
Abstract
Vascular endothelial growth factor (VEGF) is the important angiogenic factor associated with tumor growth and metastasis in a wide variety of solid tumors. The aim of this study is to investigate the tumor suppressive effect of chitosan/small interfering RNA (siRNA)-VEGF nanoplexes in the rat breast cancer model. Chitosan/siRNA nanoplexes (siVEGF-A, siVEGFR-1, siVEGFR-2) and NRP-1 were prepared in a 15 to1 ratio and injected (intratumorally) into the breast-tumor-bearing Sprague-Dawley rats. Tumor volumes were measured during 21 days. To investigate the effect of chitosan/siRNA nanoplexes on VEGF expression in tumors, VEGF was analyzed with immunohistochemistry and western blotting. The mRNA levels of VEGF in tumor samples were determined with real-time PCR (RT-PCR). After siRNA treatment, a marked reduction in tumor volumes was measured in complex-injected rats (97%). Free siRNA injection showed lower tumor inhibition. Reduction of VEGF protein was also shown with western blotting and immunohistochemistry. Similar results were obtained with RT-PCR also. These results indicate that the chitosan/siRNA targeting to VEGF nanoplexes have a remarkably suppressive effect on VEGF expression and tumor volume in breast cancer model of rats.
Introduction
VEGF is the most potent, known as direct-acting, angiogenic protein and several pathologies are associated with the upregulation of the VEGF pathway. The VEGF family consists of 6 growth factors that bind to and activate 3 distinct receptors. VEGF-A binds to VEGFR-1/Flt-1 and VEGFR-2/Flk-1/KDR, whereas VEGF-B binds to only VEGFR-1. VEGF-C and VEGF-D bind to VEGFR-2 and VEGFR-3/Flt-4 (Robinson et al., 2001; Lu et al., 2005). These receptors have functions as signaling molecules during vascular development. Neuropilin-1 and −2 (NRP-1 and NRP-2) are co-receptors for VEGF (Donovan et al., 2006).
VEGF and its receptors have been associated with angiogenesis that occurs in many solid tumors including breast, colon, hepatoma, bladder, gastric, and prostate cancers. Since formation of solid tumors is angiogenesis dependent, several strategies have been developed for targeting the VEGF pathway as a part of anticancer therapy (MCMAHON, 2000). The inhibition of VEGF signaling at the receptor level has been proposed as a therapeutic strategy for the treatment of solid cancers (SHIBUYA, 1995).
RNA interference (RNAi) has rapidly become an important tool in gene therapy for targeting various diseases such as cancer and neurological and viral infectious diseases. RNAi-mediated silencing of oncogenic regulators that play major roles in tumor transformation, growth, and metastasis has been considered a promising strategy for cancer therapy (Kim et al., 2008).
One of the main challenges in developing small interfering RNA (siRNA)-based therapies is the delivery of the siRNA molecule to the target cells due to its poor in vivo stability and inefficient cellular uptake (Kim et al., 2009). Therefore, a number of carriers have been developed for improving in vivo delivery of siRNA. As siRNAs are negatively charged and readily bind to cationic molecules, delivery carriers usually consist of cationic polymers and liposomes (Kim et al., 2007; Murata et al., 2008). Among them, chitosan, a biopolymer, also has many advantages as a siRNA carrier, including biodegradability, low toxicity, and biocompatibility (Oh et al., 2009). Numerous studies on siRNA delivery with chitosan as a carrier have shown effective suppression of reporter genes in vitro and in vivo (Howard et al., 2006; Howard et al, 2009; Zhang et al., 2005). Howard et al. (2006) have developed a novel chitosan-based siRNA nanoparticle delivery system for RNAi in vitro and in vivo. They showed nanoparticle-mediated knockdown of enhanced green fluorescent protein in human lung carcinoma cells (H1299) and murine peritoneal macrophages (77.9% and 89.3% reduction, respectively). In another study, the same group showed the efficient silencing of tumor necrosis factor-α (TNF-α) expressed in macrophages by the intraperitoneal administration of chitosan/siRNA nanoparticles (Howard et al., 2009). Zhang et al. (2005) showed siRNA protection effect of chitosan nanoparticles against RSV, and low molecular weight protamine were reported for VEGF-targeting siRNAs (Kim et al., 2007; Kim et al., 2006; Lee et al., 2007; Choi et al., 2010).
In this study, we investigated the silencing effect of chitosan nanoplexes of siRNA targeting VEGF expression and inhibition of tumor growth in rat breast cancer model after intratumoral administrations. Therefore, we used nanoplexes containing chitosan and mixture of siRNAs such as siVEGF-A, siVEGFR-1, siVEGFR-2, and siNRP-1 for tumor inhibition.
Materials and Methods
Materials
We purchased low molecular weight chitosan (75 kDa, 75–85% deacetylation) from Sigma and used as gene carriers. The siRNA oligonucleotides used in this study were purchased from Dharmacon. A thorough analysis by BLAST was performed on all siRNA sequences used in this study to demonstrate that these sequences have no homology to other genes. The target sequence of the anti-rat VEGF-A siRNA was UUUAAAUCCUGGAGCUUC, sense strand: 5’-dTdTCUUGCGAGGUCCUAAAUUU-3’ and antisense strand: 5’-GAACGCUCCAGGAUUUAAAdTdT-3’; the target sequence of the VEGFR1 (FLT1) was GGAGAAAUCUGCUCGCUAU, sense strand: 5’dTdTUAU CGCUCGUCUAAAGAGG-3’, and antisense strand: 5’-AUAGCGAGCA GAUUUCUCCdTdT-3’; the target sequence of the VEGFR2 (KDR or FLK1) was UGAAAGAAGGAACGAGAAU, sense strand: 5’dTdTUAAGAGCAAGGAAGAAAGU-3’ and antisense strand: 5’-AUUCUCGUUCCUUCUUUCAdTdT-3’; and the target sequence of the neuropilin-1 (NRP-1) was CACAAUGGGAUGUCGGAAA, sense strand: 5’dTdTAAAGGCUGUAGGGUAACAC-3’ and antisense strand: 5’-UUUCCGACA UCCCAUUGUGdTdT-3’. The sequences of a mismatch VEGF-A siRNA were 5’-dTdTCUUGAGAGGCCCUAGAUUU-3’ (sense) and 5’-GAACUCUCCGGGAUC UAAAdTdT-3’ (antisense).
N-Nitroso-N-methylurea (NMU) was obtained from Sigma-Aldrich. Monoclonal VEGF-A antibody (mAb) and chemiluminescence substrate kit were purchased from Santa Cruz Biotechnology. All other chemicals and reagents were of analytical and molecular grade and were purchased from Sigma-Aldrich.
Methods
Preparation of chitosan/siRNA nanoplexes
Chitosan was dissolved in 0.1 M Tris acetate/0.1 M acetate buffer (pH 5.4) to form a solution of 2.5 mg/mL. Lyophilized siRNAs were dissolved in 5× siRNA buffer, and final concentrations were made of 1μg/μL. Chitosan to siRNA ratio was used according to our previous study (Şalva et al., 2010a). Chitosan nanoplexes (15/1, +/−) were prepared by adding chitosan solution to siRNAs stock solutions during intense stirring on a vortex mixer for 15 seconds and the nanoplexes were left for 30 minutes for forming of the nanoplexes completely. The formations of nanoplexes were checked by agarose gel electrophoresis in phosphate buffered saline (PBS).
The diameters and zeta potentials of the nanoplexes were determined using a Malvern Zetasizer (Malvern NanoZess). The instrument was equipped with both a particle sizer and a zetameter unit. The samples were measured in PBS in glass cuvettes at 25°C with a constant angle of 90°. Each measurement was done in triplicate.
In vivo gene silencing study
In vivo intratumoral efficiencies of siRNAs (siVEGF-A, siVEGFR-1, siVEGFR-2, siNRP-1) containing nanoplexes were studied in the breast cancer model of Sprague Dawley female rats (40 days of age). All animal experiments were performed in accordance with the acceptance and guidance of Animal Ethics Committee of Marmara University (37.2006.mar).
Breast tumors were induced by injection of 50 mg/kg NMU into the peritoneal cavities of rats. After 3 or 4 months, tumors developed along the mammary lines from the axilla to inguinal areas of rats (Thompson et al., 1991). After the tumor formation, rats were randomly allocated to 5 groups: chitosan/siRNAs nanoplexes (intratumoral; i.t), free siRNAs (i.t), chitosan (i.t), chitosan/mismatch siRNA nanoplexes (i.t) and untreated (positive control group) (each group, n=5). The untreated control group received intratumoral injections of PBS (Phosphate buffered saline pH 7.0).
Tumor size was measured using a caliper across its longest (a) and shortest diameters (b) and its volume was calculated using the formula of V=0.5ab2. siRNA treatment of the tumors was started when the tumor size reached approximately 50–60 mm3 (Kim et al., 2008; Schmitz, et al., 2006).
Chitosan/siRNAs nanoplexes (w/w ratio of 15/1, 375 μg chitosan/25 μg siRNAs nanoplexes) were prepared in MiliQ water and 400 μL of the nanoplexes was directly injected twice a week into the tumor of rats at a dose of 25 μg siRNAs per rat. The animals in chitosan alone group were injected chitosan at high dose (2.5 mg/mL).
Tumor volumes were measured every 5 days and rats were examined for appearance and growth of necrosis as well as decreased physical activity (Matsumoto et al., 2006). In addition to tumor volume measurements, the following tests were carried out: quantitative RT-PCR for mRNA and immunohistochemistry and western blotting for VEGF.
Immunohistochemical analysis
Immunohistochemical staining for VEGF-A was performed. After deparaffinization and rehydration, sections were incubated with 3% hydrogen peroxide for elimination endogenous peroxidase activity. Nonspecific binding sites were blocked with protein blocking serum for 5 minutes and incubated with rat anti-mouse VEGF-A mAb (dilution 1:200; Santa Cruz) for 60 minutes at room temperature. After rinsing in PBS, sections were incubated in UltraVision Detection System Anti-Polyvalent HRP (Lab Vision) according to the manufacturer's instructions. PBS-washed sections were incubated with 3,3’-diaminobenzidine until a brown color was visible and then counterstained with Mayer hematoxylin and mounted (Schmidt et al., 2008).
Expression levels of VEGF were analyzed by scoring the extent and intensity of staining. Only tumors that had good quality staining of microvessels with the StrABC/HRP (streptavidin biotin complex/horseradish peroxidase) technique were evaluated in this study. After the areas of highest neovascularization had been identified, all microvessels were counted on a 1×400 field and according to their staining density. For semi-quantitative evaluation of staining distribution, a scoring scale from 1 to 4 was defined: score 1 indicating staining of less than 10% of tumor cells, score 2 indicating 10–49%, score 3 indicating 50–79%, and score 4 indicating staining of 80% or more. Staining intensities were also scored. Score 0 indicates no staining, whereas score 1 indicates when cytoplasmic staining was weak or focal, score 2 moderate, and score 3 when cytoplasmic staining in all tumor tissue was strong and diffuse (Schmidt et al., 2008; Bolat et al., 2006; Turhal et al., 2004).
RNA extraction and quantitative real-time PCR (RT-PCR) for VEGF
For RNA isolation, breast tumor tissues were stored in RNeasy later solution (Qiagen). After homogenization of tissues, mRNA was extracted with the RNeasy Plus Mini Kit (Qiagen) according to the supplier's protocol. The concentration, purity, and amount of total RNA were determined using spectrophotometry and 1% agarose gel. mRNA retrotranscription into cDNA was made using Transcriptor High Fidelity cDNA Synthesis Kit (Roche) according to the manufacturer's instructions. Real-time PCR was performed in a mixture containing 5 μl cDNA, 2 pmol universal probe library (UPL), 4 pmol reverse and forward primer, 1
Western blot for VEGF
In order to test VEGF-A protein expression, western blotting was applied to tumor samples. Frozen tissues were homogenized in buffer (RIPA lysis buffer: 1
Analysis of IFN-γ levels in rats
To in vivo study of immune response to chitosan/siRNA nanoplexes, serum interferon (IFN)-γ levels were measured using the Endogen rat interferon gamma ELISA kit according to the manufacturer's instructions. The results were analyzed using ELISA reader at 450 nm (Ma et al., 2005).
Statistical analysis
Tumor data were given as mean tumor volumes with standard error of the mean. For all graphs presented in this study, each data point represented the mean of at least 3 replicates. Either the standard deviation of the mean (SD) or the standard error of the mean (SEM) was calculated and used to make the error bars. Student's t-tests were applied for comparisons between two means. For more than 2 means, a one-way or two-way analysis of variance with a 95% level of confidence (ANOVA, α=0.05) was used. Unless otherwise stated, a P-value<0.05 was considered to indicate statistical significance.
Results
Preparation and control of nanoplexes
To check the formation of chitosan/siRNA (VEGF-A) nanoplexes, the gel retardation assay was performed. Full complexation was observed at chitosan/siRNA (+/−) nanoplexes in 15 to 1 ratio (Fig. 1). The size and surface charge of chitosan/siRNA nanoplexes (15/1) were measured around 276±10.7 nm and +16.1±4.3 mV, respectively.

Agarose gel photograph of chitosan/small interfering RNA (siRNA)-vascular endothelial growth factor VEGF (siVEGF) nanoplexes.
Influence of siRNA treatment on breast tumor growth
We investigated the anti-tumor effects of chitosan/siRNA (VEGF-A, VEGFR-1, VEGFR-2, NRP-1) nanoplexes after intratumoral injections on breast tumors of rats. While tumor volume was strikingly increasing in untreated (positive control) rats, intratumoral injections of chitosan/siRNA nanoplexes significantly suppressed tumor growth (Fig. 2). In the treatment with siVEGF of the experimental breast tumor model, treatment with chitosan/siVEGF nanoplexes reduced tumor volume of 94% (final volume; 90 mm3±30, n=5) compared with the untreated tumor control group (final volume; 2632 mm3±52, n=5) (Fig. 2) (P<0.001). Intratumoral treatment of chitosan/mismatch siVEGF nanoplexes (39%) and free siVEGF (36%) decreased tumor growth comparable with untreated controls, but effect on tumor volume was low (Fig. 2). Intratumoral treatment of a slow-growing breast tumor with nanoplexed siVEGF caused a significant delay in tumor growth in contrast to the nanoplexed mismatch siVEGF.
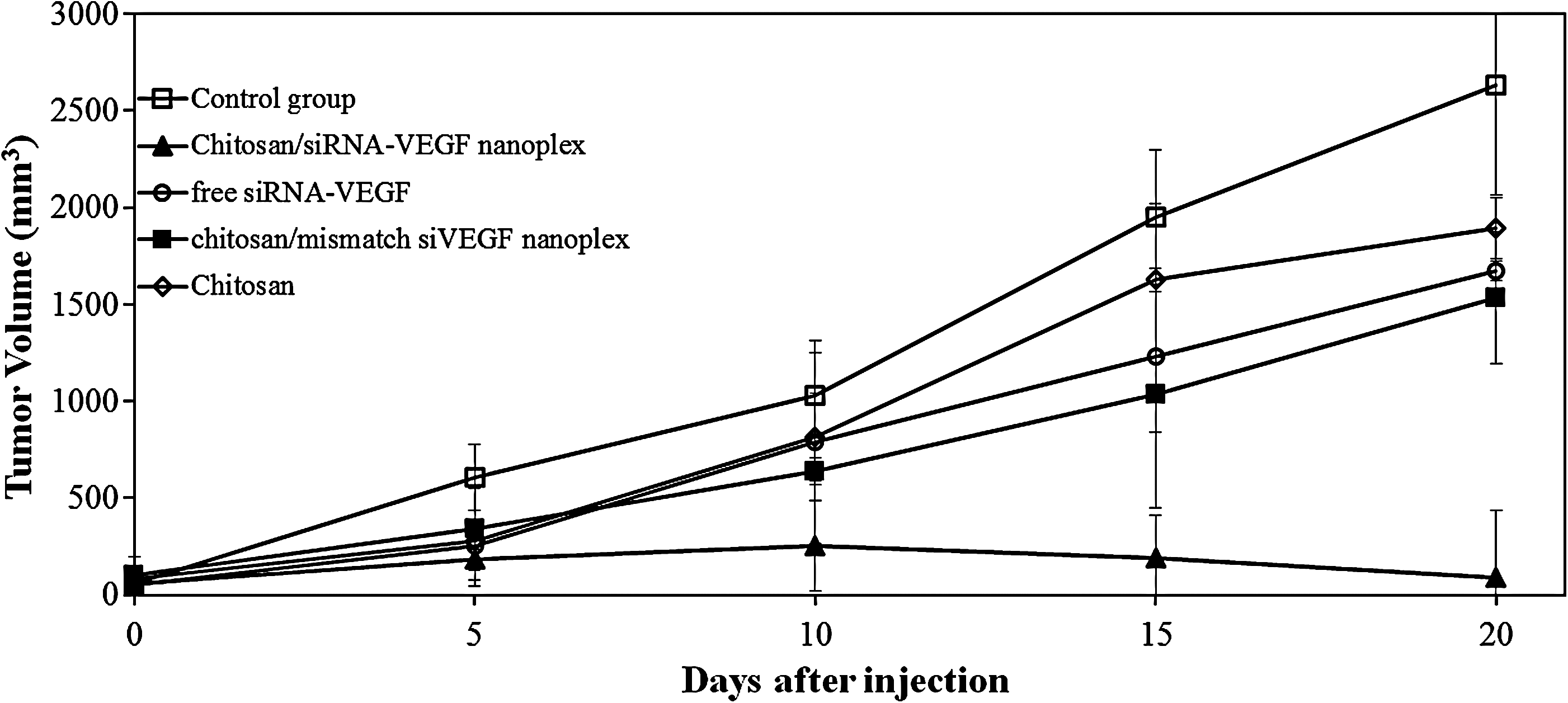
Effects of intratumoral (i.t) injection of chitosan/siRNA nanoplexes on tumor growth in rats. Chitosan/siRNAs nanoplexes were injected into rats i.t as indicated by arrows. On day 1, the tumor volume was approximately 50–60 mm3. Tumor diameters were measured with calipers at a regular interval for up to 20 days.
Immunohistochemical analysis
Gene inhibition effect of chitosan/siRNA nanoplexes on tumor tissues was investigated with the Str.ABC/HRP method using light microscopy. Areas of invasive tumor containing the greatest numbers of capillaries and small venules (areas of most intense neovascularization), which most frequently occurred at the margins of the carcinoma, were examined by microscopic and were found by scanning the tumor sections at low power.
Few positively stained endothelial cells could be detected in siRNAs injected tumors, whereas positively stained endothelial cells were readily detectable in positive control tumors (Fig. 3 A–D). As observed in photographs, when VEGF immunexpression and microvessel density of siVEGF injected tumor groups were compared to the untreated group, a statistically significant difference between them (P<0.05) was seen. On the other hand, when all the groups compared according to reduction of VEGF immunexpression, the following ranks were obtained: chitosan/siRNA i.t 80%>chitosan/mismatch siRNA i.t 51%>free siRNAs i.t 44%. The highest VEGF protein inhibition was found in chitosan/siVEGF nanoplex injected tumor (Fig. 3b).
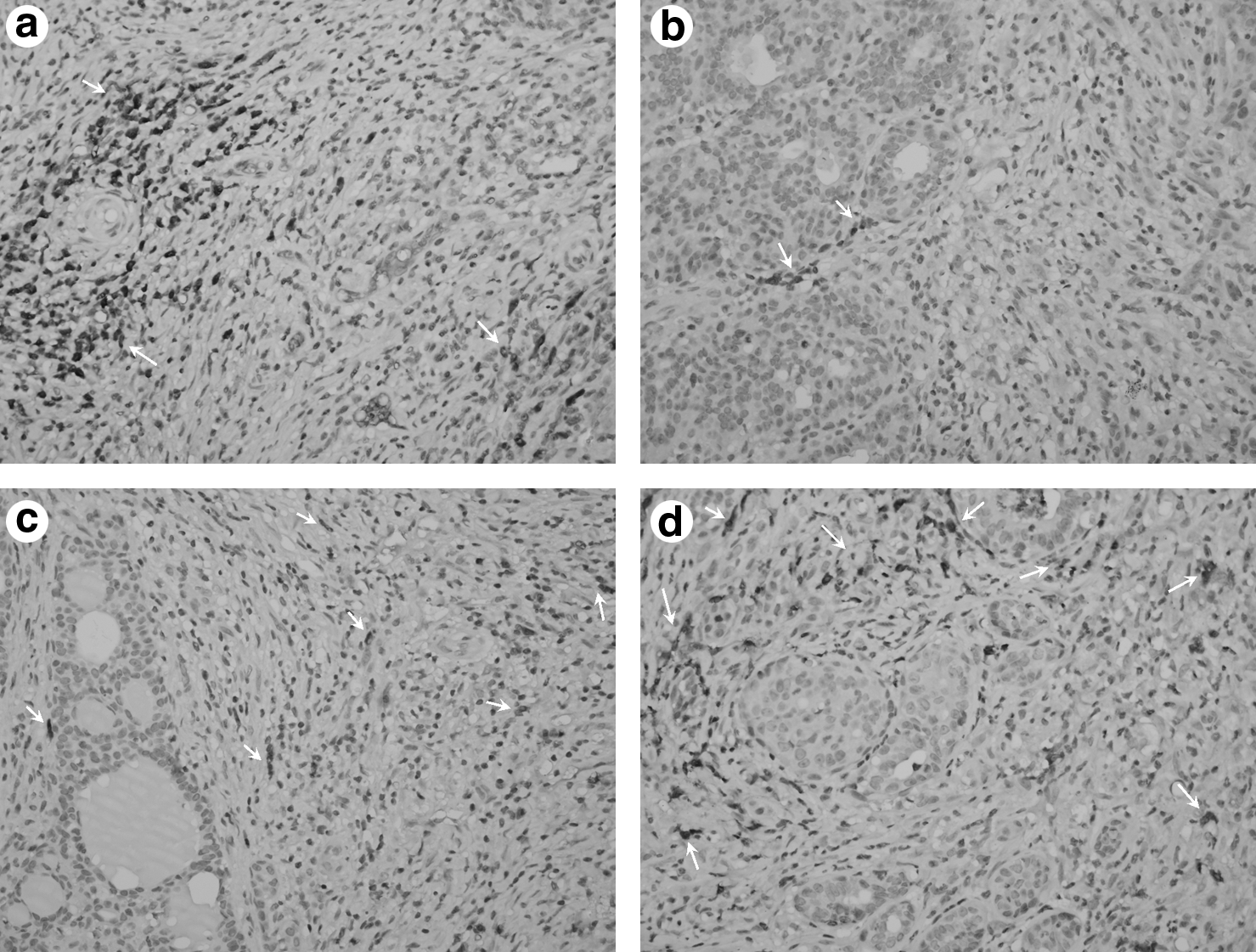
Immunohistochemical analysis of tumor blood vessels. VEGF immunexpression was analyzed by staining with VEGF monoclonal antibody (400
Effect of siVEGF on the expression of VEGF
As shown in Fig. 4, the relative VEGF mRNA level in tumor tissue after injection with chitosan/siRNA nanoplexes was markedly reduced in comparison with the level of control group. We observed that the chitosan/siRNA nanoplexes injected group showed the highest mRNA inhibition (93%). In the other groups, free siRNAs and chitosan/mismatch siRNA nanoplexes also showed a decline in the VEGF level. This result indicates that the tumor-suppressive effect is particularly due to the chitosan/siVEGF nanoplexes.
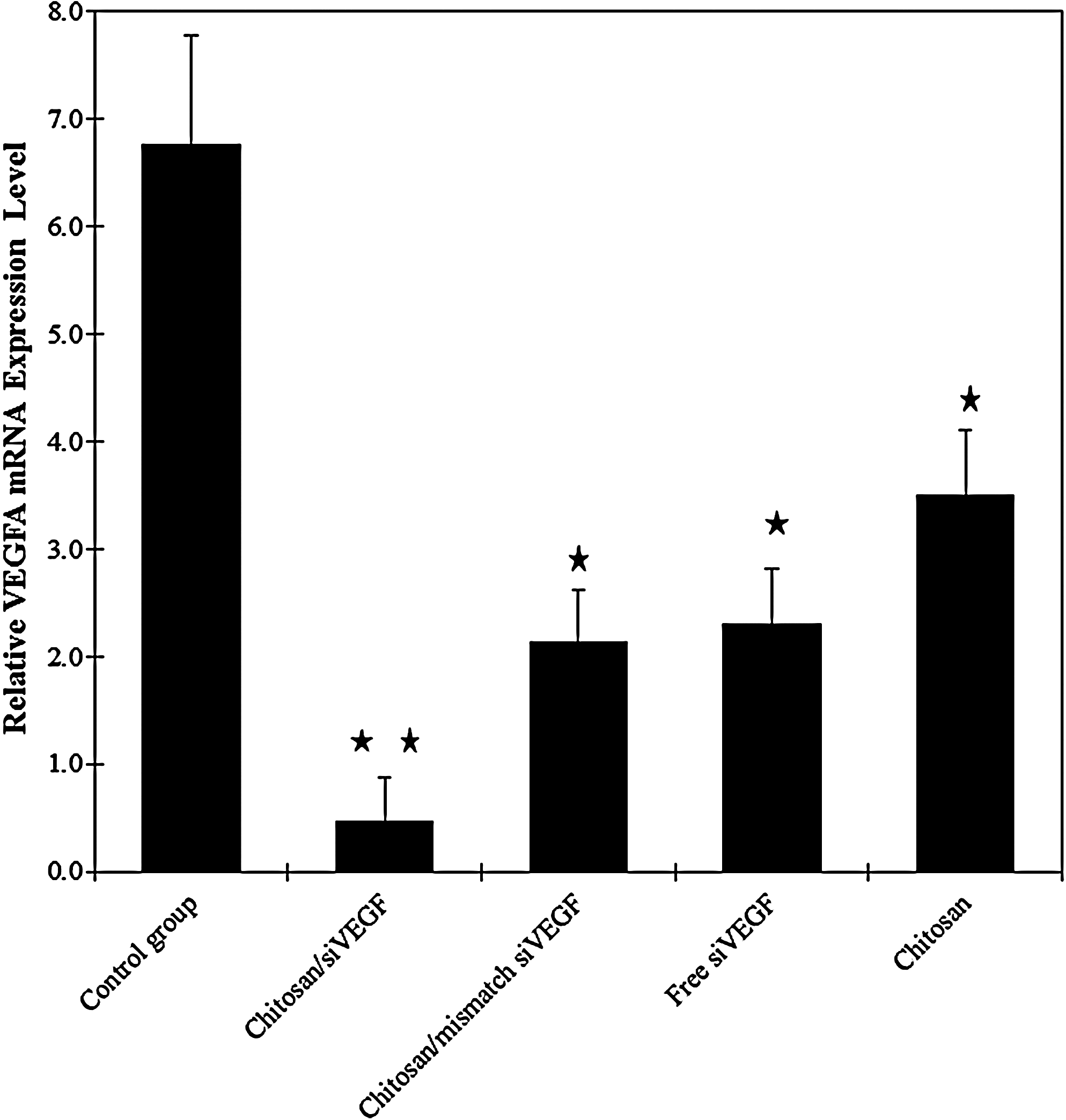
Effect of synthetic siRNA on VEGF expression in rats with breast tumor. VEGF mRNA levels were quantified by real-time-PCR. Data were normalized with the housekeeping gene beta-actin. Data are represented as mean±SD of triplicate wells. Similar results were obtained in 3 independent experiments.
In addition, western blot analyses showed a significant decrease in VEGF-A protein levels in the chitosan/siVEGF nanoplexes group, compared with the control group (Fig. 5).

Western blotting for VEGF. Inhibition rates of siVEGF. Lane 1, untreated control group; Lane 2, free siVEGF; Lane 3, chitosan; Lane 4, chitosan/mismatch siVEGF nanoplex; Lane 5, chitosan/siVEGF nanoplex.
Analysis of IFN-γ for immune response to siRNA
To determine the immunostimulatory capacity of siRNAs, rats were scarified at 21 days after siRNA therapy and serum collected. IFN-γ levels were assessed by ELISA. As shown, chitosan and siRNA did not lead to the cytokine production (P>0.05) (Fig. 6).
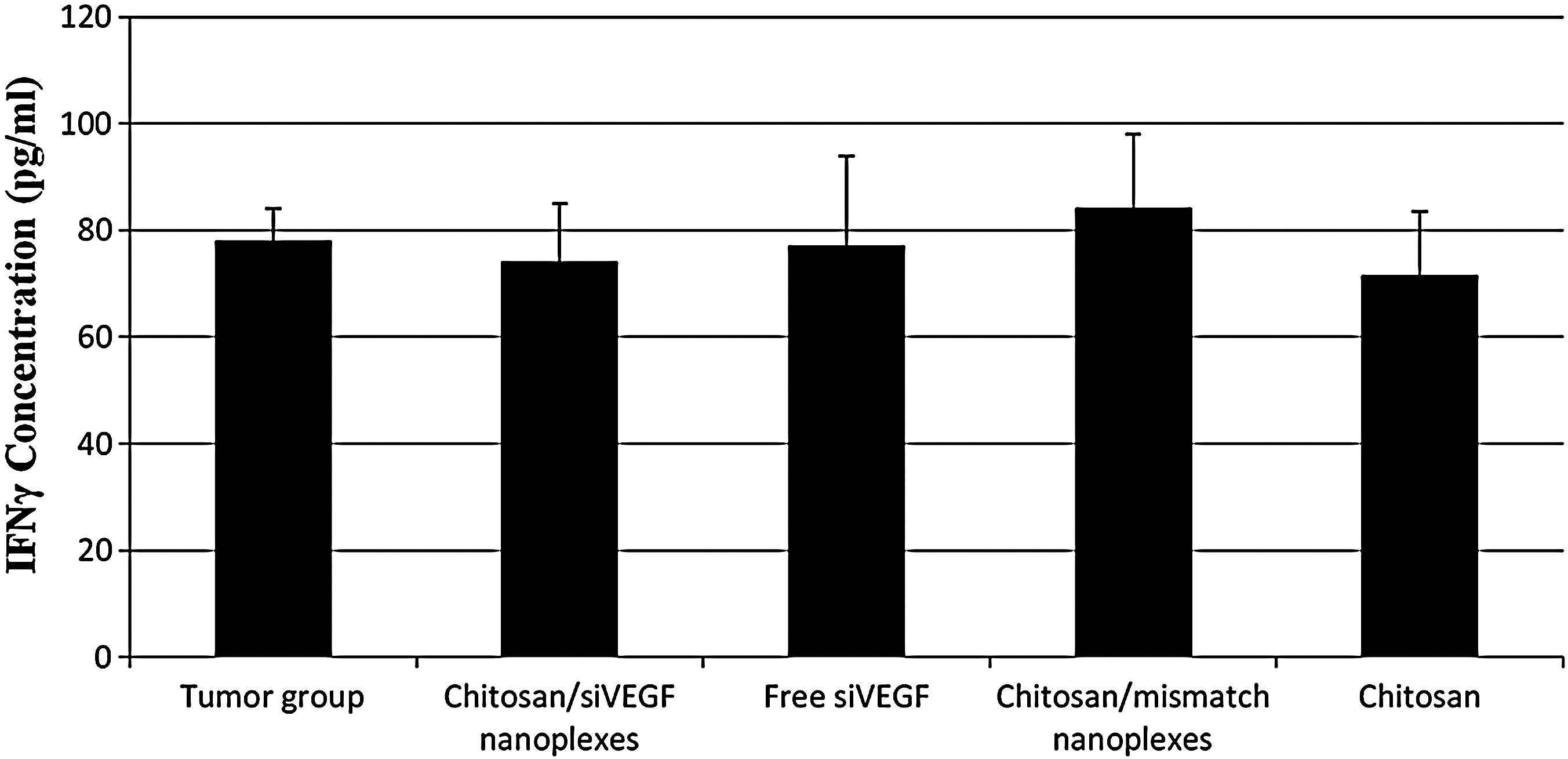
Effect on interferon (IFN)-γ production of siRNA in various groups after intratumoral treatment. Twenty-one days after treatment initiation, rats were sacrified and serum collected. IFN-γ levels were measured by ELISA.
Discussion
Cancer is characterized by the uncontrolled growth of new blood vessels. This uncontrolled process of new blood vessel growth is an important pathogenic cause of tumor growth. Thus, anti-angiogenic therapy has become an important route for cancer treatment (DETMAR, 2000). VEGF, an angiogenic factor, has received considerable attention (FERRARA, 2004). Inhibition of genes using siRNA in vitro and in vivo has improved studies on the mechanism of action for many disease genes, including those involved in the angiogenesis process (Lu et al., 2005). In cell culture-based studies, the expression of VEGF was inhibited using siRNA in HeLa cells, ovarian carcinoma cells, and melanoma cells (Yin et al., 2003). However, their therapeutic potential will be realized where the in vivo efficiency of the siRNAs can be achieved with clinically feasible delivery systems (Lu et al., 2005).
Cationic polymer or lipid-based delivery systems for anti-cancer siRNA have been studied in breast cancer cell lines and animal tumor models (Meryet-Figuires et al., 2007; Sutton et al., 2007; Şalva et al., 2010b; Schifflers et al., 2004). The inhibition of breast tumor growth by the intratumoral delivery of siRNA targeting VEGF has been demonstrated in the MDA-MB-435 xenograft model previously (XIE, 2004). In another study, the suppression of tumor angiogenesis was observed in a Ptd Cho-3 xenograft model after intratumoral VEGF siRNA administration (Takei et al., 2004).
The uptake of siRNA by non-target tissues with systemic administration is responsible for loss of the majority of the injected dose (Schiffelers et al., 2005). Therefore, most studies using siRNA to date have focused on different delivery systems and local delivery to target organs (Reich et al., 2003). Many researchers have studied how to effectively deliver siRNA to target cells and tissues. The delivery system has to be internalized into the cell, the siRNA has to be released in a reversible manner to conserve its activity in the cytosol, and the gene silencing pathway has to be activated (Murata et al., 2008). In order to increase the activity of siRNA, effective delivery systems must be developed. The use of chitosan as a nucleic acid delivery vehicle has several potential advantages over viral delivery. The interactions between the negatively charged cell membrane and the positively charged chitosan/siRNA nanoplexes increase cell uptake of siRNA. Loss of the injected dose is prevented by the systemic delivery of chitosan/siRNA nanoplexes (Schiffelers et al., 2005). Therefore, the purpose of our current study is to find the availability of chitosan as a carrier for synthetic siRNA targeting to VEGF expression in the rat breast cancer model.
Angiogenesis-related diseases are caused by the abnormal overexpression of multiple genes. Kim et al. (2004) showed a significant inhibitory effect of the mixture of siRNAs when given either locally or systemically in vehicle against HSV-induced angiogenesis. The combination of siRNAs targeting VEGF-A, VEGF-R1, and VEGF-R2 had demonstrated an improved anti-angiogenesis potency than siRNAs targeting only one gene (Kim et al., 2004). Therefore, we used a combination of siRNAs targeting VEGF-A, VEGFR-1, VEGFR-2, and also NRP-1 in our study. NRP-1 contributed to tumor growth via tumor angiogenesis and their direct effects on tumor cells. This activity was shown in different mouse tumor models and NRP-1 levels were enhanced in tumors of many types. Therefore, NRPs were shown to be the potential targets for anti-angiogenic cancer therapy (Geretti et al., 2008).
The siRNA combinations mentioned above were mixed with chitosan/siRNA nanoplexes in a 15 to 1 ratio for nanoplex formation. Chitosan/siRNA nanoplexes of approximately 276±10.7 nm were obtained and used. Before nanoplexes were given to animals, a biopsy was taken, and tumor identification was made pathologically. We started the treatment with siRNA when the tumor size reached about 50–60 mm3. Bergers et al. (2003) have shown that the makeup of the tumor vasculative varies at different stages of tumor development, so the inhibitor effect might depend on its application during a specific phase of tumorigenesis. This reveals the importance of VEGF signaling during the angiogenic switch in initial tumor growth, but not in large tumors (Takei et al., 2004). As seen in Fig. 2, tumor growth was very slow during the first 10 days after i.t application of nanoplexes. Then, reduction in tumor volumes was observed. Animals receiving chitosan/siRNAs nanoplexes (97%) indicated considerably reduced tumor growth compared to the control. This data shows accordance with the earlier paper (Murata et al., 2008). Murata et al. (2008) investigated anti-tumor effects of VEGF siRNA encapsulated poly(lactic-co-glycolic acid) (PLGA) microspheres in mice after i.t injection and showed that higher and sustained suppressive effect on VEGF gene expression was possible after siRNA-PLGA microsphere injection.
Furthermore, the results of tumor volume suppression were supported by RT-PCR, western blotting, and immunohistochemistry results (Figs. 2, 3–5). The RT-PCR was used to evaluate VEGF knockdown efficiency. As shown in Fig. 4, VEGF mRNA expression was significantly decreased when the siRNA was injected by chitosan nanoplexes to rats (P<0.05). Western blotting was used to further investigate the silence expression of VEGF in protein levels (Fig. 5). Compared to VEGF protein expression in control and other groups, the VEGF protein in chitosan/siRNAs nanoplexes group was significantly suppressed (P<0.05). According to our immunohistochemical data as seen in the Fig. 3, when the VEGF immunexpression was compared in all groups, VEGF expression and microvessel density markedly increased in untreated breast tumors (Fig. 3) (P<0.05). As a result, a higher decline in VEGF expression was obtained when chitosan/siVEGF nanoplexes (80%) were applied.
Chitosan has been shown to be a promising carrier system for DNA and siRNA in vitro and in vivo delivery because of their biocompatibility, biodegradability, and high cationic charge (Mao et al., 2010). In addition, chitosan has been shown to inhibit tumor growth by reducing glycolysis and decreasing glucose uptake and ATP levels and inducing apoptosis via activation of caspase-3 in tumor cells (Harish Prashanth et al., 2005; Qin et al., 2002; Dass et al., 2008; Hasegawa et al., 2001). As shown in Fig. 2, the tumor growth inhibitory effect of chitosan was observed at high dose (2.5 mg/mL). Furthermore, a decrease in the mRNA levels was also shown at this dose (Fig. 4). However, these effects were not observed at the low dose used in the preparation of our nanoplexes (data not shown).
In our study, the modest reduction in VEGF mRNA and protein was observed with the mismatch siRNA. Many studies were available related to the silencing effect of mismatch sequences (Naito et al., 2004; Martin et al., 2006; Ahmed et al., 2011). Our findings were consistent with the literature. We used the software presented in Naito et al. (2004) for selecting of the mismatch sequence. Martin et al. (2006) reported that mismatched siRNAs targeting endogenous mRNAs appear to reduce in protein and mRNA levels. In addition, Ahmed et al. (2011) demonstrated that mismatches between siRNA and target site and/or position of the mismatch generated in target affects the silencing efficacy.
The long, double-stranded RNAs induce the interferon system in mammalian cells. To overcome this problem, siRNAs that are shorter than 30 bp have been designed for use as potential therapeutic agents (Reynolds et al., 2006). It was recently reported that the immunostimulatory activity of siRNAs was also dependent on the nucleotide sequence (Judge et al., 2005; Kim et al., 2008). Double-stranded RNAs and some siRNAs activated a multi-component signaling complex that includes protein kinase R and signal transducer. The off-target and nonspecific effects of siRNAs have been caused the activation of the type I and II IFN pathways (Chang et al., 2011) It should also be noted that IFN-γ has also been demonstrated to suppress growth and to induce apoptosis of breast and ovarian cancer cells in vivo and in vitro (Treeck et al., 2005). In our study, serum IFN-γ levels were measured to determine whether free siRNA and chitosan/siRNA nanoplexes administered by i.t. injection to tumor-bearing animals led to immune response (Fig. 6). Analysis of IFN-γ expression showed that chitosan and siRNA did not induce an interferon-mediated immune response compared to the tumor control group, implying that anti-tumor effects were mainly VEGF specific.
Conclusion
Small interfering RNAs have been used as a new therapeutic agent for various angiogenesis-related diseases; however, their use remains problematic because of unresolved difficulties in delivery of the siRNA to tumor cells. We demonstrated potent growth inhibiting effects of VEGF siRNA on breast carcinoma in vivo, potentially applicable to the treatment of cancers as an anti-angiogenic therapeutic for siRNA delivery. On the other hand, this study also showed that chitosan was a suitable carrier for siRNA targeting to VEGF in breast cancer.
Footnotes
Acknowledgments
This study was supported by the Marmara University Scientific Research Projects Association (SAG-DKR-290906-0185) and Turkish Association for Cancer Research and Control (Terry Fox Cancer Foundation). We wish to thank them for financial support, and to thank M.E. Salva for help in designing the figures.
Author Disclosure Statement
No competing financial interests exist.