
Abstract
MicroRNAs (miRNAs) are post-transcriptional regulators of gene expression, interplaying with transcription factors in complex regulatory networks. Menin is the product of the MEN1 oncosuppressor gene, responsible for multiple endocrine neoplasia type 1 syndrome. Recent data suggest that menin functions as a general regulator of transcription. Menin expression modulates mesenchymal cell commitment to the myogenic or osteogenic lineages. The microRNA 26a (miR-26a) modulates the expression of SMAD1 protein during the osteoblastic differentiation of human adipose tissue-derived stem cells (hADSCs). We used siRNA silencing against MEN1 mRNA and pre-miR-26 mimics to study the interplay between them and to investigate the interplay between menin and miR-26a as regulators of osteogenic differentiation in the hADSCs. We found that in hADSCs the siRNA-induced silencing of MEN1 mRNA resulted in a down regulation of miR-26a, with a consequent up-regulation of SMAD1 protein. Chromatin immunoprecipitation (ChIP) showed that menin occupies the miR-26-a gene promoter, thus inducing its expression and confirming that menin is a positive regulator of miR-26a. In conclusion, results from this study evidenced, for the first time, a direct interaction between menin transcription factor and miRNA, interaction that seems to play a pivotal role during the hADSCs osteogenesis, thus suggesting a novel target for bone disease RNA-based therapy.
Introduction
MicroRNAs (miRNAs) are a family of naturally occurring, evolutionarily conserved, small (approximately 19–23 nucleotides), non-protein-coding RNAs that negatively regulate post-transcriptional gene expression. miRNAs are estimated to account for >3% of all human genes, and they control expression of thousands of target mRNAs, with multiple miRNAs targeting for a single mRNA (Flynt and Lai, 2008). Transcription factors (TFs) are essential regulators of gene expression. A TF regulates transcription of target genes by specifically binding to the TF binding site in the promoter region of genes. Since expression of a miRNA may be regulated by a TF, TFs and miRNAs may regulate each other to form feedback loops, or alternatively, both TFs and miRNAs may regulate their target genes and form feed-forward loops (FFLs) (Saini et al., 2007).
Using osteoblast precursors obtained from subcutaneous human adipose tissue-derived stem cells (hADSCs), induced to differentiate to osteoblast lineage by dexamethasone, we previously demonstrated that microRNA 26a (miR-26a) levels increased in differentiating osteoblasts, and that miR-26a inhibition, by 2′-O-methyl-RNA, produced an increase in SMAD1 TF protein levels, up-regulating osteoblastic marker genes and thus enhancing osteoblast differentiation (Luzi et al., 2008). In the present study, we investigated the possible interaction between the TF menin and miR-26a, as regulators of the early dexamethasone-induced osteoblastic differentiation of hADSCs, by silencing MEN1 mRNA.
Materials and Methods
The materials used in this study were purchased from Sigma-Aldrich (St. Louis, MO) unless otherwise specified. All the experiments have been performed on 3 different hADSC cell lines.
Cell cultures and osteogenic differentiation
Primary cultures of hADSCs were isolated and induced to differentiate toward the osteogenic phenotype as described previously (Luzi et al., 2008).
Transient transfection
Cells were grown in osteogenic medium, Ham's F12 Coon's modification medium supplemented with 10% fetal bovine serum (South America origin, BioWhittaker), 100 IU/mL penicillin, 100 μg/mL streptomycin, 10 nm dexamethasone, 0.2 mm ascorbic acid (Merck, Darmstadt, Germany), and 10 mm β-glycerolphosphate for 18 days and were then transfected using Lipofectamine 2000 (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions and incubated for 48 hours at 37°C. After that, the cells were collected and analyzed for both RNAs and proteins expression.
Transfection of small interfering RNAs (siRNAs) was carried using 200 pmol of specific duplexes for MEN1 mRNA (MEN1 siRNA) or scrambled sequence control siRNA (C siRNA), purchased from IBA (Gottingen, Germany). The sense and antisense strands of siRNAs employed were, respectively: 5′-GUCGCAAGUGCAGAUGAAGdTdT-3′ and 5′-CUUCAUCUGCACUUGCGACdTdT-3′ for MEN1, and 5-GCCAAUAUUGAAACCCACGGdTdT-3′ and 5′-UUCGGUUAUAACUUUGGGUGdTdT-3′ for scrambled sequence. Cells transfected with only lipofectamine (mock control) have been used as control as well. Reporter constructs that contain a miR-26a binding site (pGL3-26a) in the 3′ untranslated terminal region (UTR) of SMAD1 mRNA was kindly given by Dr. David P. Bartel (Howard Hughes Medical Institute, USA). Reporter plasmid (200 ng) was transfected alone or in combination with 40 pmol of siRNAs, using Lipofectamine 2000 (Invitrogen), and hADSCs were transfected at the 18th day as described above. Luciferase assays were performed 48 hours after transfection using the Dual Luciferase Reporter Assay System (Promega). Firefly luciferase activity was normalized to luciferase activity for each transfected well. Each transfected well was assayed in triplicate.
Northern blot analysis
Total RNA was isolated with Trizol reagent (Invitrogen) according to the manufacturer's instructions. For miRNA Northern blots, 15 μg of total RNA were separated on 15% denaturing polyacrylamide gels, electrotransferred to GeneScreen Plus membranes (PerkinElmer, Waltham, MA), and hybridized using UltraHyb-Oligo buffer (Applied Biosystems) at 42°C overnight. Oligonucleotides, complementary to mature miRNAs, were 32P end-labeled with T4 Kinase (Roche Diagnostics, Indianapolis, IN) and were used as probes. Probe sequences were 5′-AGCCTATCCTGGATTACTTGAA-3′ as miR-26a antisense and 5′-TTAGCTTCCGAGATCAGACGATTTTTCCTGTCTC-3′ as 5S antisense.
Primer extension analysis
The RNA pellet (15 μg/5 μL) was first denatured in the presence of [γ-32P]ATP-labeled DNA primer by heating to 80°C for 10 minutes in 5 μL hybridization buffer (60 mM NaCl, 50 mM Tris–HCl, pH 7.5, 5 mM DTT) and then slowly cooled to 30°C. Annealed samples were then added to 5 μL of primer extension buffer (0.5 mM dNTPs, 4 mM MgCl2, 1×hybridization buffer). The reverse transcriptase reaction was initiated by adding 12.5 U of avian myelobastosis virus reverse transcriptase. After 30 minutes of incubation at 50°C the reaction was stopped by addition of an equal volume of gel-loading buffer (95% formamide, 0.025% xylene cyanol, 0.025% bromophenol blue, 18 mM EDTA, 0.025% sodium dodecyl sulfate) and analyzed on a 8% polyacrylamide–7 M urea gel.
Chromatin immunoprecipitation
Chromatin immunoprecipitation (ChIP) was used to detect protein-DNA interactions. ChIP analysis was performed with a commercially available EpiQuik ChIP Kit in accordance with the manufacturer's instructions. The chromatin fraction was immunoprecipitated overnight at 4°C using anti-menin (BL342) polyclonal antibody (Bethyl, Montgomery, TX) and anti-immunoglobin G antibodies, followed by stringent washing and elution. Quantitative polymerase chain reaction (qPCR) amplification analysis was performed in a total volume of 25 μL with specific primers. The forward and reverse primers used were as follows: pri-miR-26a: forward 5′-GCCCAATGGCATAGCAAGA-3′; reverse 5′-GGCCAGTCATGCTTACAGTCAC-3′.
Quantitative reverse transcriptase-polymerase chain reaction
Quantitative reverse transcriptase-polymerase chain reaction (qRT-PCR) analyses were performed as previously reported (Luzi et al., 2008; Luzi et al., 2010).
Western blot analysis
Western blotting analyses to evaluate the expression of SMAD1 and menin proteins were performed as previously reported (Luzi et al., 2008; Luzi et al., 2010).
Statistical analysis
Comparisons were made using a one-way analysis of variance. All experiments were repeated at least 3 times, and representative experiments are shown. Differences were considered significant at P<0.05.
Results
Expression of menin and miR-26a during osteogenic differentiation of hADSCs
hADSCs, induced to differentiate into the osteoblastic lineage, presented the osteoblastic phenotype already at 20 days after induction (Luzi et al., 2008), as confirmed by increased mRNA expression of several specific osteogenic markers (Fig. 1, panel A) and from alkaline phosphatase (ALP) activity analysis (Fig. 1, panel B).

Expression analysis of osteogenic markers and evaluation of the expression of menin protein, MEN1 mRNA, and endogenous microRNA 26a (miR-26a) during hADSC osteoblastic differentiation.
Since the menin is important for the differentiation of osteoblasts, we examined the expression of menin and its mRNA in hADSCs during the osteogenic differentiation steps by western and northern blot analyses, respectively. Both menin protein and MEN1 mRNA showed an increased expression only after osteogenic induction at 10 and 20 days from induction (Fig. 1, panel C). At the 20th day of osteogenic induction, the increase of menin protein and of its mRNA paralleled that of the osteoblastic marker genes, such as SMAD1, ALP, collagen type 1 (COLIA), runt-related transcription factor 2 (RUNX2), osteopontin (OPN), and osteocalcin (OCN) (Fig. 1, panel A). Similarly, primer extension for pre-miR-26a and mature miR-26a at 10 and 20 days from induction showed that the expression of miR-26a increased during osteogenic induction of hADSCs (Fig. 1, panel C).
RNA silencing of MEN1 mRNA
A siRNA against MEN1 mRNA was used to down-regulate the expression of menin protein and to study its effects on miR-26a expression during osteoblastic differentiation of hADSCs (20 days). qRT-PCR and western blot analyses showed that MEN1 siRNA transfection resulted in a complete down-regulation of expression of MEN1 mRNA and menin protein, respectively, when compared to the cells transfected with C siRNA or only with lipofectamine (mock control) (Fig. 2A and B). The expression analysis of endogenous miR-26a by northern blot and qRT-PCR confirmed the effectiveness of MEN1 siRNA to inactivate both MEN1 mRNA and miR-26a expression (Fig. 2C and D). Western blot analysis of the target protein of miR-26a, the SMAD1 protein, showed that while menin was reduced as expected after siRNA down-regulation (Fig. 2B), SMAD1 was up-regulated by the reduction of miR-26a (Fig. 2F). Analysis by qRT-PCR of SMAD1 mRNA did not show any change (Fig. 2, panel E), suggesting that the SMAD1 protein increase was the direct result of miR-26 post-transcriptional down-regulation and, thus, of silencing of MEN1 mRNA.
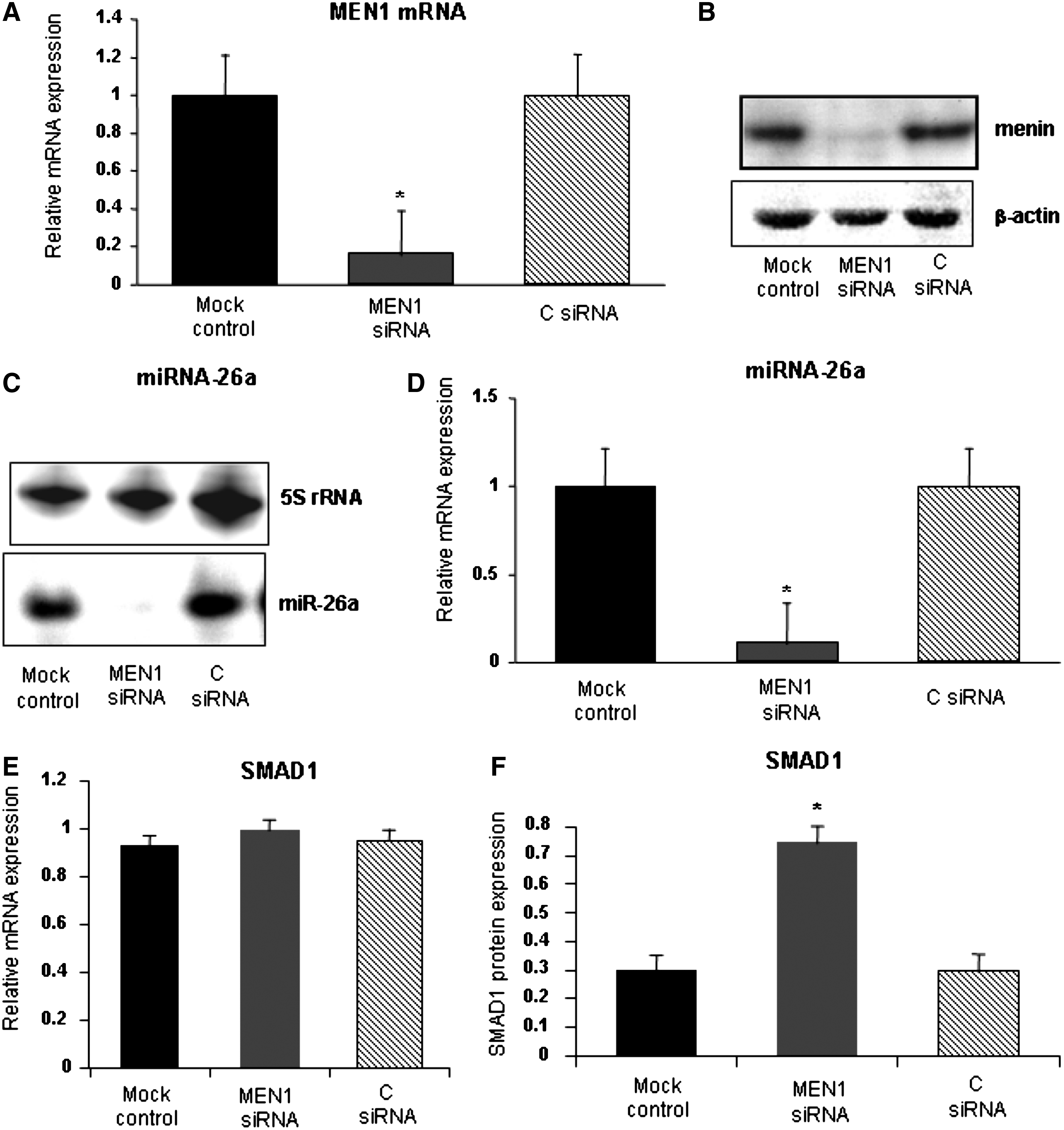
Analysis of expression of MEN1 mRNA and miR-26a in hADSCs transfected with MEN1 small interfering RNAs (siRNA) after 20 days from osteogenic induction. siRNA-induced down-regulation of MEN1 mRNA and menin protein evaluated, respectively, by qRT-PCR
Luciferase analysis of miR-26a expression during early osteoblastic differentiation of hADSCs
The specificity of menin-miR-26 interaction was verified by using pGL3/miR-26a, a luciferase expression plasmid containing 2 complementary miR-26a binding sites (Luzi et al., 2008). The luciferase expression plasmid, pGL3/26a, and siRNA were transfected in differentiating osteoblastic cells. The insertion of miR-26 binding sites rendered the reporter sensitive to endogenous miR-26a expression. Thus, the effect of cellular endogenous miR-26a on translation of the luciferase mRNA was dependent on the presence of the miR-26a cognate binding site within the 3′-UTR. In our case, down-regulation of both menin and miR-26 increased the luciferase activity reported to the negative controls (Fig. 3).
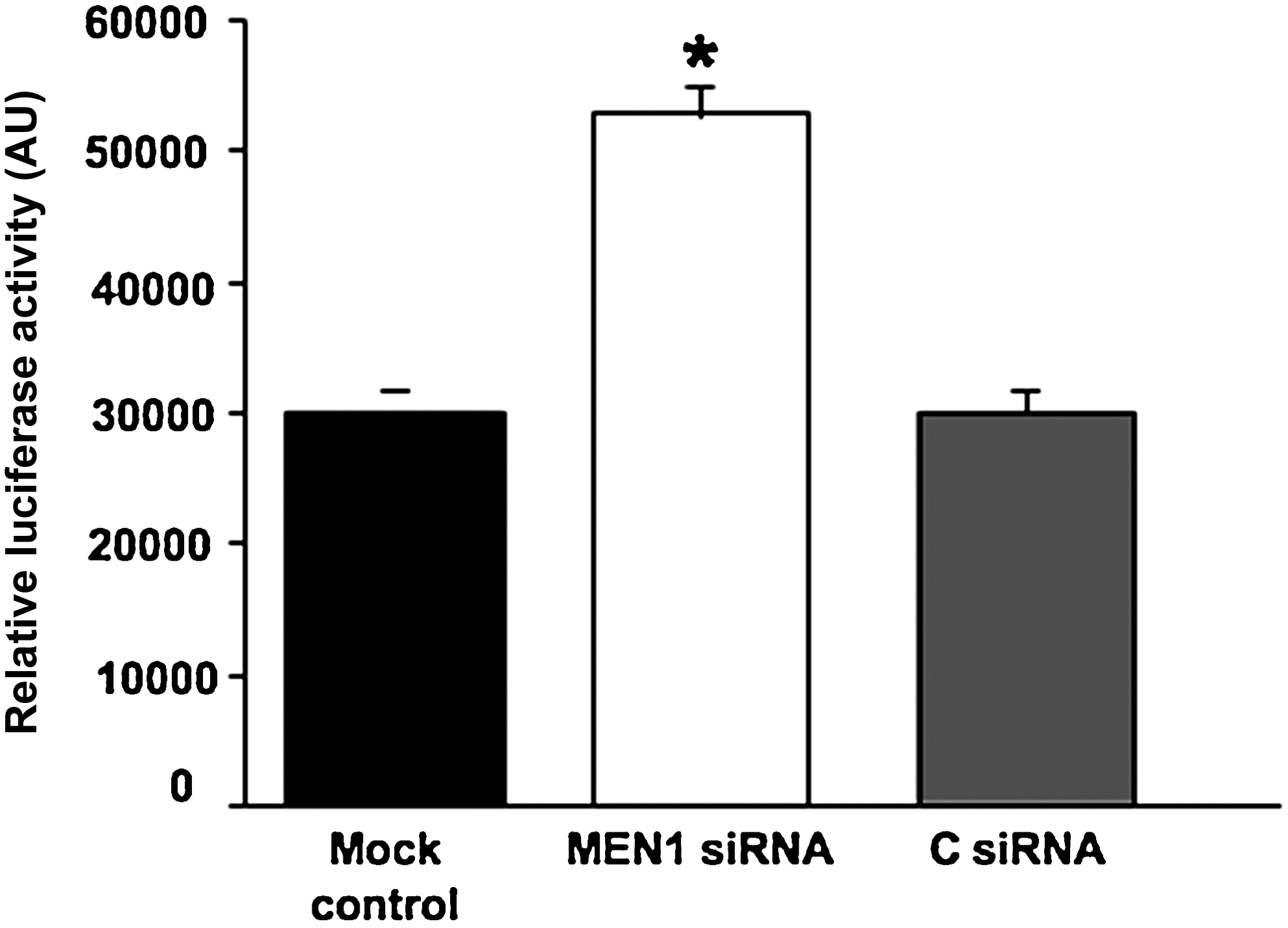
Luciferase assay of miR-26a 3′ untranslated region (UTR) binding activity in siRNA-transfected hADSCs. Reporter luciferase plasmids were transfected into hADSCs with siRNAs at 20 days. Reporter activities have been measured after 2 days in differentiation media and were normalized to Renilla luciferase activity. For all panels, an average of 3 experiments each performed in triplicate on 3 different cell lines with standard errors is presented. *=MEN1 siRNA vs. C siRNA and mock control (P<0.05).
Menin chromatin immunoprecipitation
The data above prompted us to hypothesize a direct involvement of menin in the transcription of miR-26. Menin-binding sites in the promoter of miR-26a were predicted by using the Transcription Element Search Software program (http://www.cbil.upenn.edu/cgi-bin/tess/tess), but we could not find any consensus menin binding sites. That is because, although menin is proven to be a TF, there are no data in the database used by prediction software. To check this hypothesis, ChIP assay for menin was performed using hADSCs transfected cells. Co-precipitated DNAs were analyzed by amplifying the genomic region containing miR-26a promoter region by real-time quantitative polymerase chain reaction (qPCR) (Fig. 4). Occupancy of miR-26a promoter region was lost where siRNA silencing of MEN1 mRNA markedly reduced the interaction between menin and miR26a [Fig. 4, siRNA MEN1 reported to the controls (mock and C siRNA)]. These results indicate that menin occupies the miR-26a gene promoter, thus inducing its expression. This constitutive association of pri-miR26a with menin strengthened our data, thus confirming the specific interaction between menin and miR-26a during hADSCs osteoblast differentiation.
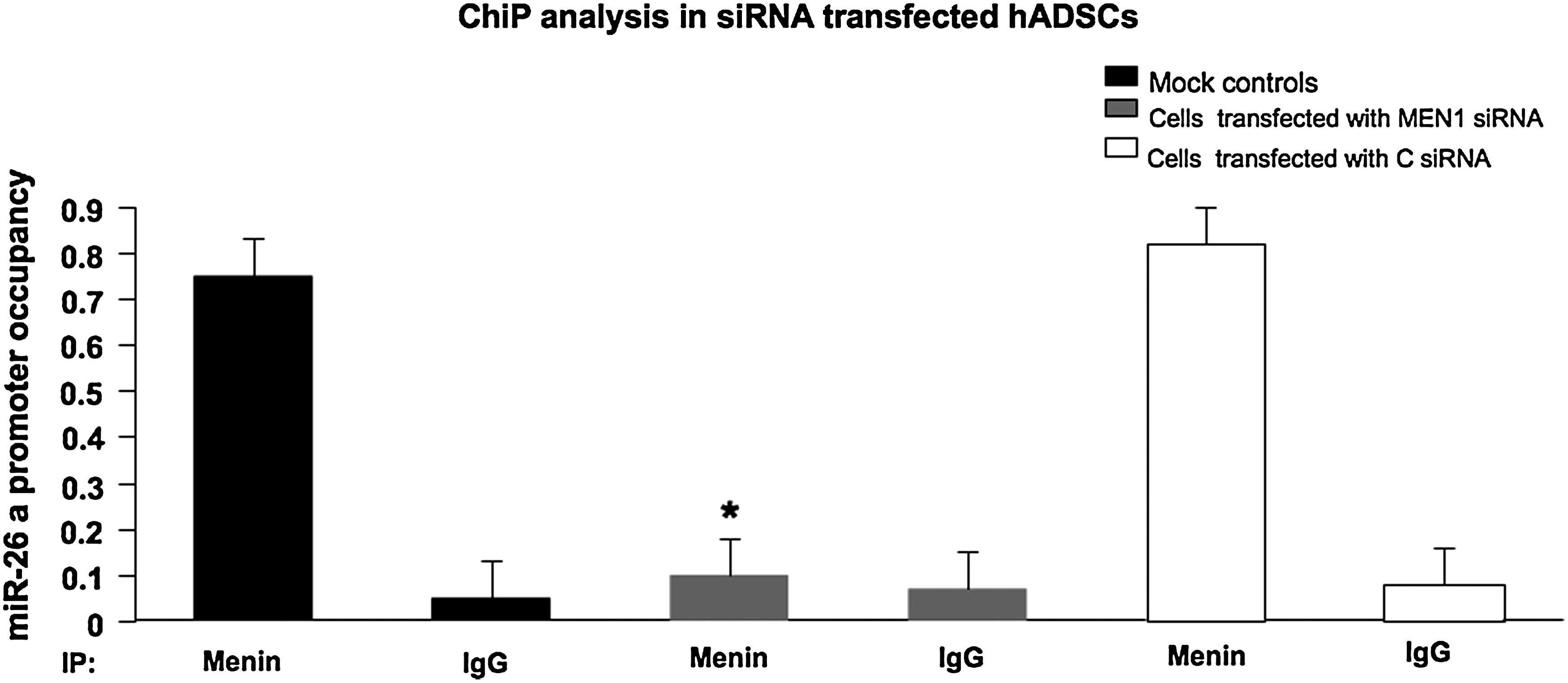
Chromatin immunoprecipitation (ChIP) analysis in hADSCs transfected with siRNAs. Chromatin was immunoprecipitated with anti-menin antibody, or with immunoglobin G (IgG) antibody as negative control. Quantitative SYBR green PCR was performed to determine whether the miR-26a promoter was present in the immunoprecipitate. Vertical bars indicate standard deviation. *=MEN1 siRNA vs. C siRNA and mock control (P<0.05; paired Student's t-test, n=3).
Discussion
Using osteoblast precursors obtained by subcutaneous human adipose tissue, the hADSC cells, induced to differentiate into osteoblastic lineage by dexamethasone we previously demonstrated that miR-26a expression was higher in differentiating osteoblasts, and that miR-26a inhibition induced osteoblastic differentiation through the increase of SMAD1 protein expression (Luzi et al., 2008), thus showing that SMAD1 is the direct target of miR-26a during the osteogenic differentiation of the hADSCs.
Menin play a central role in the regulation of gene transcription (Balogh et al., 2006) and in the regulation of osteoblast differentiation in C3H10T1/2 cells (Sowa et al., 2003); thus we hypothesize an interplay between miR-26a and menin in the regulation of SMAD1 mRNA and, consequently, in the regulation of the early osteoblastic differentiation.
The recent observations that the expression of a miRNA may be regulated by a TF, that TFs and miRNAs may reciprocally regulate themselves to form feedback loops, or that alternatively, both TFs and miRNAs may regulate their target genes and form FFLs (Saini et al., 2007), strengthen our hypothesis. We induced the down-regulation of MEN1 mRNA using a specific MEN1 siRNA in hADSCs committed to the osteoblastic differentiation. The effect of siRNA against MEN1 mRNA was the direct down-regulation of both menin protein and miR-26a expression, whereas no down-regulation of SMAD1 mRNA was observed. Conversely, we demonstrated an increase of SMAD1 protein expression, as expected in a miRNA post-transcriptional control. Thus, a decrease of menin level induced a decrease of miR-26 expression with a consequent loss of the negative control of miR-26a on SMAD1 mRNA 3′-UTR and an up-regulation of SMAD1 protein expression. This MEN1 modulation of miR-26a expression affected the expression of osteoblastic differentiation markers except for the RUNX2 gene, coding for a TF that interacts with SMAD1 for the activation of several osteogenic genes (Javed et al., 2008). BMP2 induced TFs constitute a network of activities and molecular switches for bone development and osteoblast differentiation. Among these regulators are RUNX2 is the principal osteogenic master gene for bone formation. Since our system is not induced by BMP2 but by dexamethasone, RUNX2 could follow an alternative pathway in our system. Chromatin Immunoprecipitation showed that menin occupies the miR-26a promoter region thus confirming that menin was acts as a TF that positively controls the expression of miR-26a.
The present findings indicate menin as a modulator of both miR-26a (directly) and SMAD1 protein (indirectly through miR-26a) expression in the osteoblastic differentiation pathway of hADSCs induced to differentiate with dexamethasone.
In conclusion, this is the first evidence that menin interacts directly with a miRNA, playing a pivotal role during the complex network of hADSCs osteogenic differentiation. This data opens also the possibility to develop RNA-antagomir-based potential therapeutic opportunities and challenges for improving the treatment of bone-related diseases by using miRNAs as therapeutic targets.
Footnotes
Acknowledgments
This work was supported by a grant of the Fondazione Ente Cassa di Risparmio di Firenze and F.I.R.M.O. Fondazione Raffaella Becagli to M.L. Brandi. F. Marini, Ph.D., was the recipient of the Amgen-Dompè fellowship, dedicated to Professor Isa Coghi from Osservatorio Nazionale Salute Donna (O.N.Da).
Author Disclosure Statement
All the authors declare that no competing financial interests exist.