
Abstract
DNAzymes of the 10-23 family represent an important class of antisense molecules with implications for therapeutic treatment of diseases. These molecules are single-stranded oligodeoxynucleotides combining the high specificity of oligonucleotide base pairing with an inherent RNA-cleaving enzymatic activity. However, like other oligonucleotide-based molecules these substances might exert so-called off-target effects, which have not been investigated so far for this molecule class. Therefore, the present study investigates putative off-target effects of DNAzymes on innate immune mechanisms using GATA-3-specific DNAzymes that have recently been developed as novel therapeutic approach for the treatment of allergic diseases including allergic asthma. The conserved catalytic domain of 10-23 DNAzymes contains a CpG motif that may stimulate innate immune cells via Toll-like receptor 9 (TLR-9). Therefore, potential TLR-9-mediated as well as TLR-9 independent cell activation was investigated using TLR-9-transfected HEK293 cells, macrophage cell lines and primary innate immune cells. Furthermore, putative effects of GATA-3-specific DNAzymes on the activation of neutrophil granulocytes and degranulation of mast cells/basophils were analyzed. In summary, no innate immune cell-stimulating activities of the tested DNAzymes were observed in any of the systems. Consequently, use of GATA-3-specific DNAzymes may represent a novel and highly specific approach for the treatment of allergic diseases.
Introduction
DNAzymes are catalytically active DNA molecules described for the first time in 1994 (Breaker and Joyce, 1994; Breaker and Joyce, 1995). DNAzymes of the 10-23 family comprise a novel class of single-stranded oligodeoxynucleotide antisense molecules that combine the high specificity of antisense molecules with an inherent RNA-cleaving catalytic activity (Fig. 1A). They consist of a central sequence-conserved catalytic region surrounded by 2 binding arms that are sequence-specific to the respective target mRNA (Santoro and Joyce, 1997). RNA-cleaving DNAzymes have been used in a variety of in vitro and in vivo experimental systems, for example, to interfere with targets playing a role in virus infections (viral RNAs) (Baum and Silverman, 2008), vascular permeability and inflammation (c-Jun) (Fahmy et al., 2006), or experimental glomerulonephritis (TGF-β) (Isaka et al., 2004).
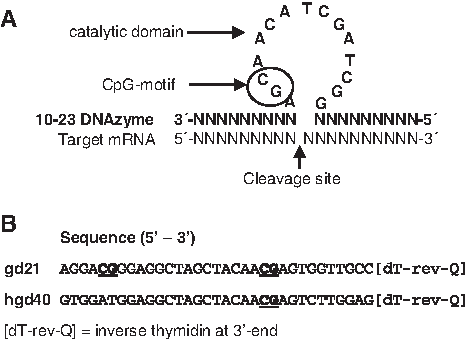
Structure of 10-23 DNAzymes.
In order to therapeutically interfere with the development of allergic diseases we developed mouse and human GATA-3-specific DNAzymes. GATA-3 is the master transcription factor for differentiation and activation of T helper 2 (Th2) cells that are crucial for the development of allergic immune reactions (Zhu et al., 2006).
Specific activities of the GATA-3 DNAzymes gd21 and hgd40 have recently been shown in vitro, for example, by using a mRNA cleavage assay system. Additionally, in vivo preclinical efficacy of these DNAzymes was demonstrated in different experimental mouse models of acute and chronic allergic asthma, including preventive as well as therapeutic approaches. It was shown that intranasal application of these DNAzymes at an optimal dose of 200 μg per application significantly reduced typical features of experimental allergic bronchial asthma pathology (Sel et al., 2008).
Despite the high target specificity of antisense molecules, putative unwanted side effects have to be considered and recent studies actually demonstrated unspecific activities of certain antisense oligonucleotides (Robbins et al., 2008; Robbins et al., 2009). Foreign nucleic acids have been shown to be recognized by pattern recognition receptors (PRRs)—among them Toll-like receptors (TLRs)—and consequently stimulate cells particularly of the innate immune system. In fact, TLR-3 recognizes double-stranded (ds)RNA, TLR-7 and -8 detect single-stranded (ss)RNA and TLR-9 is activated by unmethylated CpG-motifs in ssDNA (Wagner and Bauer, 2006). These receptors and their signaling cascades have evolutionary developed to detect and fight invading microbes such as viruses and bacteria, but may also recognize synthetic antisense oligonucleotides, due to their structural similarities with bacterial or viral nucleic acids. An example of this mode of action is the report about the therapeutic activity of a vascular endothelial growth factor-A-specific siRNA in the treatment of blinding choroidal neovascularization (Reich et al., 2003), which was later solely attributed to the unspecific activation of TLR-3 and subsequent induction of interferon-γ and interleukin-12 (IL-12) (Kleinman et al., 2008). This example underlines the importance of early investigations of potential off-target effects in the development of novel therapeutic antisense molecules.
The objective of the present study was to investigate potential off-target effects of GATA-3-specific DNAzymes that are intended for the local application into the lung for the treatment of allergic airway inflammation. Thus, these molecules will initially come in contact with first-line defense cells of the innate immune system [macrophages, monocytes, different dendritic cell (DC) subsets] and effector cells of allergic immune responses (e.g., granulocytes, mast cells, basophils) in airways and lung, even though it is expected that local DNAzyme concentrations are low due to the fast distribution over the bronchial and alveolar surface. Therefore, a panel of in vitro and in vivo experimental systems was applied to investigate putative unspecific effects on these cells with specific focus on TLR-9-dependent mechanisms since all DNAzymes of the 10-23 family contain an unmethylated CpG-motif in their catalytic domain (see Fig. 1A).
Materials and Methods
Oligonucleotides and control stimulants
GATA-3-specific DNAzymes gd21 and hgd40 and the control DNAzyme ODNg, which contains the central catalytic domain attached to scrambled binding sequences, were developed in our laboratory and have been described elsewhere (Sel et al., 2008). Gd21 was synthesized by Operon (Cologne, Germany), hgd40 and ODNg by Biospring (Frankfurt, Germany). All DNAzymes were modified by an inverted 3′-3′-thymidine. As positive controls for murine and human TLR-9 activation the CpG motif-containing oligonucleotides 1668 CpG and 2216 CpG were applied. All non-DNAzyme oligonucleotides completely consisted of phosphothioate bases (obtained from Operon). The sequences are: gd21: AGGACGGGAGGCTAGCTACAACGAGTGGTTGCC[dT-rev-Q]; hgd40: GTGGATGGAGGCTAGCTACAACGAGTCTTGGAG[dT-rev-Q]; ODNg: TGTGATGTAGGCTAGCTACAACGACAGATGGAA[dT-rev-Q]; 1668 CpG: T*C*C*A*T*G*A*C*G*T*T*C*C*T*G*A*T*G*C*T*; and 2116 CpG: G*G*GGGACGATCGTCG*G*G*G*G*G (*=phosphothioate/[dT-rev-Q]=inverse thymidine at 3′ end).
Oligonucleotides were dissolved at 4 mg/mL stock solution in Dulbecco's phosphate buffered saline (PBS) without Mg2+ and Ca2+ (PAA Laboratories, Cölbe, Germany). Further dilutions were done in respective culture medium. In addition to oligonucleotides, the following stimulants were used as positive controls: lipopolysaccharide (LPS), ionomycin, and zymosan A particles (all from Sigma, Taufkirchen, Germany).
Primary cell isolation and stimulation
Murine mononuclear spleen cells
Balb/c mice (Harlan Winkelmann, Borchen, Germany) were sacrificed, their spleens were removed, and spleen cell suspensions were prepared using 100 μm nylon meshes (BD Falcon, Bedford). Cells were resuspended in RPMI 1640 medium, supplemented with 10% fetal calf serum (FCS), 1%
Human plasmacytoid dendritic cells
Human plasmacytoid dendritic cells (pDC) were isolated from buffy coats of 3 different healthy donors. First, peripheral blood mononuclear cells were isolated by density gradient centrifugation with PanColl human (PAN Biotech) according to manufacturer's instructions. Remaining erythrocytes were lysed with double-distilled water and T lymphocytes were removed with CD3 Micro Beads (magnetic cell sorting of human leukocytes; Miltenyi Biotech, Bergisch Gladbach, Germany) according to manufacturer's instructions. Thereafter, pDC were isolated from the CD3-negative cell pool by negative isolation using the Plasmacytoid Dendritic Cell Isolation Kit (Miltenyi). pDC purity was monitored by flow cytometry following pDC staining with anti-CD303/anti-CD123 (Miltenyi). Mean purity of double-positive cells (pDC) was ≥85%. Cells were re-suspended in RPMI 1640 medium, supplemented with 10% FCS, 1%
Stimulation of macrophages
J774A.1 cells (Ralph and Nakoinz, 1975) (DSMZ, Braunschweig, Germany) were cultured in RPMI 1640 medium, supplemented with 10% FCS, 1%
U937 cells (Sundstrom and Nilsson, 1976)—a human macrophage cell line (kindly provided by Dr. Robert Bals, Institute of Pneumology, Philipps University of Marburg, Germany)—were cultured in RPMI 1640 medium, supplemented with 10% FCS, 1%
Luciferase assay using TLR-9 expressing human embryonic kidney cells
Human embryonic kidney (HEK)293 cells (Graham et al., 1977) stably transfected with a murine TLR-9 construct and a nuclear factor-kappa B (NFκB)-luciferase reporter were provided by Prof. Bauer (Institute of Immunology, Philipps University of Marburg, Germany) (Bauer et al., 2001). Cells were cultured in Dulbecco's modified Eagle's medium high glucose medium (Biochrom AG, Berlin, Germany) supplemented with 10% FCS, 1%
Stimulation of human mast cells and rat basophils and β-hexoaminidase assay
HMC-1, a human mast cell line (Butterfield et al., 1988), was provided by Dr. Petersen (Research Centre Borstel, Germany) and cultured in Iscove medium F0456 (Biochrom) with 10% FCS, 1%
Rat basophil cell line (RBL)-2H3 cells, a (Eccleston et al., 1973), stably transfected to express the human Fcɛ receptor 1 were plated at 4×104 cells/100 μL in 96-well tissue culture plates. Stimulants were added in a volume of 100 μL and incubated at 37°C for 30 minutes. Ionomycin (4 μg/mL) was used as positive control, incubation with Triton-X 100 (Sigma) served as 100% release control. In an additional experiment RBL-2H3 cells were passively sensitized with human immunoglobin E (IgE) (Alpha Diagnostics Int.) for 2 hours prior to incubation with hgd40 (2.8 μM) and the respective controls Triton-X 100 and ionomycin for 30 minutes. Analysis of β-hexoaminidase release was performed as described previously (Scholl et al., 2007).
Stimulation of neutrophil granulocytes and superoxide radical assay
Superoxide radical release assay was performed as described (Olker et al., 2004) using human granulocytes. Briefly, lithium heparinate whole blood was taken from healthy donors and the erythrocytes were removed by repeated suspension with lysis buffer [8.99 g NH4 (Merck), 1 g KHCO3 (Merck), and 0.037 g EDTA (Roth, Karlsruhe, Germany) in 1 l aqua dest.]. Cells were suspended in RPMI 1640 medium with 10% FCS, 1%
In vivo off-target effects
In vivo off-target effects of oligonucleotides were analyzed in 6- to 8-week-old Balb/c mice (Harlan Winkelmann) that were maintained under specific pathogen-free conditions. Diet and water were supplied ad libitum. Animal studies were performed with the approval of the local animal ethics committee. Mice were treated by intraperitoneal injection of murine 1668 CpG, gd21, or hgd40, each 200 μg dissolved in 50 μL PBS (PAA), as this was shown to be the optimal dose in intranasal treatments (11). Twenty-four hours later peritoneal lavage was performed with 5 mL PBS and blood was collected. IL-12p40 levels were measured in lavage fluid as well as in serum by enzyme-linked immunosorbent assay (ELISA).
Measurement of cytokine concentrations
Murine IL-6, IL-12p40, and human tumor necrosis factor (TNF)-α were determined using BD OptEIA–ELISA kits (BD Pharmingen, San Diego), and murine TNF-α was measured by use of the Cyto Set Mouse ELISA (Invitrogen, Karlsruhe, Germany) according to manufacturer's instructions, each determination performed in duplicates.
Reverse transcription-polymerase chain reaction analysis of TLR-9 expression
RNA was isolated from human pDC by use of the RNeasy Mini Kit (Qiagen, Hilden, Germany). DNA was removed with Deoxyribonuclease I Amplification Grade (Invitrogen, Karlsruhe, Germany), and cDNA synthesis was performed by Omniscript© Reverse Transcription (Qiagen). Polymerase chain reaction PCR for detection of human TLR-9 expression was performed with HotStarTaq Master Mix Kit, using the primers TLR-9-sense: CCTATTCATGGACGGCAACT; TLR-9-antisense: GAGTGACAGGTGGGTGAGGT and the following PCR conditions: 15-minute 95°C initial activation step; 1-minute 94°C denaturation, 40-second 60°C annealing, 1-minute 72°C extension, 45 cycles; and 10-minute 72°C final extension. Resulting PCR products were detected by standard agarose gel electrophoresis.
Statistical analysis
Results are presented as mean values±standard of the mean (SEM) calculated by GraphPad Prism®. Results were analyzed for statistical significance using one-way analysis of variance and Newman-Keuls test.
Results
Effects of DNAzymes on TLR-9 activation
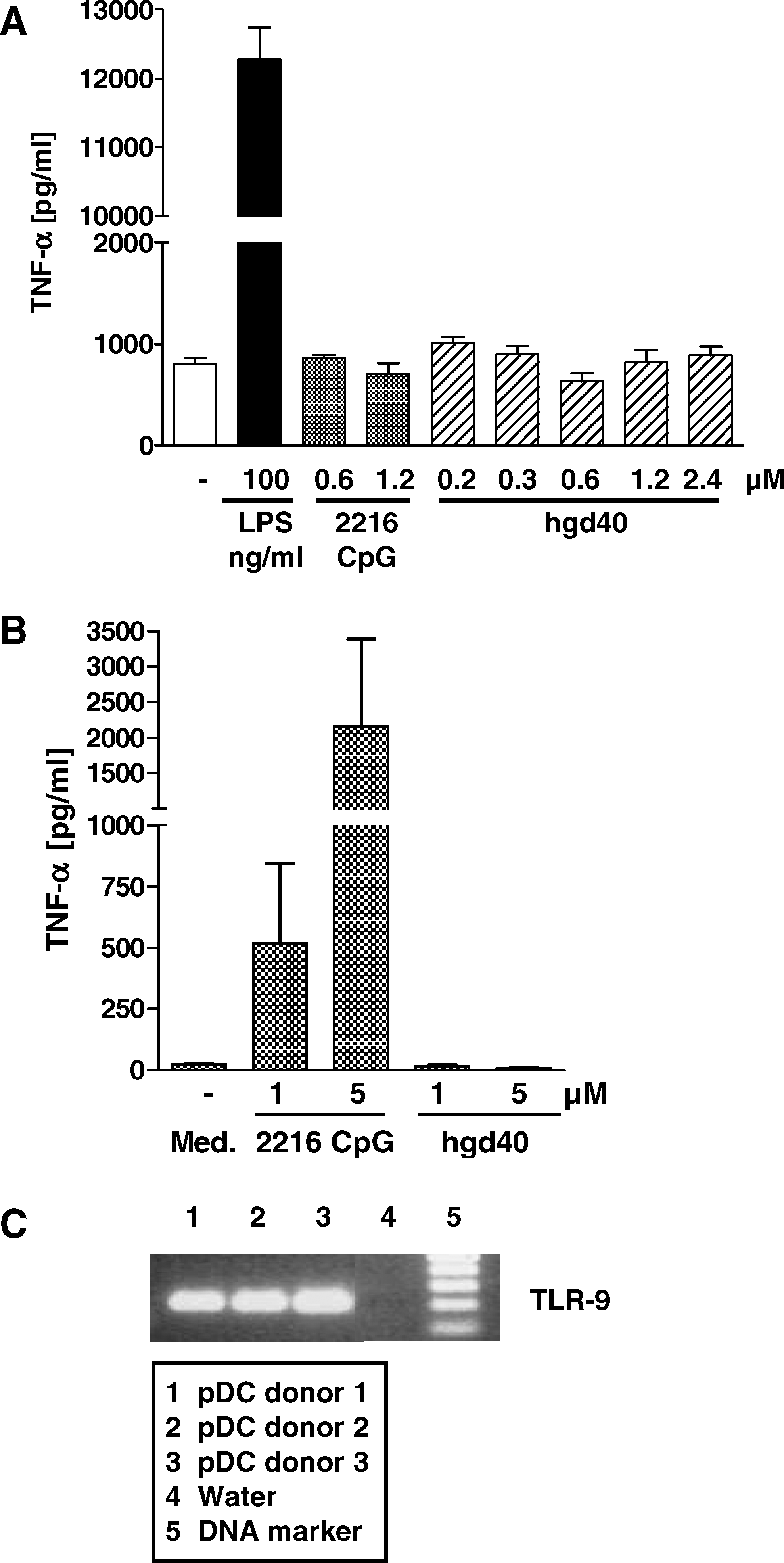
Cells of the innate immune system may be directly activated by DNA molecules via TLR-9, which recognizes single-stranded desoxyoligonucleotides with unmethylated CpG-motifs (Wagner and Bauer, 2006). DNAzymes of the 10-23 family in general contain such a motif in the catalytic domain and further CpG motifs may be present in the binding sequences (see Fig. 1B). The human GATA-3-specific DNAzyme hgd40 contains only the CpG motif in the conserved catalytic sequence and the murine GATA-3-specific gd21 has an additional CpG motif in one of the binding arms. To specifically address the question whether the DNAzymes may activate cells directly via TLR-9, initial analyses were performed using HEK293 cells stably transfected with murine TLR-9 leading to an artificially high constitutive TLR-9 expression and an NFκB luciferase reporter construct. These cells were incubated with the DNAzymes hgd40 and gd21 and the TLR-9 agonist 1668 CpG as a positive control for 24 hours in increasing concentrations, and TLR-9-induced NFκB activation was monitored. As shown in Fig. 2, no luciferase activity was observed in DNAzyme-treated cells up to the highest tested concentration of 5 μM in contrast to cells incubated with the TLR-9 agonist 1668 CpG, indicating the absence of TLR-9-activating capacities of the tested DNAzymes.

Human embryonic kidney (HEK)293 cells co-transfected with murine Toll-like receptor 9 (TLR-9) and a nuclear factor-kappa B (NFκB) reporter vector show no activation after incubation with DNAzymes. HEK293 cells were stably transfected with a murine TLR-9 construct and a NFκB luciferase reporter vector. Cells were incubated with up to 5 μM of DNAzymes for 24 hours. The known TLR-9 ligand 1668 CpG served as positive control. Luciferase activity was assessed and resulting luminescence is shown in relative light units (RLU). The dotted line represents the background activity of the cells in medium. Shown are means±standard errors of the means (SEMs) of 1 out of 2 independent experiments.
In vitro stimulatory activities of DNAzymes on murine macrophages and primary mononuclear cells
It has been shown that other molecules than TLR-9 may be involved in the recognition of foreign DNA molecules in innate immune cells. Therefore, all further experiments were performed with cells of innate immune cell lineages. First, murine macrophages of the J774A.1 cell line (Fig. 3A) and primary mononuclear splenocytes (Fig. 3B) isolated from untreated healthy Balb/c mice were stimulated with DNAzyme hgd40 and control oligonucleotides, and inflammatory cytokine production was measured. As shown in Fig. 3, neither GATA-3-specific DNAzyme hgd40 nor the control DNAzyme ODNg did induce any cytokine production (IL-6, IL-12p40, TNF-α) in these cells, even up to concentrations of 5 μM. As expected, only the murine TLR-9 agonist 1668 CpG was able to induce the production of all analyzed cytokines.

Murine macrophages and splenocytes do not release inflammatory cytokines after incubation with DNAzymes.
In vivo activities of DNAzymes on innate immune mechanisms
In order to analyze off-target effects of GATA-3-specific DNAzymes in vivo, Balb/c mice were treated intraperitoneally with 200 μg of the oligonucleotides hgd40, gd21, 1668 CpG, and ODNg. Twenty-four hours later, peritoneal lavage and serum were analyzed for the presence of the proinflammatory cytokine IL-12p40. With the exception of TLR-9 agonist 1668 CpG treated animals, no induction of IL-12p40 was observed in either peritoneal lavage or in serum of any of the other groups, indicating that GATA-3-specific DNAzymes do not exert innate immune cell activation in vivo (Fig. 4).
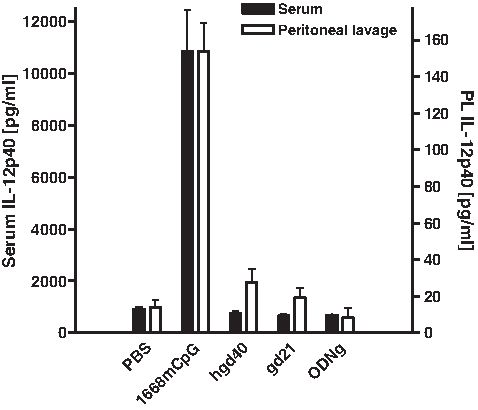
Intraperitoneal treatment of naïve Balb/c mice with DNAzymes does not induce production of IL-12p40. Naïve Balb/c mice were intraperitoneally injected with 200 μg of DNAzymes and control oligonucleotides. Twenty-four hours later peritoneal lavage was performed and blood samples were collected. IL-12p40 concentrations were determined by ELISA in serum (black bars) and peritoneal lavage fluid (PL, open bars). Means±SEMs are shown of 4 animals investigated in 2 independent experiments.
Effects of DNAzymes on human macrophages and pDC
With regard to human clinical trials, off-target effects were also analyzed in human cells. At first, human macrophages of the U937 cell line were differentiated with PMA and afterwards stimulated with hgd40 and the controls LPS and 2216 CpG. As expected, TNF-α production by these cells was induced by the TLR-4 ligand LPS. In contrast, the TLR-9 ligand 2216 CpG as well as hgd40 did not induce any TNF-α release (Fig. 5A). Furthermore, TLR-9 expressing pDC were isolated from human buffy coats and stimulated with hgd40 and the positive control 2216 CpG at 1 and 5 μM. A significant TNF-α production was induced by the TLR-9 ligand 2216 CpG, whereas hgd40 was not able to stimulate pDC to release detectable amounts of TNF-α (Fig. 5B). TLR-9 expression of pDC was proven by qualitative RT-PCR (Fig. 5C).
Human macrophages and primary plasmacytoid dendritic cells (pDC) do not release tumor necrosis factor (TNF)-α following incubation with DNAzyme.
Respiratory burst of human neutrophils after DNAzyme treatment
Inflammatory cytokine release is only a single parameter of innate immune cell activation and does not represent the major function of granulocytes. However, the activation of neutrophils results in a profound generation and release of superoxide radicals, known as respiratory burst. To investigate potential stimulatory activities of DNAzymes on neutrophils, superoxide radical release of these cells was investigated using a lucigenin-enhanced chemiluminescence system (Olker et al., 2004). This analysis was performed with human granulocytes, which were isolated from whole blood and treated with hgd40 and ODNg in increasing concentrations; zymosan A particles from yeast served as positive control. Neither hgd40 nor ODNg (data not shown) induced any superoxide radical release in human neutrophil granulocytes in contrast to zymosan A (Table 1).
Human neutrophils were isolated from whole blood of healthy donors and stimulated with increasing concentrations (0.3–4.7 μM) of the DNAzyme hgd40 and zymosan A particles (0.03×108 to 2×108 particles/mL) as positive control. Superoxide radical release was analyzed by lucigenin-enhanced chemiluminescence. Values represent integrated relative light units (RLU) released over a time period of 85 minutes following start of incubation.
Effect of DNAzymes on human mast cells and basophils
GATA-3-specific DNAzymes are intended as therapeutic treatment of allergic disorders. Since mast cells and basophils play a central role in the context of these diseases, potential direct effects of GATA-3-specific DNAzymes on these cell types were also investigated in the present study. Therefore, human mast cells of the HMC-1 cell line were stimulated with hgd40 in increasing concentrations or with ionomycin as a positive control. As read-out, β-hexoaminidase content was analyzed in the supernatants. Hgd40 did not cause any direct degranulation of human mast cells (Fig. 6). Similar results were obtained with basophils of the RBL-2H3 line stably transfected to constitutively express the human Fcɛ receptor 1 (Fig. 7A). These results suggest that the tested DNAzymes do not exert direct mast cell or basophil stimulating activities. To exclude that hgd40 contributes to IgE-mediated Fcɛ receptor cross-linking and subsequent cell activation, an additional experiment was performed with RBL-2H3 cells that were passively loaded with human IgE and subsequently treated with hgd40 at different concentrations. Also in this setting hgd40 was not able to induce basophil degranulation (Fig. 7B).
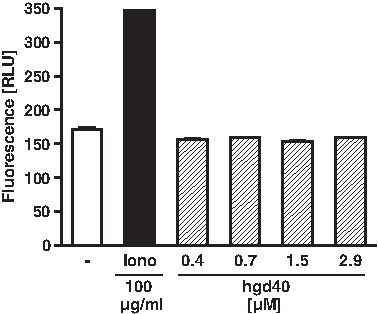
Human mast cell line (HMC)-1 cells show no β-hexoaminidase release after incubation with DNAzyme. Cells of the human mast cell line HMC-1 were incubated for 30 minutes with increasing amounts of hgd40 (0.4–2.9 μM) and with 100 μg/mL ionomycin (Iono) as positive control. β-hexoaminidase release was analyzed as indicator of mast cell degranulation. Fluorescence intensity is provided in RLU.
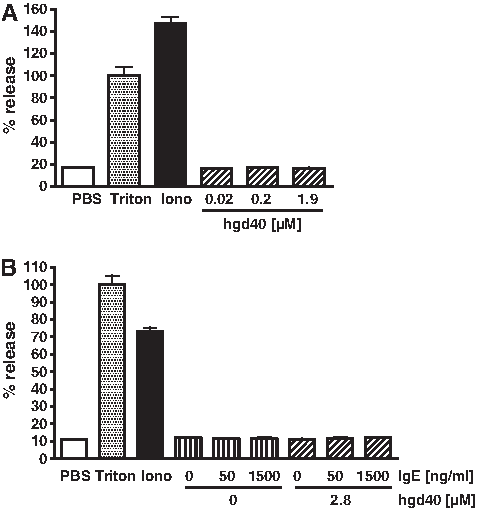
Basophils expressing the human Fcɛ receptor do not degranulate in response to incubation with DNAzyme. Rat basophils (RBL-2H3 cells) stably transfected to express the human FcɛR1 were either
Discussion
Investigation of potential off-target effects is an important issue in the development of novel antisense-based therapeutics. Such effects may be sequence-specific (binding to unwanted mRNA or DNA targets) or sequence-independent (binding to general receptors or unspecific interaction with macromolecules), even though the latter are also determined by the actual sequence, for example, by formation of motifs such as unmethylated CpGs recognized by PRRs. There are several reports on potential off-target effects of oligonucleotides (Kleinman et al., 2008; Baum et al., 2009; Dass and Choong, 2010); however, so far DNAzymes have not been investigated in comparable detail.
We have developed 10-23 DNAzymes specific for the mRNA of the transcription factor GATA-3 which represents the master transcription factor of Th2-driven (allergic) immune responses. Their efficacy has recently been demonstrated using experimental models of allergic airway inflammation in mice following local application into the lung (Sel et al., 2008). Especially when using this application route, therapeutic molecules (in our case DNAzymes) will directly get in contact with first line defense cells of the innate immune system. Thus, it was the objective of the current study to investigate potential off-target effects of GATA-3-specific DNAzymes with focus on unwanted stimulation of innate immune mechanisms. In conclusion, no adverse or unwanted stimulatory effects were observed for the tested DNAzymes in any of the systems investigated; that is, TLR-9 dependent and independent activation of innate immune cells such as macrophages or dendritic cells, unspecific activation of granulocytes, or unwanted stimulation of mast cells.
Since one unmethylated CpG motif is inherent in the conserved catalytic region of all DNAzymes of the 10-23 family and further CpGs may be present in the binding regions as exemplarily shown for gd21, we investigated DNAzyme effects on cells expressing TLR-9. Initially, HEK293 cells were analyzed that were transfected to selectively express artificially high levels of murine TLR-9 in combination with a NFκB luciferase reporter construct. Even under these conditions no signs of TLR-9 activation could be observed. These results were further confirmed by use of the murine macrophage cell line J774 or primary splenocytes as well as with human macrophages of the U937 cell line or primary plasmacytoid dendritic cells.
Experiments were performed with cells from both species for 2 major reasons: (1) TLR-9 expression patterns differ significantly between murine and human cells and (2) rodent cells are more susceptible to CpG-containing immunostimulatory DNA sequences (Campbell et al., 2009). Whereas in mice TLR-9 is constitutively expressed on the majority of innate immune cells, among them monocytes, macrophages and all dendritic cell subsets, constitutive TLR-9 expression in humans is restricted to B cells and plasmacytoid DCs (Iwasaki and Medzhitov, 2004). Therefore, the latter cell population was included in the investigations, and also here no unspecific activation was observed after DNAzyme incubation despite proven TLR-9 expression in these cells. With regard to the differences in species susceptibility to CpGs the lack of murine innate immune cell activation by the applied DNAzyme molecules in the in vitro and even more important in vivo studies strongly support the conclusion that these molecules do not act via unspecific activation of innate immune mechanism even in the human system.
Recently, a series of additional DNA-sensing molecules has been identified that may also lead to innate immune cell activation (Hornung and Latz, 2010). Among them, DAI (DLM/ZBP1) was shown to recognize free cytosolic DNA molecules followed by activation of 2 independent signaling pathways, the interferon regulatory factor and the NFκB pathway, which both induce the expression of proinflammatory cytokines such as type-1 interferons, IL-6, and TNF-α (Takaoka et al., 2007). A further cytoplasmic DNA sensor is a molecule called AIM2, the activation of which leads to inflammasome activation involving caspase-1 (Burckstummer et al., 2009). Both molecules preferentially recognize dsDNA; however, it cannot be excluded that ssDNA molecules such as DNAzymes may also be detected by these or additional DNA-sensing receptor molecules. However, these molecules have been shown to be expressed in a variety of innate immune cells, among them cells that have been used in our study. Thus, recognition of the investigated DNAzymes by these molecules and subsequent activation of the respective cells would also have been observed in the applied systems if present. Taken together, since no unspecific activation of any of the innate immune cells investigated was observed we conclude that the tested DNAzymes do not lead to unspecific activation of these cells, neither via TLR-9 nor any other DNA-sensing receptor. Such mechanisms might actually have contributed to the anti-allergic properties of our DNAzyme molecules by subsequent activation of Th1 cells, which then counter-regulate Th2-driven development of allergic immune responses. Indeed, such approaches have been shown to be effective in preclinical and initial clinical studies using immunostimulatory CpG-containing DNA sequences (Santeliz et al., 2002; Hessel et al., 2005; Fonseca and Kline, 2009; Senti et al., 2009; Gupta and Agrawal, 2010). However, these approaches are not in line with the objectives of our DNAzyme concept that aims at the specific down-regulation of pathologically enhanced adaptive immune mechanisms instead of activation of potentially counter-regulating immunological processes.
DNA molecules of general size and structure of 10-23 DNAzymes may not only interact with cellular structures via specialized DNA-sensing receptors, but also by unspecific binding to other macromolecules, for example, protein targets. This so-called aptamer property is based on the formation of stable 3-dimensional structures that are capable of high-affinity binding to target molecules and subsequent modulation of their biological function in a comparable manner to antibodies (Bunka and Stockley, 2006; Bouchard et al., 2010). Binding of DNAzymes to cellular surface molecules by this mechanism might also interfere with functional properties of the respective cells and should be excluded mainly for those cells involved in effector mechanisms of allergic immune responses. Therefore, the influence of the GATA-3-specific DNAzyme hgd40 on the activity of neutrophil granulocytes, mast cells, and basophils was investigated without showing any signs of direct stimulating effects. Especially with regard to mast cells and basophils, it was of importance to demonstrate the absence of Fcɛ receptor 1 crosslinking (with and without bound IgE), which would have induced unwanted activation of these cells. Additionally, the lack of activation of the different investigated innate immune cells indicated that also these cells are not stimulated by putative aptamer effects of the tested DNAzymes.
In conclusion, the results of this study extend our previous findings (Sel et al., 2008) on the action of GATA-3-specific DNAzymes in the treatment of allergic lung diseases by excluding the possibility of unspecific innate immune cell activation as underlying mechanism. Moreover, the general lack of activation of innate immune cells and cells contributing to the development of the allergic phenotype suggests a favorable safety profile of the GATA-3-specific DNAzyme molecule hgd40 for its application in the treatment of human diseases.
Footnotes
Acknowledgments
This work was supported by the Deutsche Forschungsgemeinschaft (SFB/TR22) and Sterna Biologicals GmbH & Co. KG. Isabella Pali Schöll was supported by the Austrian Science Fund (Hertha Firnberg project T283-B13). We thank Dr. Petersen from Research Centre Borstel, Germany, for providing HMC-1 cells. We acknowledge the expert technical assistance of Thomas Ruppersberg, Wilma Hasted, Nicole Disser, Anja Spies-Naumann, and Sophia Bernhardt.
Author Disclosure Statement
Tanja Dicke and Holger Garn are employees and Harald Renz and Holger Garn are shareholders of Sterna Biologicals.