered a key driving force in gene exchange between different bacteria. 1 LGT enables different bacteria to rapidly acquire new properties to enhance their adaptability environmental changes. Most mechanisms of LGT rely on diverse mobile elements, such as prophage, conjugative plasmids, and integrative and conjugative elements (ICEs). Over the last two decades, ICEs have been increasingly identified in chromosomes.2,3
ICEs are a class of self-transmitting mobile elements consisting of different functional modules, including integrase modules (integrase and XIS genes), conjugation modules (relaxase and type IV secretion systems [T4SS]), and diverse cargos. Among them, the integrase module is the most specific, and typically determines the integration site or sites of an ICE. 4 Based on the diversity of integrase modules, four classes of ICEs have been identified, tyrosine integrase, single serine integrase, triple serine integrase, and DDE transposase ICEs. 5
Diverse ICEs reside in the chromosomes, but can be excised from the chromosome and transferred to another cell through conjugation. During its life cycle, ICEs are excised and form a cyclic intermediate, and one strand of the ICE is transferred to a new host by rolling circle replication. The one strand of ICE, then, can synthesize the complementary DNA strand and integrate into specific site/sites by ICE integrases. During this process, diverse cargo genes, such as antimicrobial resistance (AMR) genes, 6 heavy metal resistance genes, and other functional genes, are transferred, 7 conferring ICE-specific properties to the host.
Previous studies have shown that the prolonged use of macrolides has resulted in increased resistance of Firmicutes to macrolides, lincosamides, and streptogramin group B (MLSB). 8 In up to 116 related mechanisms for the phenotype of MLSB, the erm(B) genes encoding methylase mediating target site modification is the most predominant one in streptococci and some other Gram-positive bacteria. 9
To date, numerous erm(B)-carrying ICEs have been reported, whose transferability also have been demonstrated through conjugation transfer experiments. For instance, ICESag37, an ∼73 kb ICE, was originally detected in Streptococcus agalactiae, 10 while ICESsuYZDH1 and some members of the ICESa2603 family (Tn5252 group), such as ICESsuYSB17, have been found in Streptococcus suis11,12; ICESp1116 (TnGBS2 family), an erm(B)-carried ICE, was discovered in erythromycin-resistant Streptococcus pyogenes. 13 Very recently, a mosaic ICESdyV220 comprising prophage fragments and an ICE was also found to carry erm(B), indicating that the ICE may be constantly evolving as it is transferred. 14 Despite the diverse range of ICEs throughout genomes, their hosts and transmittance range seem to be limited, and little is known about their interspecies or intergeneric transmission capacities.
In this study, we identified an erm(B)-carrying ICE in a clinical Streptococcus anginosus San95 isolate (ICESan95_hsdM), whose transferability has not yet been demonstrated. Comprehensive analysis of publicly available genome and clinical isolate sequences revealed that ICESan95-like ICEs are diverse and broadly distributed among multiple streptococcal and enterococcal species. Through conjugation transfer experiments, we assessed the cross-species transferability of this ICE from S. anginosus to S. agalactiae, Enterococcus faecium, and Enterococcus faecalis and evaluated the fitness costs. The dissemination of ICESan95_hsdM or related ICEs might be a new method of AMR gene delivery among streptococci and enterococci.
Materials and Methods
Bacterial strains and culture conditions
We obtained 27 previously sequenced strains of clinical S. anginosus that displayed erythromycin resistance (minimal inhibitory concentration [MIC] ≥1 mg/L). Of these, five harbored erm(B) gene and thus met our inclusion criteria; their principal features are summarized in Supplementary Table S1.
All isolates used in the study were obtained from isolates stored at Huashan Hospital in 2022. A total of 467 clinical isolates were available across two genera, that is, Streptococcus (161 S. agalactiae, 204 S. anginosus, 4 S. pyogenes, and 8 Streptococcus pneumoniae) and Enterococcus (62 E. faecium and 28 E. faecalis). Only strain codes were used in data analysis, and no other personal information was needed. Therefore, ethical approval and consent were not required for this study.
S. anginosus and S. pneumoniae were grown on Todd-Hewitt broth with 5% (v/v) sheep blood at 37°C for 24 hours; S. agalactiae and S. pyogenes were grown on Todd-Hewitt broth with 5% (v/v) sheep blood for 12 hours at 37°C; and E. faecium and E. faecalis were grown on Brain-Heart Infusion broth at 37°C for 8 hours.
Antimicrobial susceptibility test
An antimicrobial susceptibility test was performed using the broth microdilution method and the S. agalactiae ATCC13813 was used as the quality control strain. The breakpoint of each antibiotic was in accordance with the Clinical and Laboratory Standards Institute (CLSI) standard. 15
Analysis of ICEs in clinical isolates and available NCBI genomes
To identify similar ICEs among total 467 clinical isolates, each isolate was resuspended in 1 mL sterile water for 5 minutes at 94°C to generate a DNA template for polymerase chain reaction (PCR) analysis. The primers were designed against one of three integrases, relaxase, and virB4 of eight T4SS genes (Supplementary Table S2). Strains possessing all these genes were thought to contain at least one ICESan95-like ICE. Subsequently, strains with ICE from different species were randomly sequenced for further validation (total of four strains S. anginosus San246, S. agalactiae Sag22, E. faecium Efm394, and E. faecalis Efa12).
ICESan95-like ICEs in genomes were identified using a published method. In brief, strains where integrase showed >60% sequence identity for another and had other core structure synteny could be classified into one group. 16 In available NCBI genomes and sequences of clinical isolates (27 pre-existing S. anginosus sequences and 4 validation sequences), 3 integrases (PP359593, 50028-54845), relaxase (36229-37920), and all T4SS genes (7607-16909) were used as query terms for BLASTn. The parameter reference to online tool ICEFinders' ICE comparison function (bioinfo-mml.sjtu.edu.cn/ICEfinder): sequence identity of core genes (integrases, relaxase, and all T4SS cluster) >60%; maximum distance between genes <10 kb. ICESan95-like ICEs were integrated within several genes, where they caused original gene disruption into two open read frames (ORFs); thus, ICE delimitation was determined based on the interrupted ORFs.
The ORFs disrupted by ICE integration were used as query terms for BLASTn in NCBI to obtain intact ICE-targeted genes. Phylogenetic analysis was then performed using ICE-targeted genes. Evolutionary history was inferred using the Maximum Likelihood method based on the Tamura-Nei model. 17
Conjugation transfer experiments
Two conditions had to be met for selection of recipients in clinical isolates, that is, the isolates had to display a resistance phenotype opposite to that of the donor and had to contain at least one empty integration site for ICESan95_hsdM. Strains sensitive to erythromycin and clindamycin, but resistant to either levofloxacin or tetracycline, were selected to proceed to the next step of screening. The primers for putative integration site detection were designed based on genes that have been inserted by ICESan95-like ICEs (Supplementary Table S2). One strain that met inclusion criteria was randomly selected from each species as recipient.
Filter mating experiments were performed as described previously. 18 Strains that were used in conjugation transfer experiments are listed in Table 2. The S. anginosus San95 (ERYR, CLIR, TETS, LEVS) was used as the donor. The S. agalactiae SagR40 (ERYS, CLIS, TETS, LEVR) with one putative integration site (ICESag22_type II MTase inserted gene); E. faecalis EfaR372 (ERYS, CLIS, TETR, LEVS) with one putative site (ICEEfa12_hsdM inserted gene); and E. faecium EfmR138 (ERYS, CLIS, TETS, LEVR) with one putative site (ICEEfm394_hsdM inserted gene) were used as recipients.
The donor and recipients were grown to optical density [OD]600 0.5–0.8 and mixed at a ratio of 1:10 (donors: recipients). The mixed bacterial solution was evenly coated on a nitrocellulose membrane and incubated for 4 hours. To rule out the influence of DNA transformation, the mating process was carried out in the presence of 10 mg/mL DNase I. The strains were scraped from the nitrocellulose membrane and spread on selection medium after dilution for 24 hours. Transconjugants from S. agalactiae SagR40 and E. faecium EfmR138 were selected using 50 μg/mL erythromycin, 50 μg/mL clindamycin, and 20 μg/mL levofloxacin, while transconjugants from E. faecalis EfaR372 were selected using 50 μg/mL erythromycin, 50 μg/mL clindamycin, and 30 μg/mL tetracycline.
The transfer of the MLSB phenotype was determined through antimicrobial susceptibility tests of the donor, recipients, and transconjugants. The transfer and integration of ICESan95_hsdM were confirmed through PCR detection of erm(B) and integrase gene. The ICE integration sites were determined using PCR (Px1-P2 and P3-Px4) as well (Supplementary Table S2). To avoid false positives, the PCR products of each step were sequenced for validation. The recipients and transconjugants were further confirmed by whole genome sequencing. Transfer frequency was calculated by dividing the number of transconjugants by the number of donors. Each test was repeated in triplicate.
Competitive tests using growth difference and competitive culture
The fitness cost of ICE acquisition was evaluated using comparative growth and competitive culture tests as described previously. 19 In the comparative growth assays, the recipients and their transconjugants were adjusted to exhibit the same density (OD600 = 0.1), diluted to a ratio of 1:200 with fresh medium, and incubated at 37°C and 150 rpm for 24 hours. The OD600 value was measured every hour until stable periods were reached. Tests were performed in triplicate.
The recipients and their transconjugants were adjusted to the same density and mixed at a 1:1 ratio. At 0 and 24 hours, the mixed solution was cultured and counted on counting medium with or without 50 μg/mL erythromycin and clindamycin. The same procedure was repeated for five successive generations. The fitness value (W) was calculated using Equation (1):
where Ri and Si represent the number of transconjugant and recipient cells at 0 hour, while Rf and Sf represent the number of transconjugant and recipient cells at 24 hours.
DNA sequencing and sequence analyses
The genomes of target isolates were extracted using a QIAGEN Midi Kit (Qiagen, Hilden, Germany), according to the manufacturer's instructions. The whole genome sequencing of short reads was performed on all experimental strains in this study at BGI Genomics (HiSeq X; Illumina, San Diego, CA). Sequencing data were quality trimmed, and Illumina Nextera indexes were removed using Trimmomatic v0.39.82. The high-quality reads were de novo assembled using Spades v3.9.034. Genome annotation was performed using the Prokka 1.14.6. or online tool rast. All recipients were further subjected to long-read whole genome sequencing using the MinION Sequencer (Nanopore, Oxford, United Kingdom). Complete genomes were assembled using Canu v2.26 and then polished using Polypolish v0.5.07.
Accession numbers
The sequence of each isolate for conjugation transfer experiments in this study has been deposited in GenBank, including ICESan95_hsdM (PP359593, 4872-54845); donor S. anginosus San95 (JAXIID000000000); recipients S. agalactiae SagR40 (CP138370), E. faecium EfmR138 (CP142725), and E. faecalis EfaR372 (CP141754); and transconjugants S. agalactiae SagR40_TC (CP139638), E. faecium EfmR138_TC (JAXIIC000000000), and E. faecalis EfaR372_TC (JAXIIB000000000).
Results
Characterization of ICESan95_hsdM
The five erm(B)-carrying S. anginosus strains displayed high-level erythromycin and clindamycin resistance (MIC ≥128 mg/L). Further analysis of their genomic environment revealed that erm(B) in four strains was carried by a typical ICESa2603 family ICE (characterized by a tyrosine integrase module and Tn5252-like conjugation module). However, in the other isolate (S. anginosus San95), erm(B) was found within an uncommon genomic element that contained three integrases, a relaxase, and a cluster of conjugation-related genes (predicted by ICEFinder), which were inserted into a gene annotated as hsdM (encoding type I methyltransferase); thus, this element was identified as an ICE and named as ICESan95_hsdM.
ICESan95_hsdM was 49.974 kb in length and contained 45 putative ORFs. Only 3 bp direct repeats (GGG - GGG) flanked the ICE. The integration module of ICESan95_hsdM comprised three tandem integrase genes, whose arrangement was similar with those of the triple serine integrase of ICESsuYZDH1. 11 However, their integration modules did not have obvious nucleotide or amino acid identities, indicating that they belonged to completely different groups of ICEs. Further analysis of the integrase module of ICESan95_hsdM revealed some amino acid homology, suggesting that the three tandem integrase genes were probably derived from one ancestor with two successive duplications.
Comparative analysis of the whole ICE sequence showed the ICESan95_hsdM core modules were more than 60% identical with those of ICE10750-RD.2, whose transferability has not yet been confirmed, 20 and a nontransferable Tn1806; 21 and more than 80% identical with a silicon predicted ICESanMAS624 in S. anginosus. 5 All ICEs had been inserted themselves into the hsdM gene (Fig. 1a). Notably, the relaxase gene of ICE10750-RD.2 and snf2 within Tn1806 were disrupted by reverse transcriptase genes; thus, structural damage may explain why these ICEs had not demonstrated transferability. Meanwhile, the erm(B)-carrying segment of ICESan95_hsdM was identical to the region containing the erm(B) gene in cTn6002 of Streptococcus cristatus. 22 ICESan95_hsdM was also related to an internal element inserted into the snf2 gene within the mosaic element ICESag37741 (ALQQ01000033). 23

Comparison analysis of ICESan95_hsdM.
Widespread distribution of ICESan95-like ICEs between Streptococci and Enterococci
In the NCBI database, 24 ICESan95-like ICEs (including six identical ICEs and three reported ICEs) were identified within the genomes of five S. agalactiae, four S. anginosus, four S. pneumoniae, five S. pyogenes, two unknown Streptococci, and four E. faecalis. These ICEs shared similar core modules and carried multiple AMR genes, such as erm(B), erm(A), or erm(TR) (MLSB), and tetM (tetracycline resistance), ant(6″)-aph(2′) (aminoglycoside resistance), or mef(A)-mel (erythromycin resistance) (Fig. 1).
The locations, accession numbers, lengths, and AMR genes in ICEs are listed in Table 1. Of the 467 clinical isolates, 84 isolates (24/161 of S. agalactiae, 44/206 of S. anginosus, 6/28 of E. faecium, and 10/62 of E. faecalis) contained at least 1 ICESan95-like ICE. Subsequently, from 4 sequencing isolates and 27 of erythromycin-resistant S. anginosus, 8 additional AMR-free ICEs were identified from sequences of 5 strains of S. anginosus (designated ICESan08_snf2, ICESan26_snf2, ICESan100_snf2, ICESan188_snf2, and ICESan246_snf2), 1 S. agalactiae (ICESag22_type II MTase) strain, 1 E. faecium (ICEEfm394_hsdM) strain, and 1 E. faecalis (ICEEfa12_hsdM) strain (Figs. 1b and 2).
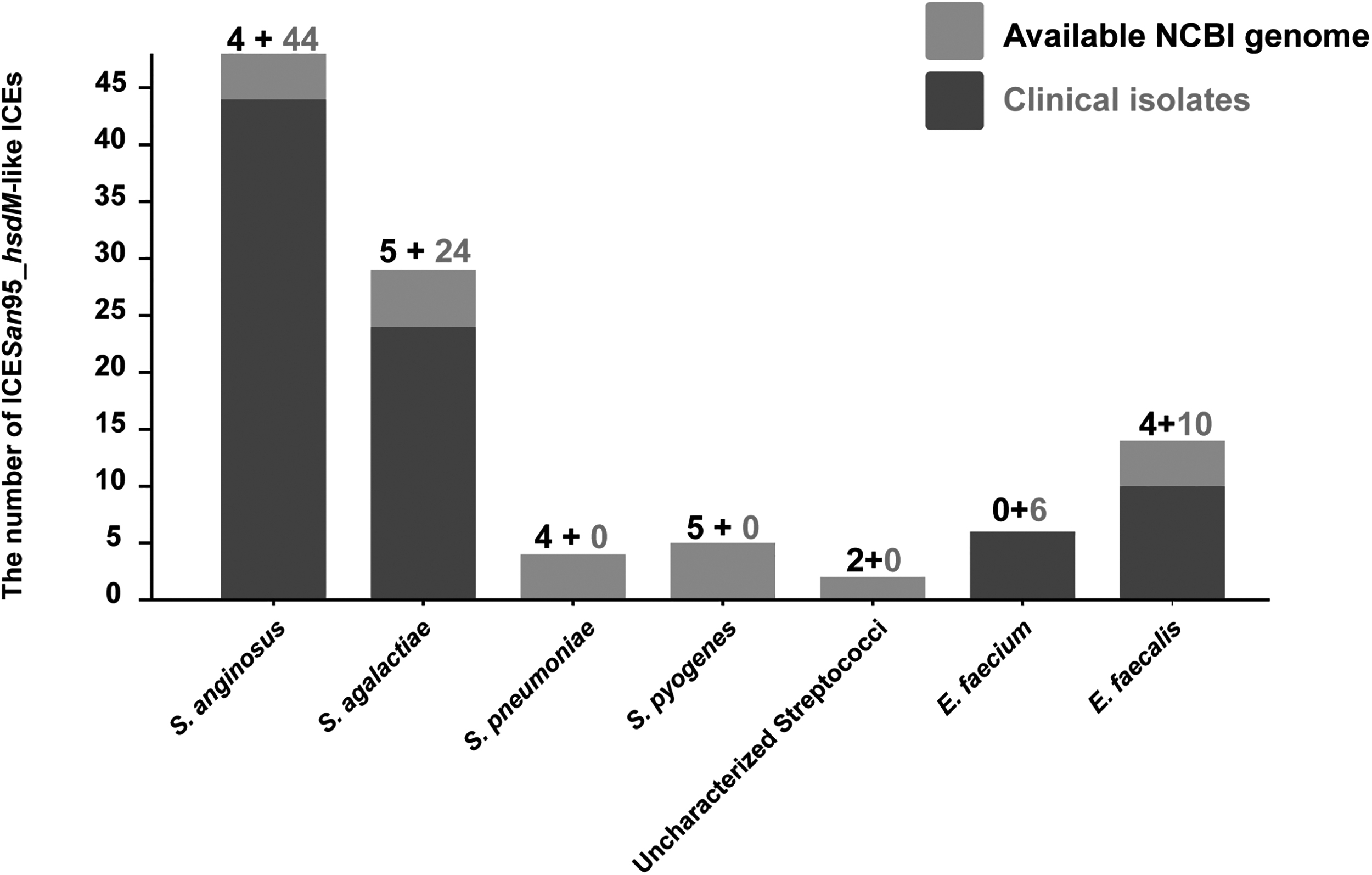
Distribution of ICESan95-like ICEs in clinical isolates and NCBI available genomes, identified from clinical isolates (n = 84) and the NCBI database (n = 24). The figure at the top of the bar graph represents the number of ICEs identified in the available NCBI genome data and clinical strains.
Characteristics of 33 ICESan95-like Integrative and Conjugative Elements
The target genes were disrupted by ICE integration, while intact genes and annotations were obtained from others genomes: aCP041968, FPL13_06075; bCP041968, FPL13_05065; cCP029749, DLM82_06795; dCP041261, FKW92_02830; eCP051005, GRB94_13135; fCP031556, DY328_11435; gCP053789, GE023_005635; hCP029749, DLM82_01500; iCP007573, DK43_04175; jCP003861, SANR_1187; kCP040798, FDP16_01455; lCP068060, I6J12_00250; mCP125360, OPT59_08340; nAP018937, hsdM; oCP033907, EHF40_08525; pCP044093, FOB76_05570; qCP118057, PUW58_06720.
AMR, antimicrobial resistance; ICE, integrative and conjugative element.
Comparative analysis revealed that the hsdM and snf2 genes were the two primary integration targets for the ICESan95-like ICE group. Seventeen ICESan95-like ICEs were inserted into ORFs annotated as hsdM or related products, and fourteen were inserted into snf2-related genes (Table 1). However, neither hsdM nor snf2 was a specific gene in a genome. Specifically, hsdM represented a class of genes that express type I methyltransferases, which are vital components of the Restriction-Modification (RM) system 24 and snf2 encoded SNF2-related proteins that play a key role in DNA reparation, expression, and duplication. 25 Thus, despite being annotated as hsdM or snf2, no significant sequence identity was observed between most of them (Fig. 3a).
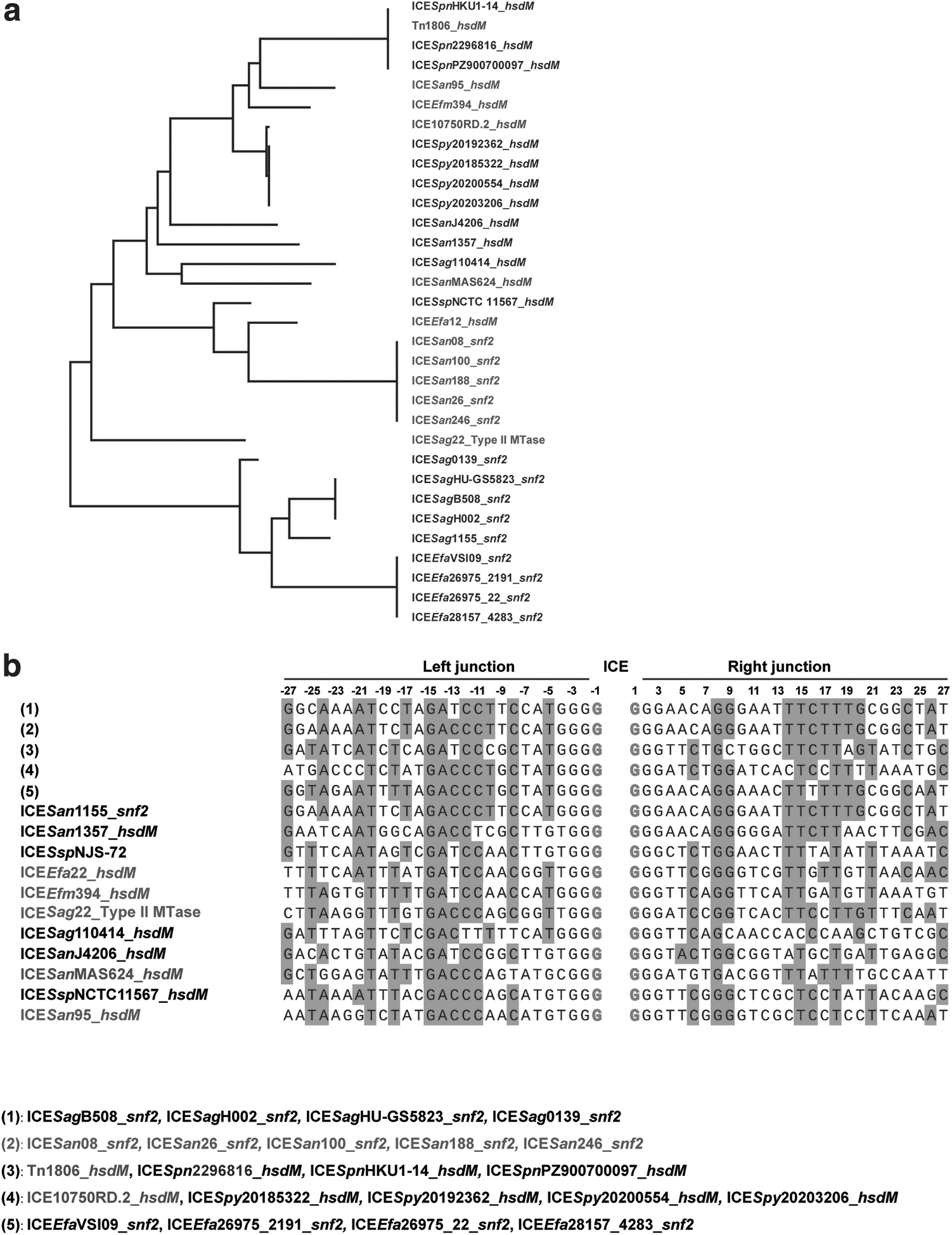
Comparative analysis of ICE integration sites.
In addition, one ICE (ICESag22_type II MTase) derived from clinical isolates was found inserted into an ORF annotated as type II restriction endonuclease, while another ICE (ICESspNSJ-72) was inserted into an intergenic position (CP060489, between H8787_07535 and H8787_07295). Further comparative analysis of ICE target position sequences did not reveal any obvious consensus sequence (Fig. 3b). Taken together, these results suggested that these ICEs probably represent a broad ICE group in different niches; however, the integrative specificity of this group of ICEs appears limited.
Conjugative transfer and excision/circular form of ICESan95_hsdM
Four primers (Pa1, P2, P3, and Pa4) were used to test for the intermediate circularization form of ICESan95_hsdM, which is necessary for ICE transfer. PCR confirmed the excision (Pa1-Pa4) and circular (P2-P3) forms of ICESan95_hsdM (Fig. 4a). In the conjugative transfer of ICESan95_hsdM to the recipients S. agalactiae SagR40, E. faecalis EfaR372, and E. faecium EfmR138, transconjugants were obtained with frequencies of 1.19 × 10−6, 6.32 × 10−7, and 7.60 × 10−7, respectively, per donor (Table 2).

PCR analysis of the donor and transconjugants.
Characteristics of the Strains Used or Obtained During the Conjugation Experiments
The transfer frequency was calculated using CFUs of transconjugant/donor. The data were averaged from five independent assays.
The fitness value (W) was the average of five consecutive passages.
ICE in short-read sequencing might be assembled in two contigs.
CFU, colony-forming unit; CLI, clindamycin; ERY, erythromycin; LEV, levofloxacin; MIC, minimal inhibitory concentration; TET, tetracycline.
All transconjugants showed high levels of erythromycin and clindamycin (>128 μg/mL) resistance as donor, indicating that the MLSB resistance phenotype was transferred to erm(B)-free recipients. PCR detection of integrase and erm(B) genes in transconjugants showed that the erm(B)-carrying ICESan95_hsdM was successfully transferred to different recipients (Fig. 4b). Further detection of ICE insertion sites showed that ICESan95_hsdM was integrated into anticipant target sites in S. agalactiae SagR40_TC and E. faecium EfmR138_TC.
However, in the transconjugant E. faecalis EfaR372_TC, no ICE was detected in the anticipant site, indicating ICESan95_hsdM had integrated into another unexpected site. Genomic sequencing of recipients and transconjugants confirmed that ICE was integrated within the anticipant genes of S. agalactiae SagR40 (SE933_06840) and E. faecium EfmR138 (VFJ37_09820). However, the ICE in E. faecalis EfaR372 was integrated at an unexpected gene (UXJ07_08525), not the anticipant gene (UXJ07_12380). Information regarding phenotypes, origin, accession numbers, and the location of ICESan95_hsdM in the genome of donor and transconjugants is listed in Table 2.
Other features, such as the fitness cost of the acquisition of ICESan95_hsdM, were also investigated. The fitness cost was evaluated by comparing growth and competitive cultures. Growth curves showed no difference between the growth of recipients and their transconjugants (Supplementary Fig. S1). The competitive cultures revealed that the transconjugants had a fitness value (W) of 1.03–1.06, indicating the recipients and their transconjugants had equal growth (Table 2).
Discussion
The long-time use of macrolide antibiotics in anti-infective treatment has led to increasing resistance rates of bacteria to erythromycin and clindamycin. In 2019, the U.S. Centers for Disease Control and Prevention (CDC) reported 21 drug-resistant threats in the United States, including “Serious threats” of drug-resistant S. pneumoniae and vancomycin-resistant enterococci, as well as “Concerning threats” of erythromycin-resistant S. pyogenes (GAS) and clindamycin-resistant S. agalactiae (GBS). 26
S. anginosus, as previously neglected species, is a part of the Streptococcus milleri group and subgroup of viridans group streptococci (VGS), which includes three different species, S. anginosus, Streptococcus intermedius, and Streptococcus constellatus. Improved diagnostic methods have identified an increasing number of life-threatening infections associated with S. anginosus, including pyogenic infections, bacteremia, and chronic maxillary sinusitis. 27 As opportunistic pathogens, S. anginosus are often detected in mixed infection cases as well. 28 A previous study suggested that VGS may act as important reservoirs of resistance genes for the more pathogenic streptococci. 29 However, risks of antibiotic resistance and resistance gene delivery from S. anginosus to other species have been rarely studied.
In streptococci, the ICESa2603 family (Tn5252 group), one of the broadest ICE groups, mediates the spread of multiple AMR genes among streptococci. 30 In our study, we identified an unusual erm(B)-carrying ICESan95_hsdM in clinical S. anginosus, whose transferability had not been demonstrated. Unlike ICESa2603, which contains a tyrosine integrase and mediates ICE integration into a 15 bp att sequence downstream of rplL, ICESan95_hsdM is characterized by triple serine integrase and integration into hsdM genes. The presence of Tn1806 in S. pneumoniae, ICE10750-RD.2 in S. pyogenes, ICESan95-like ICE within the mosaic element of ICESag37741 in S. agalactiae, and ICESanMAS624_hsdM in S. anginosus implies that ICESan95_hsdM probably is a class of common genomic element between major streptococci and S. anginosus.5,20,21 Meanwhile, hsdM and snf2 are two previously unreported hotspots for exogenous elements.
ICEs similar to ICESan95-like ICEs were broadly distributed between streptococci, and even enterococci. Searches of the whole ICESan95_hsdM sequence from available NCBI genomes revealed up to six identical ICEs from S. anginosus, S. pyogenes (uploaded from United States), and an unknown streptococcal species (from China), indicating that ICESan95_hsdM clone likely underwent a worldwide transfer. Moreover, the identification and characterization of ICESan95-like ICEs in databases and 467 clinical isolates revealed that they represent a group of ICEs that shares a similar backbone.
In addition to the MLSB resistance gene erm(B), many resistance genes, such as tetM for tetracycline resistance, ant(6″)-aph(2′) for aminoglycoside resistance, and mef(A)-mel for erythromycin resistance, were found to be carried by this group, which provides ecological advantages for their host. However, ICEs lacking AMR genes were also present in this group. These ICEs may confer unknown selective advantages that allow them to become widely transferred and presented; however, additional evidence is needed. Another feature for this ICE group is its relatively limited integration specificity. These ICEs were integrated into two genes, hsdM and snf2; however, they varied considerably, and no obvious conserved sequence was observed.
Interestingly, the function of their products was similar, including methyltransferase and SNF2 proteins. Thus, triple serine integrase-mediated ICE integration is preferential rather than specific. However, the mechanism for their preferential integration is unclear. Some relatively conserved nucleotides (position −24, −20, −15 to −10, and 8, 9, 12, to 20 of Fig. 4b) may limit the randomness of integration sites. Importantly, this limited integration specificity enables ICEs with similar triple serine integrases to integrate into a broader genome, thereby facilitating their further dissemination.
In this study, for the first time, we demonstrated the cross-species conjugative transfer of ICESan95_hsdM carrying erm(B) from S. anginosus to S. agalactiae, E. faecium, and E. faecalis. This transfer also allows the lateral transfer of the MLSB phenotype. As predicted by bioinformatics analyses of ICESan95-like ICEs, ICESan95_hsdM was preferentially integrated into the hsdM of recipients S. agalactiae SagR40 and E. faecium EfmR138.
Surprisingly, the transfer allowed the integration of ICESan95_hsdM into another unexpected hsdM site in recipient E. faecalis EfaR372. These results are consistent with the conclusion that ICE integration is preferential rather than specific. However, we have to be recognized that our detection of transconjugants is limited, as many unexpected sites may present in recipients. These preferential sites can obtain ICE and generate different transconjugants, far more than we have detected during ICE transfer. Moreover, the transfer of ICESan95_hsdM does not appear to be limited by species, as they preserved transfer efficiency across recipients (ranging from 6.32 × 10−7 to 1.19 × 10−6 per donor).
The hsdM genes, together with the restriction endonuclease gene hsdR and S subunit hsdS, constitute the R-M system cluster, whose products play a key role in defense against exogenous DNA invasion.23,31 Integration of ICEs into hsdM genes may result in two consequences. First, disruption of the hsdM structure may result in inaction of the RM system, thereby impacting the new host.
However, no fitness cost was observed between recipients and transconjugants, suggesting that disruption of hsdM caused very limited damage to the bacteria. ICEs or other RM systems in the genome may provide compensatory functions. Second, the integration of ICEs into hsdM may result in the accumulation of ICEs within other elements, as hsdM or other types of methyltransferases are abundant in genomic elements as well. These hsdM-target ICEs might interact more frequently with other genomic elements, which might be an important mechanism leading to the emergence of larger and more complex ICEs. However, it remains to be further investigated.
Conclusion
In conclusion, the low fitness cost and broad host range could benefit the dissemination of these ICEs. S. anginosus is typically a commensal bacterium involved in multiple concurrent infections, and its frequent contact with other pathogens may lead to the transfer of ICE and provide a new way for the transmission of resistance genes among species. Widespread S. anginosus could act as an ICE reservoir, thereby enhancing the possibility of some streptococci and enterococci to acquire drug-resistant genes.
Footnotes
Acknowledgments
We thank Jinjin Zhao, Qingqing Du, and Xiaofei Yi for their help in data collection. We are most grateful to Professor Minggui Wang for critically reviewing the study proposal.
Authors' Contributions
S.Y. contributed to conceptualization, validation, methodology, data curation, formal analysis, visualization, investigation, resources, and writing original draft; K.Z. was involved in formal analysis, data curation, and writing review and editing; and X.X. was in charge of conceptualization and writing, supervision, and funding acquisition.
Disclosure Statement
We declare no conflicts of interest in this work.
Funding Information
This work was supported by grants 81991531, 81991530, and 81971978 from the National Natural Science Foundation of China.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.