
Abstract
Background:
Interleukin-6 (IL-6) is a proinflammatory cytokine involved in the pathogenesis of chronic inflammatory diseases such as coronary artery disease (CAD). The −174 IL-6 G/C promoter polymorphism influences mRNA levels and protein expression and is implicated in CAD. The Indian population in South Africa, unlike the black community, has a high prevalence of premature CAD. This polymorphism has not been fully explored in this population. The present study assessed the −174 IL-6 G/C polymorphism in young Indian patients with angiographically documented CAD and compared them with age- and gender-matched Indian and black control subjects.
Methods:
Polymorphic variants were assessed by polymerase chain reaction–restriction fragment length polymorphism, and IL-6 levels were measured using enzyme-linked immunosorbent assay (ELISA).
Results:
The −174 IL-6 C allele was found with a higher frequency (23%) in the total Indian group compared to 2% in the black participants [P<0.0001, odds ratio (OR)=0.05, 95% confidence interval (CI) 0.018–0.14). The difference in frequency was more pronounced when Indian controls were compared to black controls (29% vs. 2%, respectively) (P<0.0001, OR=0.05, 95% CI 0.02–0.17). A significant association between the −174 IL-6 G allele and CAD was found in Indian patients compared to Indian controls (84% in cases vs. 71% in Indian controls; P=0.043, OR=0.47 95% CI 0.23–0.95). Levels of IL-6 in circulation were higher in black controls (6.62±0.63 pg/mL) compared to Indian controls (2.51±0.57 pg/mL) and CAD patients (1.46±0.36 pg/mL) (P<0.0001). Levels of IL-6 were higher in all groups with homozygous −174 IL-6 C alleles, but only significant in the healthy Indian control group (GG 3.73±0.94 pg/mL vs. GC/CC 0.89±0.5 pg/mL, P=0.0001).
Conclusion:
The presence of the IL-6 −174 G allele influences levels of IL-6 and increases the risk of CAD in South African Indians.
Introduction
Single-nucleotide polymorphisms (SNPs) identified in the promoter regions of the IL-6 gene have been shown to influence mRNA and IL-6 levels in the circulation and are implicated in the development and progression of inflammatory conditions. 5 –7 The IL-6 SNP at position 174 in the promoter region is of particular importance in chronic inflammatory diseases. 8
The IL-6 gene is located at chromosome 7 position p21, 9,10 whereas the IL-6 −174 SNP is situated near a transcription factor “hotspot area” that is closely related to transcriptional regulation of IL-6. 10 Two multiple responsive elements, as well as additional transcription factor binding sites, namely cyclic adenosine monophosphate (cAMP)-responsive elements and CAAT/enhancer binding sites, are contained within the IL-6 gene region between −173 and −145.
Previous studies have found the IL-6 −174 G/C promoter polymorphism at different frequencies in various populations and ethnic groups, including Indians. 11 –13 A recent meta-analysis of studies conducted amongst Caucasian, Turkish, Tunisian, and Chinese populations with CAD showed that the majority of studies documented a high frequency of the IL-6 −174 GC genotype compared to the CC genotype. 14 A study amongst Indians in India showed a high frequency of GG genotypes with higher frequencies of the g allele. 15 Because the data on the association of the IL-6 −174G/C polymorphism with early-onset CAD is limited in South African (SA) Indians, our goal was to investigate the distribution of the −174 G/C IL-6 SNP and IL-6 levels in a SA CAD group.
Materials and Methods
Patient recruitment
One hundred young (mean age 37.5, range 24–45years) SA Indian male CAD patients and 100 age-, race-, and sex-matched controls, 87 age- and sex-matched SA black controls were enrolled in the study following institutional ethical approval (BE030/010, BE154/010). A full pathology report was compiled. The inclusion criteria for CAD patients were Indian ancestry and unrelated, adults below the age of 45 years, and stable CAD confirmed at angiography. The exclusion criteria for controls were an acute coronary syndrome/revascularization procedure in the preceding 3 months, chronic renal or liver disease, malignancy, and known active inflammatory or infectious disease.
DNA isolation
Genomic DNA was extracted from whole blood collected from each study participant. Cells were transferred to 500 μL of lysis buffer [0.5% sodium dodecyl sulfate (SDS), 150 mM NaCl, 10 mM ethylenediaminetetraacetic acid (EDTA), 10 mM Tris-HCl (pH 8.0)]. RNase A (100 μg/mL, DNase-free) was added to this buffer and incubated at 37°C for 1 hr. Subsequently proteinase K (200 μg/mL) was added and incubated for 3 hr at 50°C, and a 0.1% volume of 5 mM potassium acetate was added before centrifugation at 5000×g for 15 min. Supernatants containing genomic DNA were transferred to fresh tubes, extracted with 100% isopropanol, and washed with 70% ethanol. DNA samples were solubilized in 10 mM Tris and 0.1 mM EDTA (pH 7.4, 4°C). Concentration of DNA was determined spectrophotometrically and standardized.
Polymerase chain reaction–restriction fragment length polymorphism
Evaluation of SNPs in the −174 G/C IL-6 (rs1800795) promoter region was done using polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP). The IL-6 promoter region was amplified using 40 pmol of both forward and reverse primers. PCR reaction volume for each sample was 40 μL, comprising GoTaq® Flexi Buffer, 25 mM MgCl2, 200 μM of each deoxyribonucleotide triphosphate (dNTP), 0.5 U of GoTaq® DNA polymerase (Promega), and 10 ng of genomic DNA template. Amplification conditions were initial denaturation at 95°C for 5 min, followed by 35 cycles of denaturation at 95°C for 30 sec, annealing at 55°C for 30 sec, and extension at 72°C for 30 sec. Final extension was achieved at 72°C for 7 min. The following primers were used to amplify the IL-6 promoter region: −174 (5′-TGACTTCAGCTTTACTCTTTG-3′ and 5′-CTGATTGGAAACCTTATTAAG-3′). PCR amplified a 168-bp product. DNA bands were visualized by ultraviolet (UV) light and digitally photographed using a gel documentation system (Uvitech Alliance 2.7). To determine the −174G/C genotype, the PCR product was restricted with the restriction enzyme NlaIII (5 U/μL) (Fermentas). NlaIII cuts in the presence of the −174 C-allele, resulting in two fragments of 119 bp and a smaller fragment that migrates off the gel. No cleavage occurred in the presence of the −174 G-allele (Fig. 1). 12
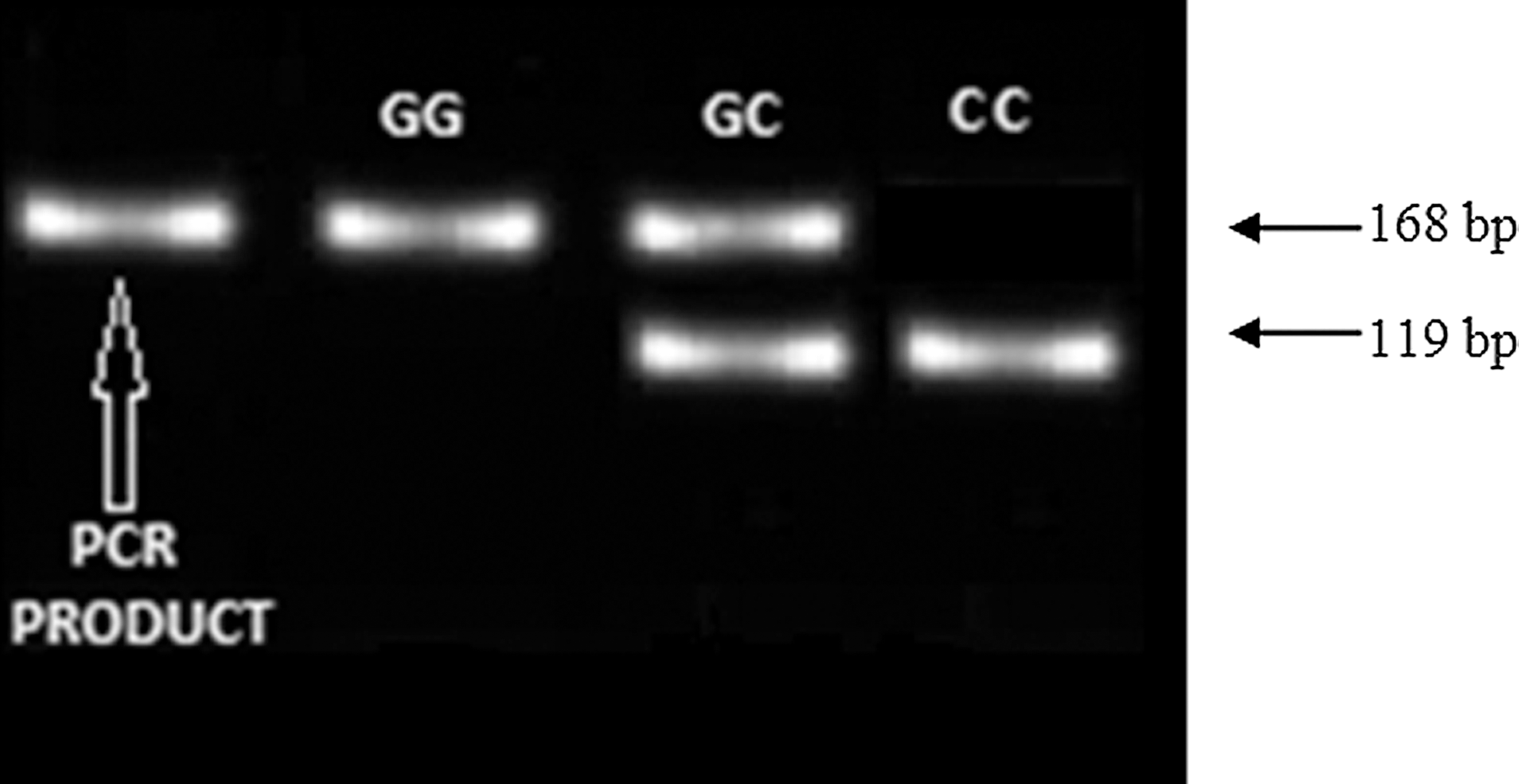
Interleukin-6 (IL-6) −174 band pattern following restriction of the PCR product with NlaIII.
IL-6 enzyme-linked immunosorbent assay
A human IL-6 ELISA Kit II (BD OptEIA™) was used to measure the IL-6 levels of patients and controls. As per the user manual, enzyme-linked immunosorbent assay (ELISA) diluent (12 mL of buffered protein base, 0.09% sodium azide) was added to a 96-well microtiter plate coated with anti-human IL-6 monoclonal antibody, and standards and samples were added into appropriate wells. Following incubation for 2 hr at room temperature, wells were rinsed with wash buffer (1× detergent solution with ProClin™-150). Working detector (containing streptavidin–horseradish peroxide and biotinylated anti-human IL-6 monoclonal antibody) was added into each well and incubated for 1 hr at room temperature in the dark). Wells were washed as previously, seven times, followed by the addition of TMB One-Step Substrate Reagent (3,3′,5,5′-tetramethylbenzidine) and incubated for 30 min at room temperature in the dark. Following incubation, stop solution (1 M phosphoric acid) was added, and absorbance measured at 450 nm (reference, 570 nm) with a microplate reader (Bio-Tek μQuant).
Statistical analysis
Hardy–Weinberg statistics were calculated. Graphpad Prism Software was used for the Fisher exact test to assess the contingency of alleles. The D'Agostino and Pearson normality tested column statistics, followed by the Mann–Whitney test for differences between groups.
Results
A single nucleotide change at position −174 in the IL-6 promoter region was investigated using PCR-RFLP. The GG/GC/CC genotype frequencies for the −174 IL-6 polymorphism conformed with those predicted to Hardy–Weinberg equilibrium (Table 1).
Fisher exact test for heterogeneity between a total SA black and total SA Indian (P<0.0001), b healthy SA black and healthy SA Indian, (P<0.0001), and c healthy SA Indian and coronary artery disease (CAD) patients (P=0.0431) show significant differences in allele frequency where the wild-type G allele occurs most frequently in SA black and CAD patients.
T2D, type 2 diabetes; HWE, Hardy–Weinberg equilibrium.
Interestingly, the healthy SA black group presented with a significantly higher frequency of the GG genotype (96%−GG; 4%−GC; 0%−CC) compared to healthy Indian controls [56%−GG; 31%−GC; 13%−CC; P<0.0001, odds ratio (OR)=0.0507 95% confidence interval (CI) 0.02–0.17].
A high percentage of the Indian control (39%) and CAD patients (59%) were prediabetic/diabetic. Therefore, we assessed genotypic frequencies between these groups but found no significant association with the IL-6 −174 G/C polymorphism (Table 1). The frequency of −174 IL-6 genotypes in the healthy Indian control group was 56% GG, 31% GC, and 13% compared to CAD patients with 71% GG, 27% GC, and 2% CC (Table 1). A significant association with −174 IL-6 G allele and CAD patients was observed compared to healthy Indian controls (P=0.0431, OR=0.4683 95% CI 0.23–0.95) (Table 1).
The levels of IL-6 and CRP between groups with significant outcome for genotypic frequencies (healthy blacks, healthy Indians, and Indians with CAD) were then compared. Levels of IL-6 were only significantly different between ethnic groups (P<0.0001; Table 2). A consistent trend amongst the groups was seen with the homozygous −174 G genotype presenting with the higher level of IL-6 compared to C allele carriers. This elevation was only statistically significant in the healthy SA Indian control group [GG 0.9(0.9, 4.29) vs. GC/CC 0.75 (0.64, 0.9); P=0.0001; Fig. 2].
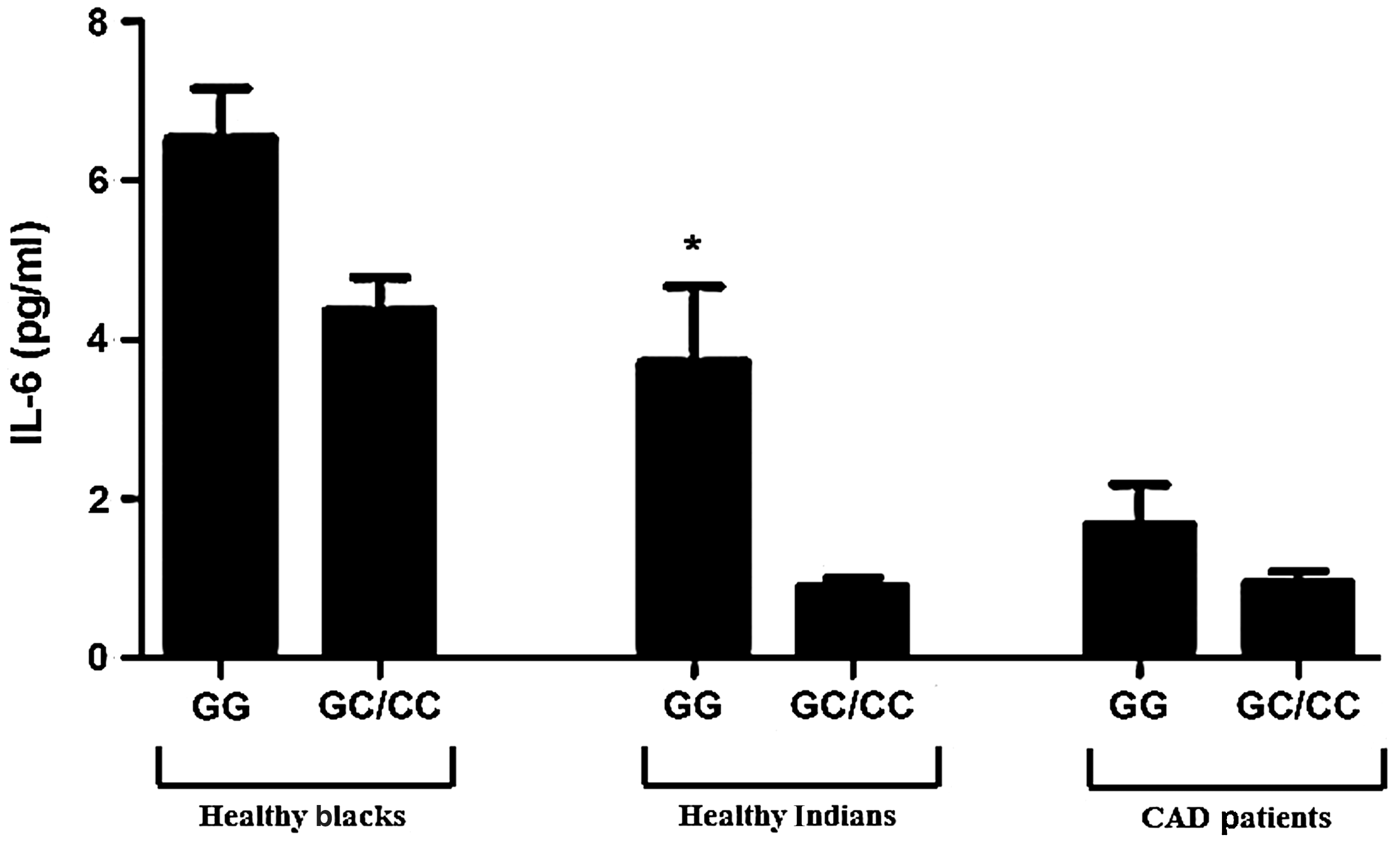
Levels of interleukin-6 (IL-6) stratified by −174 IL-6 genotypes. (*) P=0.0001 where GG is significantly higher than GC/CC in the healthy Indian group (Mann–Whitney test (sum of ranks: 1212, 499.5 and Mann–Whitney U=174.5). Data are represented as mean±standard error of the mean. CAD, coronary artery disease.
Data are represented as the median (25th percentiles, 75th percentiles).
P<0.0001 compared to total blacks.
P<0.0001 compared to healthy blacks.
CRP, C-reactive protein; CAD, coronary artery disease.
Discussion
Modification and entry of low-density lipoprotein (LDL) into the endothelium promotes an inflammatory response to vascular injury. 16 This stimulates the recruitment of inflammatory cells such as monocyte-derived macrophages and lymphocytes and subsequent release of proinflammatory cytokines. Both CRP and IL-6 are implicated in the pathogenesis of atherogenesis and have clinical significance as risk factors for cardiovascular diseases. 9,17
IL-6 and its signaling effects have been shown to contribute to both atherosclerotic plaque formation and eventual destabilization through several mechanisms. 16 These include endothelial activation, smooth muscle cell proliferation, lymphocyte recruitment, and mediation of the acute-phase response. 18 In addition, IL-6 affects expression of scavenger receptors SR-A and CD-36, which are involved in the uptake of modified LDL, and therefore promotes foam cell production. 16
The IL-6 −174 SNP location near the “hotspot vicinity” for transcription factor NF-1 binding acts as a repressor of gene expression. 19 Several population genetic studies revealed conflicting results regarding the influence of the IL-6 −174 SNP on IL-6 production. 20 A study of IL-6 levels in a group of healthy men and women (adjusted for age, body mass index, gender, and smoking) found lowest levels in the presence of the CC genotype 1.63 (95% CI 1.44–1.86) compared to GG 2.74 (95% CI 2.43–3.10) and GC 2.64 (95% CI 2.35–2.97) [P=0.02 by analysis of variance (ANOVA)], 8 whereas another study on 127 patients (following coronary artery bypass graft surgery) showed a recessive effect of the C allele to increase IL-6 levels. 18
In our study, the highest levels of IL-6 were detected in nondiabetic CAD subjects with the homozygous G allele (P=0.0001; Fig. 2). The transcriptional regulation of IL-6 is highly complex, and nearby genetic variants may influence transcription of the IL-6 gene. 17,21 A study in an Italian group of 88 patients assessed the prognostic significance of IL-6 in the diagnosis of acute myocardial infarctions. They found that elevated creatine kinase MB (P<0.05) and IL-6 levels (P<0.01) were independently associated, with a final diagnosis of acute myocardial infarctions. Elevated IL-6 levels also significantly predicted the risk of acute myocardial infarction (AMI) (OR=2.47, P=0.006). 22
The role of SNPs in IL-6 genes in pathogenesis of atherosclerosis still remains unclear. The present study indicates a potential protective effect of the C-allele with respect to CAD. This effect, however, is ethnicity specific. 23
Footnotes
Acknowledgments
A.P. thanks Miss Shivona Gounden for technical assistance and the NRF-DAAD (SA) for a scholarship. We thank UKZN (College of Health Sciences) for funding this study.
Author Disclosure Statement
No competing financial interests exist.