
Abstract
Nucleic acid aptamers are short single-stranded DNA or RNA oligonucleotides that can bind to their targets with very high affinity and specificity, and are generally selected by a process referred to as systematic evolution of ligands by exponential enrichment. Conventional antibody-based therapeutic and diagnostic approach currently employed against biotoxins pose major limitations such as the requirement of a live animal for the in vivo enrichment of the antibody species, decreased stability, high production cost, and side effects. Aptamer technology is a viable alternative that can be used to combat these problems. Fully sequestered in vitro, aptamers eliminate the need for a living host. Furthermore, one of the key advantages of using aptamers instead of antibodies is that they can be selected against very weakly immunogenic and cytotoxic substances. In this review, we focus on nucleic acid aptamers developed against various biotoxins of plant, microorganism, or animal origin and show how these can be used in diagnostics (e.g., biosensors) and therapy.
Introduction
Aptamers are short single-stranded RNA or DNA oligonucleotides that can bind to a target with very high affinity and specificity (Ellington and Szostak, 1990; Tuerk and Gold, 1990; JAYASENA, 1999; Hermann and Patel, 2000; Nimjee et al., 2005; Famulok et al., 2007). In solution, the nucleotides form intramolecular interactions that fold the molecule into a complex 3-dimensional shape that allows the aptamer to bind tightly to the target molecule (Fig. 1). Aptamers can be developed for various targets with structures ranging from small molecules to complex proteins. Aptamer technology can be applied in diagnostics over therapeutics to nanotechnology. Aptamers are generated by a process referred to as systematic evolution of ligands by exponential enrichment (SELEX, Fig. 2) (Klug and Famulok, 1994; GOPINATH, 2007; Stoltenburg et al., 2007). Although aptamers serve a function similar to that of antibodies in molecular recognition, they do posses several advantages. One distinct advantage is that they can be chemically synthesized and modified to improve nuclease resistance and delivery efficacy. With a significantly low production cost compared with antibodies, aptamers can be selected and identified by screening very large libraries (ca 1015 unique members) with different conformations, and the selection can be optimized to generate aptamers that suit specific needs and applications. Due to their small size, aptamers can avoid immunological responses and show favorable biodistribution (Healy et al., 2004; Nimjee et al., 2005).
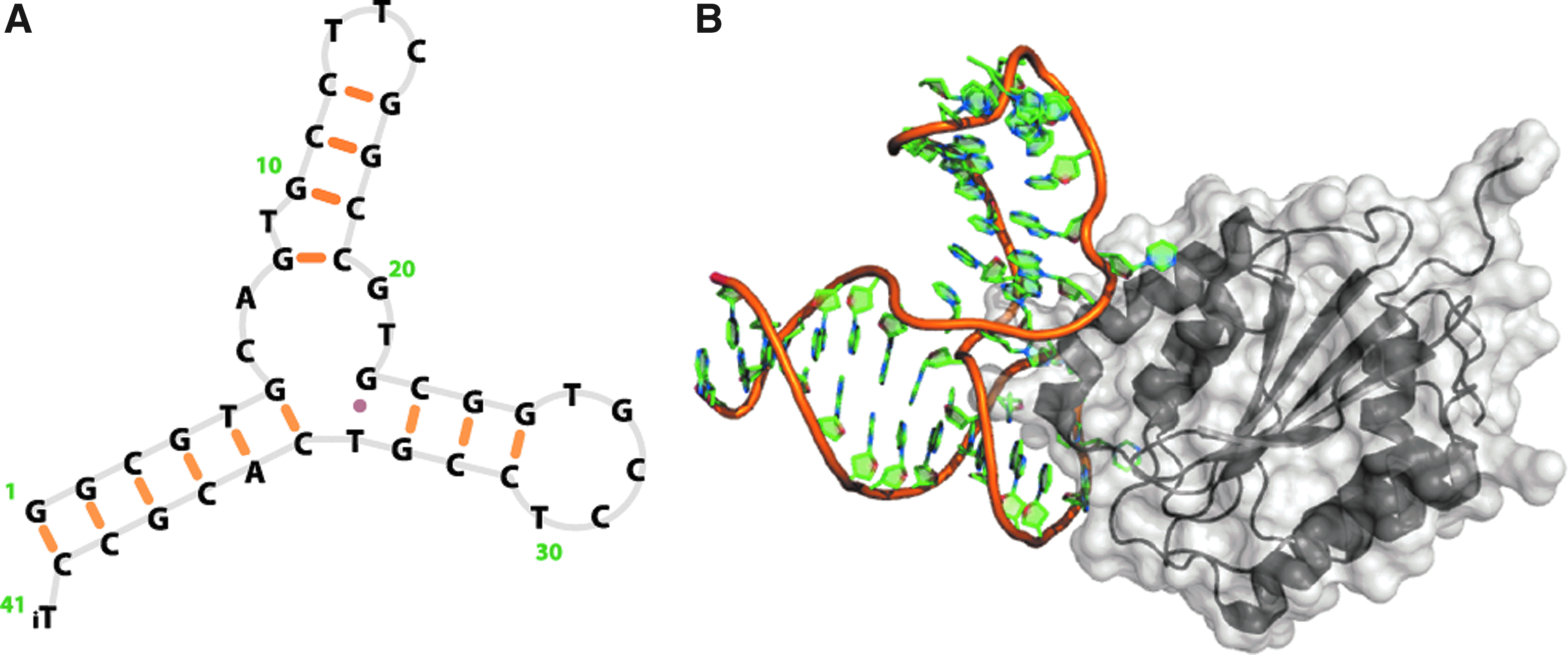
Structural representations of a nucleic acid aptamer bound to its specific target. The exemplified structure shows
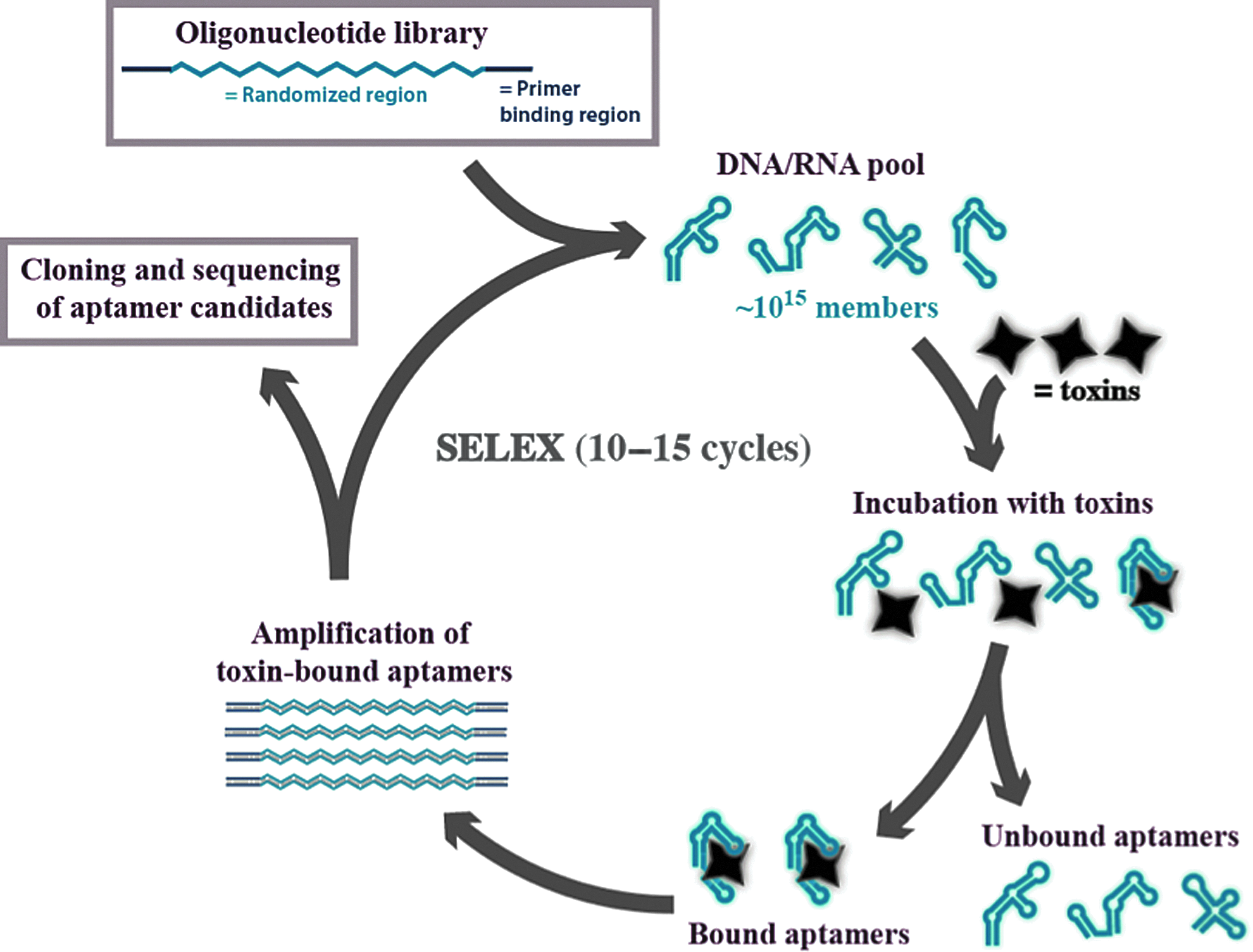
SELEX technology principle. A chemically synthesized combinatorial oligonucleotide library pool is subjected to incubation with a toxin target; the binding motifs are then separated and then enzymatically amplified and enriched. After 10–15 rounds of enrichment, the individual aptamers are identified by sequencing.
In this review, we focus on nucleic acid aptamers developed against various toxins of plant, microorganism, or animal origin and their usefulness in diagnostic and therapeutic development.
Bacterial Toxins
Botulinum neurotoxin
Clostridium botulinum produces one of the most potent neurotoxins known to man (Cartee and Monheit, 2011). Botulinum neurotoxins (BoNTs) are characterized by a heavy 100 kDa amino acid (aa) chain that is involved in the internalization of the toxin into neurons. The heavy chain (Hc) is linked by a disulphide bond to a 50 kDa light chain (Lc) zinc endopeptidase that cleaves soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) proteins synaptosomal-associated protein 25 (SNAP-25), syntaxin, and synaptobrevin. This prevents vesicle docking and acetylcholine release at the neuromuscular junction leading to severe flaccid respiratory paralysis. Although the toxin is widely used for treating dermal wrinkles, it also has a reported Please expand abbreviated term LD50 LD50 of around 1 ng/kg, making it one of the most potent bacterial toxins (GILL, 1982). Current treatment relies on respiratory support and treatment with antitoxin, a polyclonal mixture of antibodies isolated from a host injected with toxin. No treatments exist for the reversal of the flaccid muscle paralysis caused by BoNTs. The current diagnostic tools include mouse bioassay for sybtyping BoNTs, which can take several days, or immunaffinty assays that take up to several hours for obtaining a measurable readout (Ferreira et al., 2003). A mathematical modeling of the deliberate release of small amounts of BoNTs in a single milk-processing facility estimated that several hundred thousand casualties could be observed in a matter of days (Wein and Liu, 2005).
Tok and Fisher (Tok and Fischer, 2008) first reported a nucleic acid aptamer developed against Botulinum toxin. They used aldehyde-inactivated toxin and a short peptide fragment (aa 1177–1195) of the heavy chain in a single microbead-based selection assay. This method used only a single target-functionalized bead for the aptamer selection. The aptamers obtained by this approach bound the toxoid with low nanomolar affinities (Kd=3 nM) and the peptide fragment with low micromolar affinity (Kd=1.09 μM). The aptamers for the Hc fragment (aa 1177–1195) competed with commercially available antibodies for the same epitope. It was also argued that this approach yielded aptamers in a single round since there was no observed change in bulk Kd measured by fluorescence anisotropy (Tok and Fischer, 2008). Recently, the aptamers generated by Tok and Fisher have been used in a lab-on-a-chip device combining antibodies and aptamers in an electrochemical detection system. As the aptamer binds the BoNT toxoid, it folds up and exposes a fluorescein tag that is recognized by a fluorescein antibody conjugated to horseradish peroxidase. The measured signal allowed the detection of toxoid concentrations down to 40 pg/mL within minutes (Wei and Ho, 2009; Lillehoj et al., 2010; Wei et al., 2011). Later, Fan et al. published a novel method for selecting DNA aptamers against toxins. Using double-stranded DNA libraries of different lengths labeled with a quencher and a fluorophor, they selected aptamers against BoNT A light chain (Fan et al., 2008a). Very recently Bruno et al. performed a DNA SELEX experiment using BoNT A Lc funtionalized tosyl activated beads, and 60 DNA aptamer sequences were identified against BoNT subtype A light chain with binding affinities ranging from μM to nM (Bruno et al., 2012). Bruno et al. further performed an on-site quantification of BoNT A Lc using a Quantifluor™ handheld fluorometer. The reported detection limit was 1 ng/mL (Bruno et al., 2012), which was lower than the reported limit in the previously mentioned mathematical modelings by Wein and Liu (Wein and Liu, 2005).
To our knowledge no DNA aptamers have been evaluated for their therapeutic efficacy. However, the high binding affinities reported toward the light chain subdomain of BoNT indicate the potential for endopeptidase inhibition. On the other hand, using an automated selection protocol, 3 RNA aptamers were developed against Botulinum neurotoxin subtype A (Chang et al., 2010). The RNA aptamers selected against the light chain of BoNT A were able to bind and inhibit endopeptidase activity of Lc BoNT with dissociation constant (Kd) and half maximal inhibitory concentration (IC50) values in the low nanomolar range. Enzyme kinetics and in silico docking studies confirmed that one of the aptamers exhibited noncompetitive inhibition of function, while the remaining two showed noncompetitive inhibition of enzyme function (Chang et al., 2010).
Anthrax toxin
Anthrax is an acute infection caused by the spore-forming gram-positive bacterium Bacillus anthracis. The pathogenicity of the bacterium is mainly due to the secretion of a tripartite protein complex referred to as the anthrax toxin. The toxin consists of a lethal factor (LF), an edema factor (EF) and a cell-binding protein component called the protective antigen (PA). PA binds to cell membrane receptors and is sub-sequentially cleaved to form a 63-kDa active species (PA63) that causes heptamerisation of PA63/receptor complexes. This cell-membrane complex is able to bind the remaining 2 enzymatic components (LF and EF) of the anthrax toxin protein with very high binding affinity (1–2 nM) and initiate internalization. In the cytosol, EF catalyzes the conversion of adenosine-5′-triphosphate to cyclic adenosine monophosphate (cAMP,) changing the water homeostasis and inhibiting macrophage function, while LF proteolytically cleaves certain mitogen-activated protein kinases, leading to apoptosis (Young and Collier, 2007). Therefore, aptamers binding to any of these toxin components may disrupt the interactions leading to the onset of anthrax or could be used in diagnostic applications.
The first aptamer against PA was a 2′-fluoro-RNA aptamer developed by Archemix (Wilson et al., 2004). Nitrocellulose filter selection was used to enrich aptamers binding the c-terminal domain with Kd values of around 500 nM (Wilson et al., 2004). Another 4 aptamers were generated using ultrafiltration as the means of separation. These 30-mer DNA aptamers were generated against the 63-kDa fragment of PA. The PA63 aptamers had an apparent Kd vlaue of 1–120 nM depending affinity assay. The aptamers shared a common 8-mer consensus 5′-d(CCGTAAGG)-3′. When truncated, the 8-mer exhibited an apparent Kd value of 1.99 nM but failed to display specificity towards PA63 as the Kd value was equally low (high affinity) for bovine serum albumin (BSA). The structure of the 30-mer exhibiting the highest affinity showed no prominent secondary structure. In the same year, Oh et al. tested the efficacy of two DNA aptamers reported by Choi et al. by a fluorescence-based assay (Choi et al., 2011; Oh et al., 2011). The tested aptamers facilitated the detection of 1 nM PA.
Staphylococcal enterotoxin B
Staphylococcal enterotoxins (SE, 23 to 29 kDa) produced by Staphylococcus aureus bind the major histocompatibility complex class II receptors with a Kd of 10–100 nM and crosslink to human leukocyte antigens (HLA-DR) or HLA-DQ and T-cell receptors (Herman et al., 1991). The SE toxins are classified as super antigens because of the fact that even picomolar concentrations can induce nonspecific oligoclonal activation of T-cells and massive cytokine release, without having to be internalized and presented via normal immune response mechanisms (MISFELDT, 1990; Herman et al., 1991). This results in profound inflammation of the gastrointestinal tract leading to vomiting, nausea, and cramping, often within a few hours of exposure. In some cases, the infection with Staphylococcus aureus can lead to allergic and autoimmune response, even toxic shock syndrome with a previously reported mortality rate of over 50 % (Crass and Bergdoll, 1986). Staphylococcal enterotoxin B (SEB) has been investigated thoroughly as a biowarfare agent, especially in 1960s. (KRAKAUER, 1999). One incident reported that the exposure of 15 individuals while testing the aerosolized SEB on monkeys, which ultimately ended up hospitalizing nine people.
In 2002, Bruno and Kiel used a magnetic bead-based aptamer selection approach to evolve DNA aptamers against SEB (Bruno and Kiel, 2002). Using the double-stranded DNA pool generated after 5 rounds of SELEX, they could detect SEB down to less than 10 pg using electrochemiluminescence with Ru(bpy)32+. Using the enriched aptamer pool directly after in vitro selection for further assays is a rare procedure in aptamer development. The authors reasoned that this constituted the oligonucleotide equivalent of polyclonal antibodies used in antisera. We believe that this may be useful also in biosensors that normally use only one aptamer sequence, although several candidates that might have affinity to different regions on the same target are provided by the SELEX experiments. Purschke et al. reported a DNA aptamer (spiegelmer) against SEB (Purschke et al., 2003). The selection was conducted with a 25-mer truncation of the full-length (239 aa) SEB protein synthesized by solid-phase chemistry using only
Shiga toxin
Shiga toxin is produced by the Shigella dysenteriae toxin that shares many of the same pathological feats as the plant-derived ricin toxin (Spooner and Lord, 2012). Shiga toxin induced diarrhea-associated hemolytic uremic syndrome mainly affects the function of the kidneys and causes severe renal failure (Obrig and Karpman, 2012). Shiga toxins can also be found in some mutant cell lines of Escherichia coli, which posses a potential for extensive intoxication of humans due to their increased virulence (HUNT, 2010; Mora et al., 2011).
Fan et al. also reported an aptamer sequence for Shiga toxin. The Shiga toxin aptamer was used in a sensing system by employing double-stranded DNA labeled with a quencher and a fluorophor to detect Shiga toxin (Fan et al., 2008a). The sensitivity of the Shiga toxin aptamer was not tested but showed cross-reactivity with Bacillus anthracis spores and Francisella tularensis bacteria.
Cholera toxin
Cholera is caused by an infection with the toxigenic Vibrio cholerae. A life-threatening diarrheal disease is caused by both O1 and O139 serogroups of the bacterium producing the potent oligomeric protein named cholera toxin (Mandal et al., 2011). Cholera toxin causes the hypersecretion of electrolytes and fluid by interacting with intestinal mucosal cells and subsequent internalization resulting in elevated intracellular cAMP levels. The heightened levels of cAMP cause secretion of chloride and bicarbonate that draw water out into the small intestine (Mandal et al., 2011). Bruno and Kiel reported a DNA aptamer against cholera toxin in the same article about SEB aptamer development (Bruno and Kiel, 2002). By performing similar detection methodology as for SEB aptamers, they observed that the detection limit of the developed cholera toxin aptamer was between 10 and 40 ng (Bruno and Kiel, 2002). However, the sequence information of the developed aptamer was not disclosed in the article.
Mycotoxins
Ochratoxin A
Ochratoxin A (OTA) is a well-studied mycotoxin as a target for aptamer selection. In 2008, Cruz-Aguado and Penner reported the first aptamer-based mycotoxin detection system based on fluorescence polarization (Cruz-Aguado and Penner, 2008a). They selected a DNA aptamer that bound to OTA with a dissociation constant of 49 nM. Washing with different related compounds and wheat extract yielded an aptamer that bound specifically to OTA. The binding of the aptamer was closely associated with the concentration of divalent cations. Interestingly, changing 5 mM Mg2+ of the selection buffer to 10 mM Ca2+ resulted in a 4-fold increase in the affinity. Furthermore, the binding affinity did not change significantly with the complete removal of sodium or potassium ions, indicating no reliance on ionic strength of the assay solution (Cruz-Aguado and Penner, 2008a). The aptamer has been used to purify OTA from grain preparations (Cruz-Aguado and Penner, 2008b). Cruz-Aguado and Penner also reported that the aptamer was efficient enough to measure as low as 5 nM OTA using fluorescent polarization technique. The same aptamer has become a sort of gold-standard for many different OTA applications such as cyclic voltammetry-based detection assays (Kuang et al., 2010), electrochemiluminescent biosensor (Wang et al., 2010), colorimetric biosensors (Yang et al., 2011), fluorescent chromatographic strips (Wang et al., 2011a; Wang et al., 2011b), column extraction (Chapuis-Hugon et al., 2011), magnetic bead extraction (Wu et al., 2011b), electrochemical biosensors (Bonel et al., 2011), graphene biosensors (Sheng et al., 2011), luminescent nanoparticle assays (Wu et al., 2011b), impedimetric sensors (Prabhakar et al., 2011), exonuclease-assisted electrochemical detection (Tong et al., 2011), carbon nanotubes (Guo et al., 2011), fluorescence polarization (Kidd et al., 2011), DNAzyme biosensing (Yang et al., 2012), and rolling circle amplification aptasensing (Tong et al., 2012). Table 1 shows the listing of OTA aptamer applications and the achieved limit of detection (LOD) for all of the assays. The aptamers developed by Cruz-Aguado and Penner performed equally well as compared with the antibodies (Barthelmebs et al., 2011).
The table gives a comparative overview of biotoxin aptamers and biosensor performance from references cited in this review. (* indicates sequences not shown in reference-article.)
K d , dissociation constant; Do., ditto; LOD, limit of detection.
Fumonisin B1
Fumonisin B1 (FB1) produced by the fungus Fusarium verticillioides is a nephrotoxic, neurotoxic and hepatotoxic molecule that recently also has been attributed as a carcinogen in humans (Stockmann-Juvala and Savolainen, 2008). The toxin was first isolated and described from moldy corn. The fumonisins consist of 15 different subtypes grouped into 4 categories (A, B, C, and P), but the most prevalent of all fumonisins is B1. Fumonisin B1 aptamers have been developed by McKeague et al. in 2010 using FB1 immobilized magnetic beads. Eighteen rounds of SELEX provided aptamers that bound FB1 with affinities around 100 nM (McKeague et al., 2010).
Plant Toxins
Ricin toxin
Ricin is a potent toxin of the castor oil plant, which is isolated from the castor beans. A small dose of 500 μg is enough to kill an adult (Musshoff and Madea, 2009). Ricin toxin is a glycosylated heterodimer of 2 protein chains linked together with a disulfide bond. Ricin A chain (RTA) is a 32-kDa N-glycoside hydrolase that depurinates 28S RNA at position A4324. The position is called the sarcin-ricin loop and is important for the binding of elongation factors during protein synthesis. Hence, the loss of adenine moiety from position 4324 quickly and irreversibly abolishes ribosome function (Montanaro et al., 1973; Sperti et al., 1973). Ricin-filled metallic pellets discharged from the tip of an umbrella have been used as weapon in at least 8 assassinations attempts in history (Christopher et al., 1997). Although not always an effective toxin, the need for quick detection and therapeutic measures has signaled more research and development. Ricin sensing has been achieved in animal models (CLARKE, 1953), antibodies (Poli et al., 1994; Narang et al., 1997; Delehanty and Ligler, 2002; Shyu et al., 2002; Ligler et al., 2003) and by mass spectroscopy (Darby et al., 2001). The development of novel ricin toxin antibodies is still a growing field in biosensing.
Given the strong history and use as harmful agent, ricin has become the most investigated plant toxin for aptamer development. Ellington and co-workers first reported an RNA aptamer against the RTA domain (Hesselberth et al., 2000). The structure of an RNA aptamer reported by Hesselberth et al. has been used as a recognition tool in ricin-sensing applications (Hesselberth et al., 2000). Kirby et al. used another aptamer for its application in an electronic tongue array (Kirby et al., 2004). This assay consisted of a bead bound aptamer that captured ricin and was subsequently stained with anti-ricin antibody. This sandwich approach allowed for a LOD of 320 ng/mL. Remarkably, the beads could be stripped of ricin protein by denaturing aptamers with 7 M UREA buffer and reused for sensing (Kirby et al., 2004). The same group later published another setup in 2006 by utilizing glass slides coated with streptavidin or neutravidin to immobilize ricin aptamers. The assay was dependent on labeling of protein with a fluorescent dye and detected ricin in concentrations as low as 500 pg/μL (Cho et al., 2006). In 2006, Tang et al. developed two anti-ricin aptamers using two different selection approaches (Tang et al., 2006). They found that the 2 different methods provided high affinity binders with the best candidates having surprisingly high sequence similarity. Subsequent single molecule interaction studies have verified that the DNA aptamer had a slightly higher affinity, than the commercially available antibody (Wang et al., 2012). Superior in vivo stability was the key motivation for developing DNA aptamer instead of an RNA aptamer by Tang et al. However, Haes et al. demonstrated that a previously known RNA aptamer was able to sense ricin toxin down to 28 ng/μL in samples spiked with RNAse A (Haes et al., 2006).
In 2008, the RTA aptamer developed by the Hesselberth et al. was used in an in vivo study to test the inhibitory effect compared to the antibody against deglycosylated RTA (pRTA-IgG) (Fan et al., 2008b). They found that 200 ng/μL ricin aptamer reduced cell death from 80% to around 50%. The pRTA-IgG antibody generally performed better in the cell-death assays under the conditions used (Fan et al., 2008b). Single molecule recognition of ricin has also been achieved with the RNA aptamer dropping the detection limit to 100 fM. Interestingly, this unlabeled approach allowed measuring the actual interaction time between 2 ricin molecules binding to the same aptamer (i.e., time between binding, release, and rebinding). Ricin concentration was estimated from the interaction time as it was observed that the interval between first and second binding decreased from 35 to 2.6 minutes when ricin concentration was increased from 2.8 nM to 2.8 μM (Wang et al., 2012).
The structure of a DNA aptamer against ricin B-chain is recently published (Lamont et al., 2011). This aptamer was selected using tosyl-activated beads and after 8 rounds of selection, they identified that 93 out of 100 selected clones had the same sequence which is remarkable. The aptamer was used for sensing ricin toxin in juice and milk at a pH ranging from 2–7. In contrast, the commercially available enzyme-linked immunosorbent assay is performed only at neutral pH (Lamont et al., 2011).
Abrin toxin
Abrin is a class-2 ribosome inactivating protein. Abrin toxin uses the same pathogenic pathway as ricin toxin, however, ricin is 75 times more effective at depurinating A4324 (Dickers et al., 2003; OLSNES, 2004). Like ricin, abrin also possesses an enzymatically active A chain and a receptor-binding B chain. Abrin has been heavily investigated as a chemotherapeutic agent and also as a biological warfare agent. Tang et al. reported a DNA aptamer against abrin toxin (Tang et al., 2007). Since abrin features a galactose binding site, for aptamer development, it was immobilized on a galactose functionalized agarose beads. After 8 rounds of selection eight clones were sequenced. All 8 aptamer sequences had affinity to abrin in the low nanomolar range (28–130 nM). Using [Ru(phen)2(dppz)]2+ as an intercalator allowed studying the binding of abrin. One truncated version showed concentration-dependent reduction in chemiluminescence of [Ru(phen)2(dppz)]2+ upon binding. The assay facilitated the detection of a low amount of abrin toxin in complex serum samples and showed no cross-reactivity towards ricin or BSA (Tang et al., 2007).
Prospects of Biotoxin Aptamers
High levels of the above-mentioned biotoxins in humans can cause severe health problems that necessitate innovative detection technologies with high specificity and sensitivity. Table 1 highlights the development of aptamers against major biotoxins and their detection platforms. The existing traditional antibody-based methods are also selective and sensitive to some extent, however, there is still a need for simpler, more rapid and cost-effective approaches to produce the accurate results. Aptamer-based biosensor platforms generally show high sensitivities and specificities that could allow them to be competitive with existing detection methods. In Fig. 3, we summarized 4 efficient aptamer-based methods for rapid diagnosis of biotoxins. The performance of the developed aptamers under real-world conditions are yet to be seen although several of these existing aptamer platforms have been tested in various in vitro assays.

Conclusion
Nucleic acid aptamers can be viewed as molecular probes with a range of advantages that sometimes make them superior to traditional antibody-based technologies. In this report we have presented aptamers selected against naked toxins produced by living organisms. The presented aptamers are mainly used as biosensors for toxin detection, and their therapeutic efficacies have largely been unexplored. As aptamers are increasingly used in biosensor platforms and assays, we firmly believe that their therapeutic usefulness will also follow. Many of the aptamers presented herein will have direct applicability in vivo therapeutic assays and could potentially be used as therapeutic agents. With patent rights ending and more commercial ventures seeking to exploit the obvious advantages of aptamers as therapeutics and affinity ligands, aptamers will become increasingly present and hopefully live up to their predicted potential.
Footnotes
Acknowledgment
We greatly appreciate the funding from the University of Queensland (UQ fellowship and UQ ECR Grant schemes awarded to R.N.V.). L.H.L. acknowledges funding from the Novo Nordisk Foundation to support his stay at The University of Queensland, Brisbane.
Author Disclosure Statement
No competing financial interests exist.