
Abstract
Background:
We investigated the possible association of uncoupling protein 3 gene (UCP3) single nucleotide polymorphisms (SNPs) with nonalcoholic steatohepatitis (NASH) and metabolic syndrome (MetS) in nonalcoholic fatty liver disease (NAFLD) Brazilian patients.
Methods:
UCP3 SNPs rs1726745, rs3781907, and rs11235972 were genotyped in 158 biopsy-proven NAFLD Brazilian patients. Statistics was performed with JMP, R, and SHEsis softwares.
Results:
The TT genotype of rs1726745 was associated with less occurrence of MetS (P = 0.006) and with lower body mass index (BMI) in the entire NAFLD sample (P = 0.01) and in the NASH group (P = 0.02). The rs1726745-T was associated with lower values of AST (P = 0.001), ALT (P = 0.0002), triglycerides (P = 0.01), and total cholesterol (P = 0.02) in the entire NAFLD sample. Between groups, there were lower values of aminotransferases strictly in individuals with NASH (AST, P = 0.002; ALT, P = 0.0007) and with MetS (AST, P = 0.002; ALT, P = 0.001). The rs3781907-G was associated with lower GGT elevation values in the entire NAFLD sample (P = 0.002), in the NASH group (P = 0.004), and with MetS group (P = 0.003) and with protection for advanced fibrosis (P = 0.01). The rs11235972-A was associated with lower GGT values in the entire NAFLD sample (P = 0.006) and in the NASH group (P = 0.01) and with MetS group (P = 0.005), with fibrosis absence (P = 0.01) and protection for advanced fibrosis (P = 0.01). The TAA haplotype was protective for NASH (P = 0.002), and TGG haplotype was protective for MetS (P = 0.01).
Conclusion:
UCP3 gene variants were associated with protection against NASH and MetS, in addition to lower values of liver enzymes, lipid profile, BMI and, lesser fibrosis severity in the studied population.
Introduction
Nonalcoholic fatty liver disease (NAFLD) is characterized by the deposition of triglycerides (>5%) in hepatocytes in patients without significant alcohol consumption. 1 The spectrum of the disease encompasses simple steatosis, nonalcoholic steatohepatitis (NASH), and a risk of progression to cirrhosis and hepatocellular carcinoma. 2 NAFLD has shown epidemic behavior, making it the most common chronic liver disease worldwide. 3
The pathophysiology underlying NAFLD is complex and incompletely understood. 4 Despite its high prevalence, only a small minority of affected patients develops inflammation and subsequently fibrosis and chronic liver disease, while most of them only exhibit simple steatosis. 5 The “multiple hit” hypothesis considers several insults acting together on genetically predisposed subjects to induce NAFLD, including insulin resistance (IR), hormones secreted from the adipose tissue, nutritional factors, gut microbiota, and genetic and epigenetic factors. 4 –7
NAFLD is strongly associated with components of metabolic syndrome (MetS), 8 with IR being the critical association condition. 9,10 In the general population, NAFLD is estimated to occur in 6.3–33%, 11 reaching 40–80% of individuals with type 2 diabetes mellitus (T2DM) and 30–90% of obese individuals. 12 In view of the evident association between NAFLD and metabolic dysregulation, a panel of international experts recently proposed a new definition criteria and terminology for the disease as metabolic dysfunction-associated fatty liver disease (MAFLD). 13 However, NAFLD and MAFLD definitions do not identify exactly the same individuals, and therefore, the two terms are not fully interchangeable, 14 especially in lean NAFLD without coexisting metabolic dysregulation, in which the influence of NAFLD-related genetic variants may be decisive. 15
During the last 10–15 years, genetic studies, including genome-wide association studies (GWAS) and candidate gene studies, have provided new data on the importance of single nucleotide polymorphisms (SNPs) in susceptibility and/or progression of NAFLD. 16,17 It is possible that SNPs in genes involved in excessive fatty acid oxidation would predispose NASH. In contrast, these SNPs could determine the inadequate mitochondrial overload during times of excessive free fatty acid (FFA) supply. 18
Uncoupling protein 3 (UCP3) is a mitochondrial anion carrier located on chromosome 11q13, which is selectively expressed in human skeletal muscle. 16,19 UCP3 belongs to a family of mitochondrial transporters known for uncoupling oxidative phosphorylation through proton leakage from the inner mitochondrial membrane and has been implicated in (a) modulation of oxidative status and reduction in reactive oxygen species (ROS) production; (b) body energy homeostasis; (c) regulation of lipid substrates; and (d) protection against IR and T2DM. 20 –22
The possible adaptation of uncoupling proteins to increased fatty acid oxidation could confer a theoretical benefit in the prevention of hepatic steatosis and, through the flow of electrons in the respiratory chain, might prevent the over-reduction of respiratory complexes and excessive mitochondrial ROS formation, protecting cells from lipoperoxidation and diminishing necro-inflammation and apoptosis. 23,24
Several SNPs in the UCP3 gene (UCP3) have been variably associated with obesity-related phenotypes and variations in body mass index (BMI), 25 atherogenic lipid profiles, and a decreased risk of T2DM, 26 with contradictory associations with a higher risk of T2DM 27 and prediabetes. 28 However, only a few studies have evaluated the association of UCP3 SNPs with NAFLD.
A Chinese study 29 described a higher prevalence of the GG genotype of UCP3 SNP rs11235972 in the NAFLD Chinese children group, and a Spanish study 30 reported an association between the CT genotype of UCP3 SNP -55 C/T (rs1800849) with higher frequency of moderate-intense steatosis and inflammation on liver biopsy in NAFLD individuals. Therefore, the present study aimed to evaluate the possible association of UCP3 SNPs rs1726745, rs3781907, and rs11235972 with NASH and MetS in NAFLD Brazilian individuals.
Methods
Clinical design and patient's selection
This was a cross-sectional study in NAFLD individuals obtained from a database of the Hepatology Outpatient Clinic at the Hospital das Clínicas da Faculdade de Medicina da Universidade de São Paulo (HC-FMUSP), São Paulo, Brazil. The material for the SNP analysis was obtained from a DNA sample database stored in the Laboratório de Gastroenterologia Clínica e Experimental (LIM-07) do Departamento de Gastroenterologia e Hepatologia do HC-FMUSP, from individuals who were followed at the Hepatology Outpatient Clinic and who agreed—freely and spontaneously—that their DNA could be stored for future research. The study was approved by the Ethics Committee (2.160.492) of the HC-FMUSP and was conducted in compliance with the Helsinki Declaration.
The diagnosis of NAFLD was made by liver biopsy, which was performed only when the individuals had clinical and laboratory indications. The inclusion criteria were age between 18 and 75 years and biopsy-proven NAFLD. The exclusion criteria were the presence of any other chronic liver disease (viral hepatitis, autoimmune hepatitis, hemochromatosis, alpha-1-antitrypsin, Wilson's disease, celiac disease, schistosomiasis), significant alcohol intake (>100 g ethanol/week), and/or previous use of steatogenic medication (corticosteroids, amiodarone, methotrexate, tamoxifen).
A total of 258 patients were initially evaluated. In the absence of a DNA sample (n = 58) and/or absence of histological criteria for NAFLD (n = 32), a sample of 158 patients was obtained. After UCP3 polymorphism genotyping, patients were evaluated according to the presence of nonalcoholic fatty liver (NAFL) or NASH and according to the absence or presence of MetS.
Clinical variables and biochemical analysis
On the day of the liver biopsy or within a period of up to 3 months thereafter, demographic (age, sex), anthropometric (weight, height, BMI), clinical and laboratory data were obtained. The criteria for MetS were according to the National Cholesterol Education Program/Adult Treatment Panel III (NCEP ATP III). 31
The biochemical tests were performed after 12 hrs of fasting: aspartate aminotransferase (AST), alanine aminotransferase (ALT), gamma glutamyl transferase (GGT), FBG, plasma insulin, total cholesterol and fractions, triglycerides, serum copper, ceruloplasmin, iron profile, hepatic antibodies (antinuclear antibody, anti-smooth muscle antibody), alpha-1-antitrypsin, celiac disease antibodies, and serologies for viral hepatitis B and C. For the evaluation of IR, Homeostatic Model Assessment of Insulin Resistance (HOMA-IR) (fasting glucose (mg/dL)/18 × fasting insulin (μU/mL)/22.5) was used. A HOMA-IR value greater than or equal to 2.5 was used as a marker of IR. 31
DNA extraction and UCP3 SNP genotyping
Genomic DNA was extracted from the blood of each individual by the salting-out method, and the quantification of the DNA was determined by spectrophotometry (NanoDrop 1000 Spectrophotometer; Thermo Fisher Scientific, Waltham, MA). After extraction, the DNA was stored at −20°C.
Three UCP3 SNPs rs1726745, rs3781907, and rs11235972 were genotyped by real-time polymerase chain reaction using the TaqMan Genotyping Master Mix and TaqMan SNP Genotyping Assay (Applied Biosystems, Foster City, CA). The Assay identifications were: C__8751342_10 (rs1726745), C__25800726_10 (rs3781907), and C__25992010_10 (rs11235972).
Histological analysis
Fragments of hepatic tissue previously fixed in 4% formalin solution were processed and stained with hematoxylin–eosin, Masson's trichrome, and Picrosirius. Liver samples were blindly evaluated by an experienced liver pathologist. The histological criteria were evaluated according to the histological scoring system for NAFLD and NAFLD Activity Score (NAS), defined by the NASH Clinical Research Network Pathology Committee (CRN). 32
Patients with simple steatosis were categorized as NAFL individuals. Fibrosis was evaluated accordingly to absence vs presence of fibrosis, significative fibrosis (F ≥ 2), and advanced fibrosis (F3–F4).
Statistical analysis
Results were expressed as mean ± SD except where stated otherwise. Continuous variables were log-transformed for the analyses when the normality of the distribution was rejected by the Shapiro–Wilk W test. Fisher's chi-squared test, Student's t test (or Mann–Whitney test), and ANCOVA were used for comparisons between groups. The Hardy–Weinberg Equilibrium (HWE) test was performed in all polymorphisms. Associations with NASH, MetS, and fibrosis were assessed by regression models. Logistic regression analyses were used for cross-sectional analyses.
Odds ratios (OR) with its 95% confidence intervals (CI) were computed for the minor allele of each SNP in a codominant, dominant, or recessive model. A value of P ≤ 0.02 was considered as significant for genotype-related comparisons. Statistical analyses were performed using JMP (SAS Institute, Inc., Cary, NC), R (
Results
Characteristics of the study population
The demographic, biochemical, and clinical characteristics of 158 NAFLD patients are listed in Table 1. The mean age was similar between the NAFL and NASH groups, as well as the distribution between genders, with female sex being most prevalent in both groups. NASH patients showed a higher frequency of MetS (P = 0.02), higher BMI (P = 0.02), and higher concentrations of AST (P = 0.002).
Demographic, Biochemical, and Clinical Characteristics of Patients with Nonalcoholic Steatohepatitis/Nonalcoholic Fatty Liver
The results are expressed as the mean ± SD.
P ≤ 0.05.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; BMI, body mass index; GGT, gamma glutamyl transferase; HDL, high-density lipoprotein; HOMA-IR, homeostatic model of assessment of insulin resistance; LDL, low-density lipoprotein; NAFL, nonalcoholic fatty liver; NASH, nonalcoholic steatohepatitis.
The overall prevalence of MetS was 82.6% (n = 128), predominating in the NASH group (85% vs. 63%). The prevalence of NASH was higher in patients with MetS (90.6% vs. 74%).
Single site associations
The genotype frequency distributions of the three gene variants were consistent with the HWE (Table 2). The TT genotype of rs1726745 was associated with less occurrence of MetS (OR = 0.18; 95% CI = 0.05–0.61; P = 0.006) (Table 3). No association was found between UCP3 SNP genotypes and NAFL or NASH groups.
Genotypic and Allelic Distributions of UCP3 Polymorphisms
genotyping was not obtained in two individuals.
Genotype representation: major/minor alleles.
HWE, Hardy–Weinberg Equilibrium; SNPs, single nucleotide polymorphisms; UCP3, uncoupling protein 3 gene.
Genotype Frequencies of UCP3 Polymorphisms According to the Presence of Metabolic Syndrome
OR (odds ratio) for the minor allele in a recessive model obtained in logistic regression analysis adjusted for sex, age, type 2 diabetes mellitus, and dyslipidemia. *P ≤ 0.02.
OR [2428148 (9.9 × 1015 – -)]: due to the AA genotype absence among Without MetS individuals, it was not possible to calculate precisely the OR.
MAF, minor allele frequency; MetS, metabolic syndrome.
According to the clinical and biochemical variables
For rs1726745, T-allele carriers were found to have significantly lower values of AST, ALT, triglycerides, and total cholesterol (Table 4). In comparison between groups, there were lower aminotransferase values strictly in NASH (vs. NAFL) and MetS (vs. without MetS) subjects and lower triglyceride values in NASH subjects (vs. NAFL). In homozygosis, the T-allele carriers had lower BMI in the general sample (CX = 31.53 ± 0.56 vs. TT = 29.21 ± 0.84 kg/m2; mean ± standard error of the mean, SEM; P = 0.01) and in NASH individuals (NASH group: CX:31.86 ± 0.61 vs. TT 29.65 ± 0.95; mean ± SEM; P = 0.02) versus NAFL group (CX = 28.52 ± 1.56 vs. TT 27.21 ± 2.05; mean ± SEM; P = 0.65).
Associations Between Uncoupling Protein 3 Gene Single Nucleotide Polymorphisms with Biochemical Variables
The results are expressed as the mean ± standard error of the mean (SEM), with comparisons made by ANCOVA, adjusted for sex, age, body mass index, and type 2 diabetes.
P ≤ 0.02.
For rs3781907 (G-allele) and rs11235972 (A-allele), there was an association with lower GGT elevation values, especially in NASH (vs. NAFL) and MetS (vs. without MetS) individuals (Table 4).
According to the histological variables
The presence of A allele of rs11235972 was associated with fibrosis absence (F0 vs F1–F4; OR = 0.34; 95% CI = 0.14–0.80; P = 0.01; Table 5) and protection against advanced fibrosis (F0–F2 vs. F3–F4; OR = 0.17; 95% CI = 0.03–0.56; P = 0.01). The G allele of rs3781907 also indicated to be protective against advanced fibrosis (F0–F2 vs. F3–F4; OR = 0.25; 95% CI = 0.08–0.69; P = 0.01). No association was found between rs1726745, rs3781907, and rs11235972 genotypes and the histological criteria for NASH (steatosis, lobular inflammation, ballooning), NAFLD Activity Score, and significative fibrosis (F ≥ 2) (Supplementary Tables S1–S3).
Genotype and Allele Frequencies of UCP3 Polymorphisms According to Presence of Fibrosis
OR (odds ratio) for the minor allele in a dominant model obtained in logistic regression analyses adjusted for sex, age, dyslipidemia, metabolic syndrome, and HOMA.
P ≤ 0.02.
Haplotype analyses
Pairwise measures of LD between the analyzed loci were quantified by the correlation coefficient r2 (Fig. 1). According to the presence of NASH, TAA (rs1726745, rs3781907, rs11235972) haplotype presented a protective effect against NASH (OR = 0.01; 95% CI = 0.00–0.12; P = 0.002; Table 6) and, according to the presence of MetS, TGG haplotype exerted a protective effect against MetS (OR = 0.22; 95% CI = 0.07–0.69; P = 0.01). None of the studied haplotypes were associated with fibrosis.
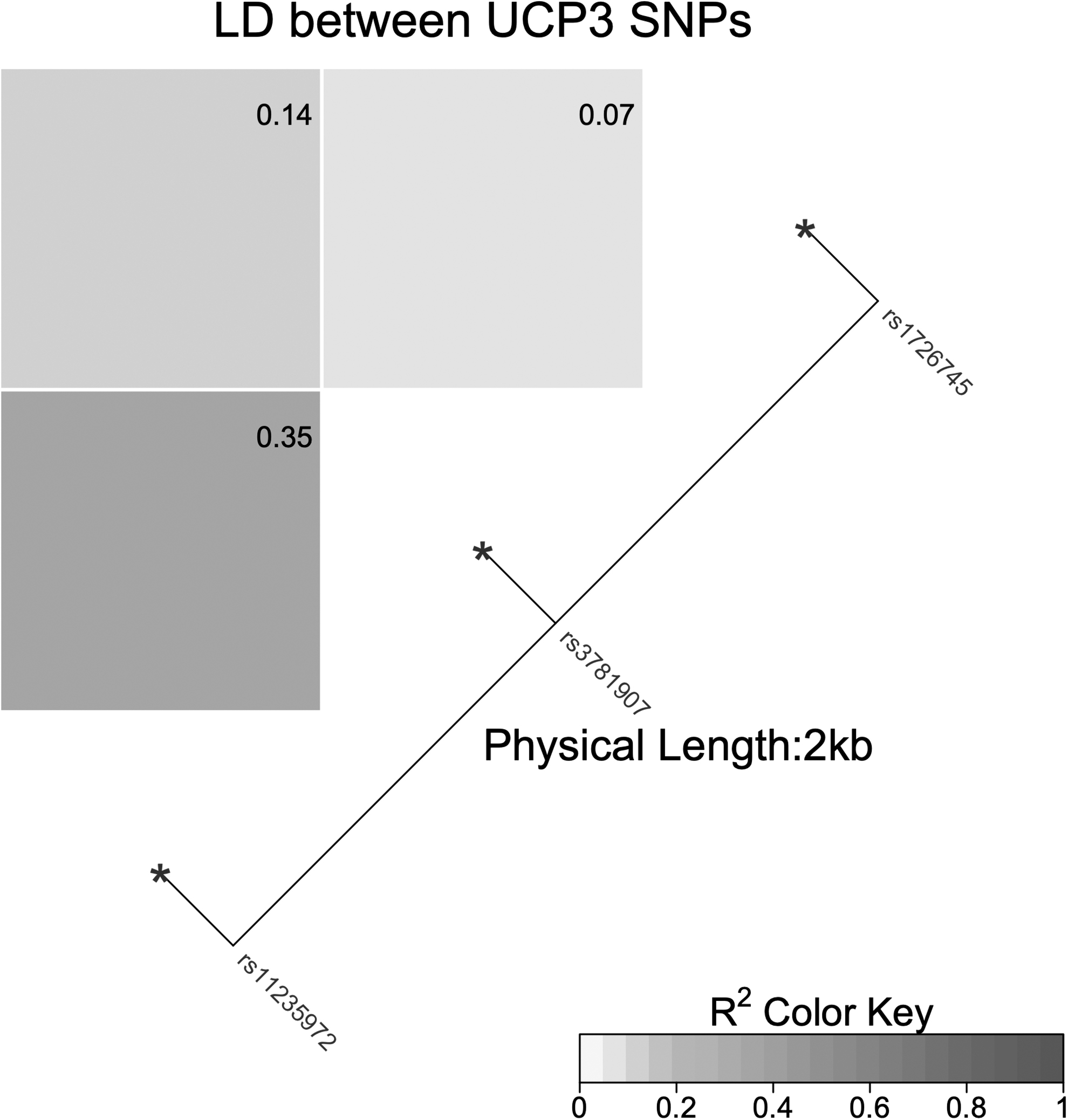
Linkage disequilibrium (r 2 metric) measurements for the UCP3 SNPs. UCP3, uncoupling protein 3 gene; SNP, single nucleotide polymorphism.
Haplotype Analysis in UCP3 Polymorphisms According to Presence of Nonalcoholic Steatohepatitis
Haplotypes represent the alleles of rs1726745, rs3781907, and rs11235972, respectively. Odds ratios (OR) and 95% CI for NASH for each haplotype. P value adjusted by FDR using Benjamini–Hochberg.
*P ≤ 0.02.
Discussion
In the present study, we investigated the possible association of UCP3 polymorphisms with NASH and Mets in adult NAFLD individuals. To our knowledge, this was the first study to evaluate this association in the Brazilian population, with the relevant differential of histological diagnosis of NAFLD.
Currently, a global prevalence of NAFLD (diagnosed by imaging) is estimated at 25.24% and a global prevalence of NASH (histological diagnosis) of 59.1%, ranging from 60.64% to 69.25% in patients with NAFLD with biopsy indication (elevation of liver enzymes, clinical signs of liver disease or retrospective biopsy evaluations of tertiary care center) and reduced prevalence (6.67%) in patients without biopsy indication. 33 Consistent with the profile of NAFLD patients from a tertiary center, we identified a high occurrence of NASH (87.3%) in our sample.
A strong association between NAFLD and MetS components was observed in our study, reinforcing a possible bidirectional association with reciprocal causality between the two conditions. 10,34 About 44% of genes associated with NAFLD are shared by other MetS components (almost 14% with T2DM, obesity, and hypertension combined); however, NAFLD and MetS depend not only on genetic factors but also on environmental factors and the interaction between the two. 35
In the individual evaluation of the rs1726745, rs3781907, and rs11235972 UCP3 polymorphisms according to presence of NAFL/NASH, no significant association was identified. However, in the haplotype analysis, the TAA (rs1726745, rs3781907, rs11235972) haplotype presented as protective for NASH.
SNPs (biallelic) sometimes capture few information about genetic variability in certain regions of the genome. 36,37 Haplotypes (multiallelic) have a greater possibility of detecting phenotypic changes in relation to isolated SNPs, providing a more informative and effective analysis. 36,37 This result may also be related to the NAFLD multifactorial pathogenesis and its polygenic behavior, 38,39 whereas the genetic component of the disease is not as simple as the Mendelian pattern of unique genetic inheritance. 7 A single Chinese study analyzed the NAFLD risk in pediatric patients with UCP3 haplotypes (rs1800849, rs11235972, rs11235972, rs3781907, rs1726745); however, no significant association was identified. 29
Regarding MetS, our study identified the TT genotype of rs1726745 (individual analysis) and the TGG (rs1726745, rs3781907, rs11235972) haplotype as protectors for MetS. These results are in agreement with Groop, 40 who considered, among genes contributing to MetS and regulation of lipolysis, the UCP3 as a particularly attractive candidate gene. Conversely, Meirhaeghe et al. 41 and de Luis et al. 42 did not identify a significant association between UCP3 SNP rs1800849 and MetS as an entity. We consider that a possible reduction in the MetS overall risk through the individual interaction of the UCP3 SNPs with MetS components must be taken into account. 43
Several UCP3 polymorphisms have been variably associated with obesity-related phenotypes 42 and changes in BMI. 43 The TT genotype of UCP3 SNP -55 C/T (rs1800849) was associated with higher BMI in Asians. 44 A significant association between homozygotes for the lowest allele of UCP3 SNPs rs647126 (allele A), rs1685356 (allele A), and rs2075577 (allele G) and higher BMI in Dutch was reported. 45 Conversely, a Japanese study described an association between the TT genotype of UCP3 SNP -55 C/T with significantly lower BMI and considered that, in homozygous form, the SNP could contribute to the reduction in the prevalence of obesity by increasing UCP3 mRNA levels and fat oxidation, thus reducing BMI. 46
In the present study, the TT genotype of rs1726745 correlated with lower BMI values in the general sample and in the NASH group (vs. NAFL), supporting that UCP3 seems to be involved in obesity, given its possible participation in energy regulation and lipid metabolism. 21,47,48 The result variability between UCP3 SNPs and BMI changes seems to be importantly related to ethnicity, due to differences in lifestyle and body weight distribution, as well as differences in SNP genotype frequencies among ethnicities. 44,45
Another interesting finding in our study was the association of the T allele of rs1726745 with lower values of total cholesterol and triglycerides in the general sample and lower values of triglycerides in the NASH group, corroborating the possible role of UCP3 in lipid metabolism. 49,50 The association of UCP3 SNPs to a better lipid profile, through higher HDL values 46 and/or lower cholesterol and LDL values, 42 has been described in literature. It is conjectured that the risk allele presence of UCP3 SNP could produce a more beneficial lipid state in skeletal muscle and adipose tissue, by modifying the fatty acid consumption rate. 42
Controversially, a Finland study described an association of UCP3 SNPs rs3781907 (allele G), rs1726745 (allele G), rs11235972 (allele A), and rs1800849 (allele T) with higher values of total cholesterol and LDL. 27 The disagreement results between UCP3 SNPs and lipid profile may be related to variations in allelic frequencies and penetrance in different populations, particularly in different ethnic groups. 44
According to Sanyal et al., 51 the liver enzyme elevation (AST, ALT, and GGT) is a liver damage sign and can be considered as useful measure in NAFLD assessment, but their values do not adequately correlate with histological severity. 52 Our study identified a significant association of the T allele of rs1726745 with lower values of AST and ALT in the general sample and in the NASH (vs. NAFL) and MetS (vs. without MetS) groups, without a correlation with histological variables.
Hypothetically, the T allele (rs1726745) correlation with a better metabolic profile (cholesterol total, triglycerides, and BMI lower values and protection against MetS), especially in NASH patients, could contribute to FFAs/adipokine supply reduction, thereby protecting against lipotoxicity, mitochondrial dysfunction, ROS production, inflammation, and cell death. 50,53 However, the interpretation of aminotransferase changes in UCP3 SNPs and NAFLD remains uncertain.
Another liver enzyme frequently elevated in NAFLD is GGT 54 and it has been independently associated with several other pathological conditions, including T2DM, MetS, and CVD. 55 GGT is involved in glutathione metabolism, playing a critical role in the antioxidant defense, detoxification, and inflammation. 56 In oxidative stress, serum levels of oxidized glutathione increase, inducing hepatic GGT, which enables the oxidized glutathione conversion into reduced glutathione. 57 Thus, GGT is considered to play an important role in the antioxidant defense system at the cellular level, being a valuable marker of oxidative stress in NAFLD. 57,58
Some studies suggest that GGT is a significant predictor of NAFLD, 59 –61 and more specifically, that high levels of GGT are associated with a more severe NAFLD histological spectrum (presence of NASH and fibrosis), 62 –64 and that the GGT level reduction could predict histological improvement in NAFLD patients after bariatric surgery. 65 Supporting this correlation, a Turkey study demonstrated that increased GGT levels (cut off of 96.5 U/L, with 83% sensitivity and 69% specificity) was a risk factor for advanced fibrosis in NAFLD (OR = 1.0, 95% CI = 0.98–1.01; P = 0.032). 64
In the present study, both groups (NASH and NAFL) had GGT elevations. Interestingly, in a subanalysis, our study identified an individual risk allele association of rs3781907 (allele G) and rs11235972 (allele A) with lower levels of GGT elevation in the entire NAFLD sample and in the NASH (vs. NAFL) and MetS (vs. without MetS) groups. In addition, the rs3781907 (allele G) was associated with protection for advanced fibrosis, and rs11235972 (allele A) was associated with fibrosis absence and protection for advanced fibrosis.
Thus, it was possible to observe a correlation between lower levels of GGT elevation with less histological severity (fibrosis) associated with the risk allele presence of UCP3 SNPs rs3781907 and rs11235972, possibly related to a potential influence of UCP3 SNPs in reducing ROS production and oxidative stress, and, consequently, apoptosis, fibrogenesis, and GGT levels. In NAFLD, oxidative stress, worsening of the inflammatory state, and endothelial dysfunction are factors implicated in CVD pathogenesis 66 ; however, it is rash to infer a relationship between the lowest GGT levels and cardiovascular risk in our sample.
The UCP3 SNPs protective effect in our sample is highlighted by the interesting statistical significance found strictly in the NASH and with MetS groups for certain variables (AST, ALT, GGT, triglycerides, and BMI), inferring that the minor allele presence promoted a greater protection in groups that, usually, would present worse levels of liver enzymes, lipid profile, and BMI.
Our results must be interpreted within the limitations of cross-sectional studies and the heterogeneity between studies regarding allele frequencies and penetrance in different ethnicities, in addition to variations in age, lifestyle behaviors, medication usage, diet, and sample sizes. 44 The NAFLD histological diagnosis as an inclusion criteria enabled a detailed analysis of the histological variables in relation to the UCP3 SNPs, in the meantime, restricted the sample size. The absence of a control group was another limitation of the present study.
In conclusion, our study suggests that UCP3 polymorphisms rs1726745, rs3781907, and rs11235972 are associated with protection against NASH and MetS, in addition to lower values of liver enzymes, lipid profile, BMI, and lesser fibrosis severity in NAFLD Brazilian patients. Additional studies are necessary to further validate these findings in larger samples and in different populations.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.