
Abstract
Background:
Statins significantly reduce cardiovascular events in a broad population of patients with hyperlipidemia. However, a small, but significant risk of new-onset diabetes has been reported in patients treated with statins. The mechanism by which statins cause diabetes has not been elucidated and therefore preventive strategies have yet to be defined.
Method:
Our goal was to study the differing effects of a lipophilic (simvastatin) statin, hydrophilic (pravastatin) statin, and ezetimibe on glucose transporter-4 (GLUT4) protein expression in 3T3-L1 adipocytes. We hypothesized that the reductions in GLUT4 protein secondary to statin treatment would be prevented when cells were co-incubated with coenzyme Q10 (CoQ10). GLUT4 protein expression was determined using the In-Cell Western technique. Confluent adipocytes were differentiated using a hormonal cocktail for 3 days; followed by treatment with simvastatin, pravastatin, ezetimibe and CoQ10. Cell morphology was observed after treatment using phase-contrast microscopy.
Results:
Treatment with simvastatin (P<0.001) and simvastatin plus ezetimibe (P<0.001) significantly decreased GLUT4 protein expression in the adipocytes compared to control conditions. GLUT4 protein levels were similar to control after treatment with ezetimibe alone (P=0.52) or pravastatin (P=0.32). There was no significant difference (P=0.098) in GLUT4 protein levels after co-treatment with CoQ10 between any of the treatments and control conditions.
Conclusion:
Our studies have shown that lipophilic statins (simvastatin) reduce the GLUT4 protein levels in adipocytes, whereas hydrophilic statins (pravastatin) or ezetimibe do not. Co-treatment with CoQ10 appears to prevent the reduction in GLUT4 protein levels caused by simvastatin.
Introduction
The addition of mevalonate to cell culture medium has been shown to reverse the effects of lovastatin on GLUT4 expression, 9 suggesting that changes in GLUT4 might be secondary to decrease synthesis of isoprenoid cholesterol precursors. 9 Ubiquinones are electron carriers linking the electron transport chain complexes in mammalian mitochondria and contain isoprenoid residues that are synthesized from mevalonate. Therefore, statins, by reducing the biosynthesis of mevalonate, reduce endogenous production of ubiquinone, including coenzyme Q10 (CoQ10), which is a member of the ubiquinone family. Recently, Muraki et al. demonstrated that treatment of mice with CoQ10 reversed mitochondrial dysfunction and exercise intolerance induced by atorvastatin. 11 Mitochondrial dysfunction has been implicated in causing insulin resistance in the elderly and young lean offspring of parents with T2DM through reduced mitochondrial oxidative phosphorylation activity. 12 Therefore, it is possible that concomitant exposure of adipocytes to CoQ10 may have positive effects on statin-induced reductions in GLUT4 protein expression.
Ezetimibe is often combined with a statin to further reduce LDL-C in patients not achieving their target levels on maximum statin doses or unable to tolerate maximum doses. 13,14 Dagli et al. have shown that a low-dose pravastatin and ezetimibe combination improved insulin resistance and was more effective at reducing LDL-C relative to high-dose pravastatin therapy. 15 Yang et al. have shown that chronic administration of ezetimibe improves glycemic control and pancreatic β-cell mass and increases serum active glucagon-like peptide-1 levels, with possible beneficial effects on glycemic control. 16 The effects of ezetimibe on GLUT4 protein expression have not been described.
We hypothesized that simvastatin and pravastatin would have differential effects on the expression of GLUT4 protein, ezetimibe would have a neutral effect, and that CoQ10 would ameliorate reductions in GLUT4 protein induced by the statins.
Materials and Methods
Chemicals
Dulbecco modified Eagle medium (DMEM) was purchased from Life Technologies (Grand Island, NY). DRAQ5™ was obtained from Cell Signaling Technology (Boston, MA). Dimethylsulfoxide (DMSO), fetal calf serum (FBS), ethanol, isopropyl alcohol, penicillin-streptomycin cocktail, TrypLE™ Express (1×), insulin, dexamethasone, isobutylmethylxanthine (IBMX), and pravastatin sodium salt hydrate were purchased from Sigma-Aldrich (St. Louis, MO). Simvastatin hydroxyl acid, CoQ10, and ezetimibe were obtained from Fischer Scientific (Pittsburgh, PA). Blocking buffer and secondary antibodies were obtained from LI-COR Bioscience (Lincoln, NE), and rabbit monoclonal GLUT4 primary antibody was obtained from Novus biological.
Cell culture
3T3-L1 fibroblasts was generously donated by Dr. Babu Tekwani from the University of Mississippi and originally purchased from the American Type Culture Collection (ATCC; Manassas, VA) was used. The cells (passage 25) were cultured and grown in a 96-well plate in high-glucose DMEM with 10% FBS and penicillin (10,000 IU/mL)/streptomycin (10,000U/mL) at 37°C in a humidified atmosphere, containing 5% CO2. Culture medium was changed three times a week. Four days following confluence, the cells were incubated in DMEM with 10% FCS, 1 μg/mL insulin, 100 μM IBMX, and 0.25 μM dexamethasone (DXM) for 3 days. The cells were further incubated for 3 days in FCS medium with simvastatin (10 μM) alone and combined with ezetimibe (10 μM), pravastatin (10 μM, or FCS medium alone in quadruplet with and without CoQ10 at a concentration of 10 μM. Cell morphology was observed after treatment using phase-contrast microscopy (at 20× magnification).
In-Cell Western analysis
The cells were then fixed with 3.7% formaldehyde in phosphate-buffered saline (PBS) for 20 min at room temperature, followed by three permeabilization washes with 0.1% Triton X-50 in PBS for 5 min on a titer plate shaker. The cells were then rinsed with PBS, and nonspecific binding was blocked with LI-COR blocking buffer for 1.5 hr at room temperature. The cells were incubated overnight at 4°C with primary antibody. The cells were rinsed with PBS and incubated with secondary antibody in blocking buffer for 1 hr at room temperature. The relative intensities of GLUT4 (800-nm channel) were calculated by normalizing to cell number using DRAQ5 fluorescent dye (700-nm channel).
Statistical analysis
Statistical analysis was conducted using SigmaPlot software (v 11.0; Systat Software, Inc., San Jose, CA). One-way analysis of variance (ANOVA) was used to compare the mean relative intensity between treatments with an a priori significance level of P<0.05. All pairwise multiple comparisons were conducted using the Holm–Sidak method.
Results
A significant decrease (P<0.001) in relative intensity of GLUT4 protein by approximately 36% was observed when the 3T3-L1 adipocytes were treated with simvastatin (0.042±0.006) compared to control (0.066×0.003), as shown in Fig. 1. Pravastatin (0.067±0.004) and ezetimibe (0.072±0.009) did not statistically alter the relative intensity of GLUT4 compared to control. The combined treatment with ezetimibe and simvastatin also significantly reduced (P<0.001) the relative intensity GLUT4 protein (0.044±0.0027). As compared to controls, cell morphology appeared to be altered by simvastatin and simvastatin combined with ezetimibe (Fig. 1). Cell morphology appeared to be similar between adipocytes treated with pravastatin and ezetimibe compared to control conditions.
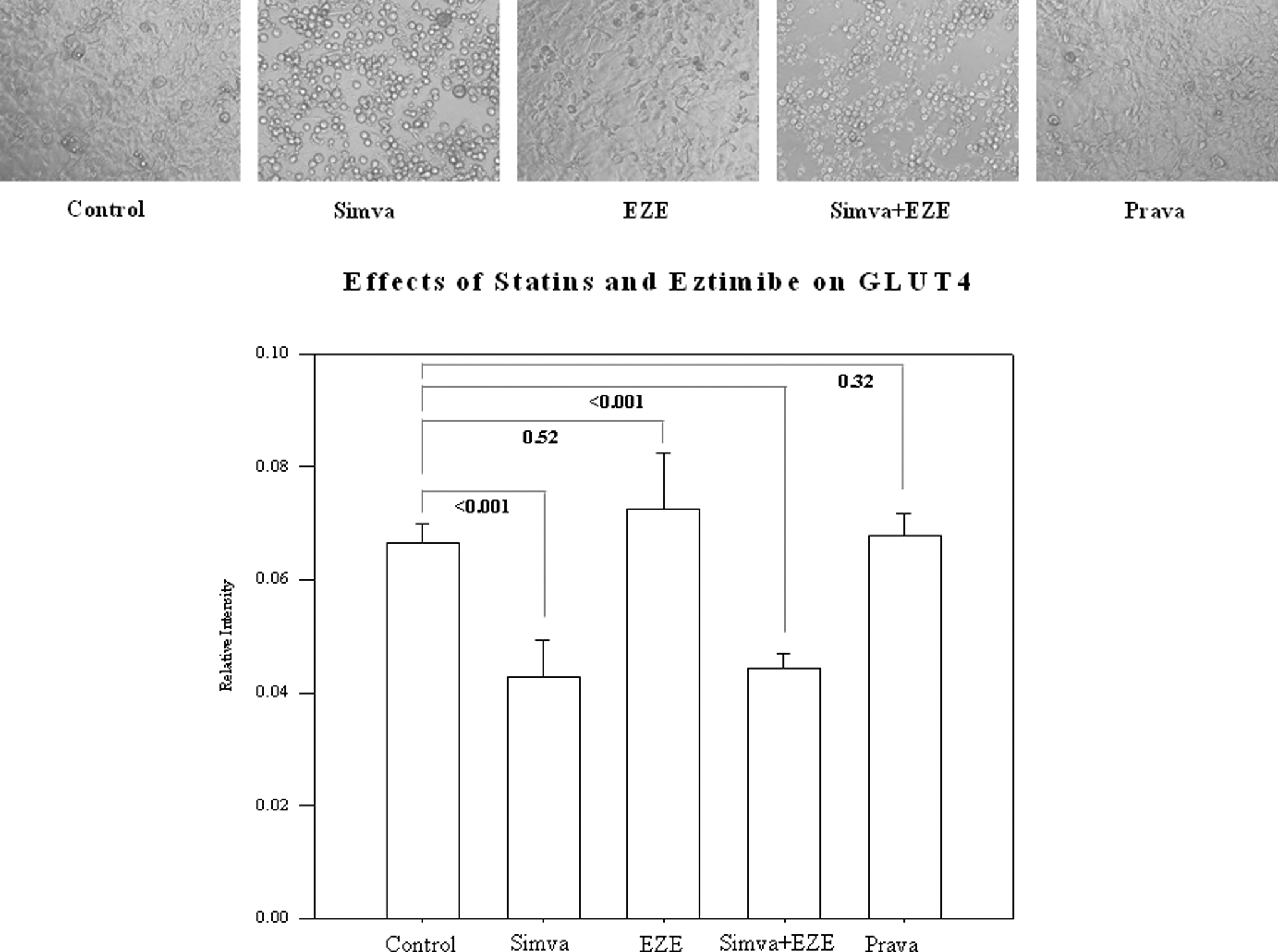
Effects of simvastatin (Simva), ezetimibe (EZE), and pravastatin (Prava) on the relative intensity of glucose transporter-4 (GLUT4) levels in 3T3-L1 adipocytes and cell morphology using phase-contrast microscopy. Magnification, 20×.
The addition of 10 μM CoQ10 to simvastatin alone (0.053±0.007) or combined with ezetimibe (0.061±0.014) in the culture media ameliorated the reduction in relative intensity of GLUT4 protein expression (P=0.098) compared to control (0.065+0.009), as depicted in Fig. 2. Cell differentiation appears to be improved in simvastatin-treated adipocytes after concomitant exposure to CoQ10 (Fig. 2).
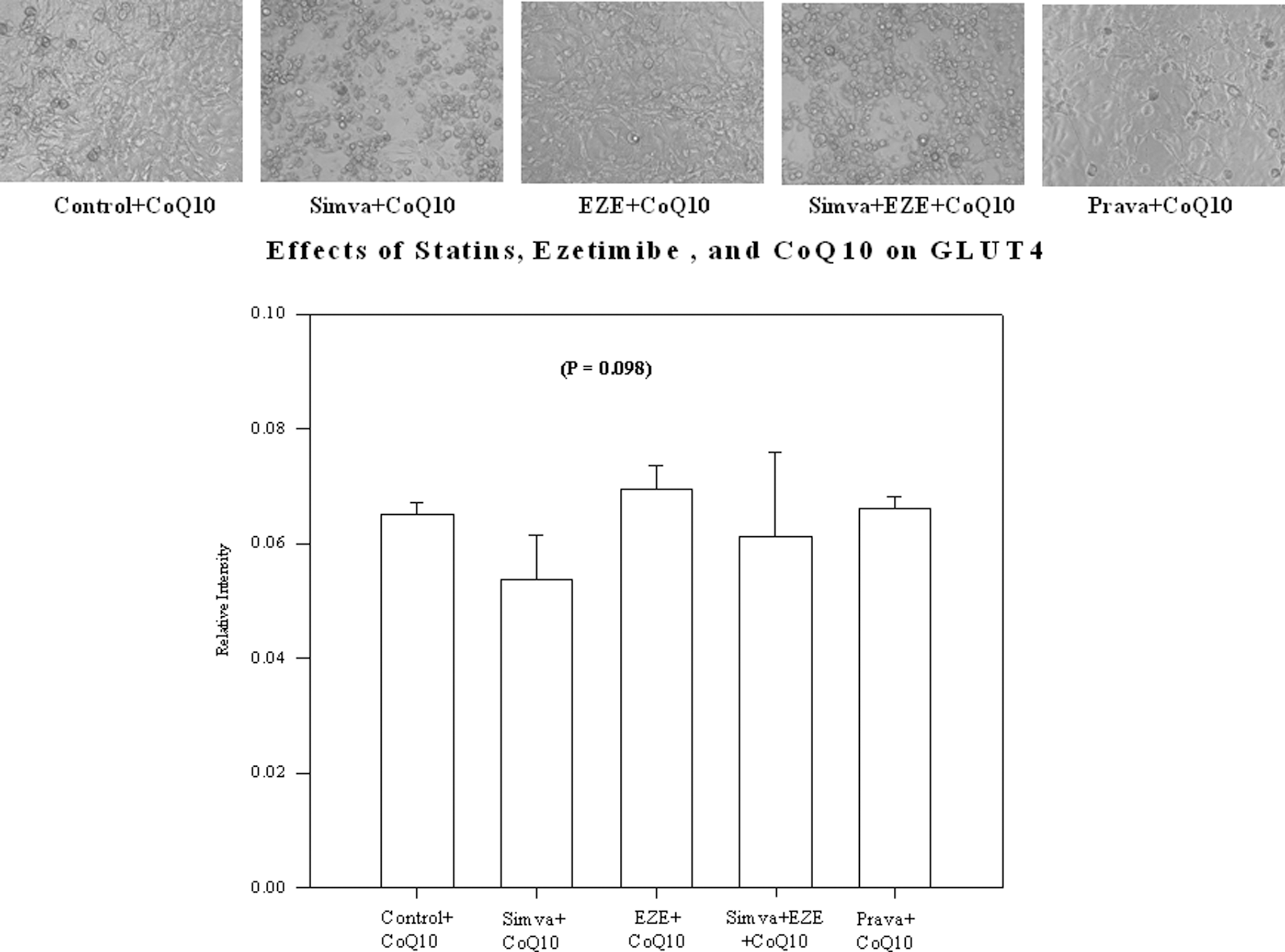
Effects of simvastatin (Simva), ezetimibe (EZE), and pravastatin (Prava) on the relative intensity of glucose transporter-4 (GLUT4) levels in the presence of coenzyme Q10 (CoQ10) in 3T3-L1 adipocytes and cell morphology using phase-contrast microscopy. Magnification, 20×.
Discussion
The insulin resistance and development of diabetes while taking a statin are undesirable side effects and remain a concern in clinical practice. The mechanism underlying the cause of statin-induced diabetes is still poorly understood and therefore preventive strategies are unknown. The results of this current investigation are consistent with other in vitro studies that have shown that lipophilic statins reduce the expression of GLUT4 protein in adipocytes 9,10 and that pravastatin does not appear to adversely affect GLUT4 protein expression. 10 We believe that the difference in lipophilicity between the statins alters their ability to permeate the adipocyte cell membranes and appreciably interact with 3-hydroxy-3-methyl-glutaryl-CoA (HMG-CoA) reductase, which may explain the differences observed in GLUT4 protein expression. The new finding from this study is that CoQ10 ameliorated the decrease in GLUT4 protein levels induced by simvastatin.
This is an interesting observation that provides impetus for additional mechanistic studies for CoQ10 supplementation and the pathogenesis of statin-induced diabetes. CoQ10 has been found to be a key component in mitochondrial bioenergy transfer. 17 Its enzymatic processes facilitate electron transfer in the generation of adenosine triphosphate (ATP). Magnetic resonance spectroscopy studies have shown that insulin is an important regulator of skeletal muscle mitochondrial ATP synthesis and in offspring of parents with T2DM, insulin-stimulated ATP synthesis is significantly reduced compared to healthy subjects. 18 Ubiquinone content in muscle mitochondria has been shown to be significantly reduced with atorvastatin-treated mice compared to mice treated with pravastatin or control conditions. The reduction in ubiquinone levels secondary to atorvastatin treatment was reversed by supplementation with CoQ10 and resulted in improved exercise endurance. 11 We did not measure ubiquinone levels in this study, but this would be one of several important parameters to measure in future studies.
In summary, our study showed that a lipophilic statin, simvastatin, caused impairment in GLUT4 protein expression compared to the pravastatin, a hydrophilic statin. CoQ10 appears to ameliorate the reduction in GLUT4 protein expression caused by simvastatin. Finally, ezetimibe alone does not affect GLUT4 protein expression and did not ameliorate the effect of simvastatin on the transporter.
Footnotes
Author Disclosure Statement
No competing financial interests exist.