
Abstract
RNA interference (RNAi) has been proven in recent years to be a newly advanced and powerful tool for development of therapeutic agents toward various unmet medical needs such as cancer, in particular, a great attention has been paid to the development of antineoplastic agents. Recent success in clinical trials related to RNAi-based therapeutics on cancer and ocular disease has validated that small interfering RNAs (siRNAs) constitute a new promising class of therapeutics. Currently, a great wealth of multi-target based siRNA structural modifications is available for promoting siRNA-mediated gene silencing with low side effects. Here, the latest developments in RNAi-based therapeutics and novel structural modifications described for siRNAs—in particular multi-target siRNAs—are reviewed.
Introduction
R
RNAi-Based Therapeutics
RNAi technology used to suppress expression of pathologically and physiologically important genes is applicable for the treatment of human diseases caused by specific genes that are difficult to treat through traditional approaches. Since the first description of RNAi phenomenon in mammalians, development of RNAi-based therapies has rapidly been advanced with many powerful new drug candidates in clinical trials against various human diseases, such as age-related macular degeneration (AMD) (McFarland et al., 2004; Mccullough et al., 2005; Gu et al., 2010), respiratory syncytial virus (RSV) infection (Bitko et al., 2005), neurodegenerative disorders (Raoul et al., 2006) and various cancers (Takeshita and Ochiya, 2006; AIGNER, 2007; Davidson and McCray, 2011).
Candidates of RNAi-based drugs
Increasing popularity in the development of RNAi-based therapeutics over recent years has evidenced many siRNA preclinical and clinical trials, currently with about 30 siRNA therapeutic candidates reaching various clinical trials stages for treatment of at least 16 diseases (DeVincenzo et al., 2008; Davis et al., 2010; Leachman et al., 2010; Burnett et al., 2011; Davidson and McCray, 2011; Burnett and Rossi, 2012). Some major global pharmaceutical companies such as Novartis and Merck have also been involved in clinical applications of RNAi-based therapeutics. RNAi is considered to be an exceptional technology for knock-down or silencing of disease related target genes, and scientists anticipate that it will significantly shorten the drug development timeline. Furthermore, clinical pipelines of RNAi-based therapies using siRNAs have been gradually growing since approximately 2011 as the technology matures. As shown in Table 1, there are many candidates of RNAi-based drugs under clinical development in 2013.
ADRB2, adrenergic receptor beta-2; AKI, acute kidney injury; AMD, age-related macular degeneration; APOB, apolipoprotein B; ATTR, amyloidosis; BCR-ABL, breakpoint cluster region-abelson murine leukemia; CML, chronic myeloid leukemia; DGF, delayed graft function; DME, diabetic macular edema; DOD, Department of Defense; DOPCC, 1,2-dioleoyl-sn-glycero-3-phosphocholine; EPHA2, ephrin type-A receptor 2; EUS, endoscopic ultrasound; GM-CSF, granulocyte-macrophage colony-stimulating factor; HIF-1A, hypoxia-inducible factor 1-alpha; HMO, Hadassah Medical Organization; HSP47, heat shock protein 47; IPCC, International Pachyonychia Congenita Consortium; iPsiRNA, inducible Proteasomal subunit-specific siRNA; IV, intravenous; IVT, intravitreal; KRASG12D, K-Ras mutation G12D; KSP, kinesin spindle protein; LMP2, large multifunctional protease 2; LMP7, large multifunctional protease 7; LNA, locked nucleic acid; LODER, local drug eluter; MECL1, multicatalytic endopeptidase complex-like 1; NCI, National Cancer Institute; NDT, Nitto Denko Corporation; OCRF, Ovarian Cancer Research Fund; PCSK9, proprotein convertase subtilisin/kexin type 9; PKN3, protein kinase N3; PLK1, polo-like kinase 1; RONDEL, RNA/oligonucleotide nanoparticle delivery; RRM2, ribonucleoside-diphosphate reductase subunit M2; RSV, respiratory syncytial virus; siRNA, small interfering RNA; SNALP, stable nucleic acid lipid particle; TRPV1, transient receptor potential vanilloid 1; TTR, transthyretin; UT MDA, M.D. Anderson Cancer Center; VA-lip, vitamin A–coupled liposome; VEGF, vascular endothelial growth factor; VEGF-R1, vascular endothelial growth factor receptor 1.
RNA delivery systems
RNAi-based drugs rely on safe and efficient delivery systems without unwanted side effects. Up to now, there are a number of efficient delivery systems which can be categorized as ex vivo, topical/local and systemic methods (Peer D and Lieberman, 2011; Vicentini et al., 2013). Local delivery of siRNA is practicable for tissues that are external and/or locally restricted including ocular, epidermal, pulmonary, and bladder. Additionally, local delivery may be suitable for noninvasive therapies that require administration by patients, such as eye drops and nasal sprays. The treatment of AMD and diabetic macular edema using intravitreal injections were the first clinical applications of siRNAs as these drugs could be delivered directly to ocular tissue (Barakat and Kaiser, 2009; Kaiser et al., 2010). Other therapeutic applications using siRNAs have been developed via topical delivery and are currently in clinical trials (QPI-1007, TD101, SYL040012, SYL1001, ALN-RSV01). In the past 5 years, the development pipeline has shifted from local to systemic delivery as more advanced delivery vehicles become available for systemic applications, including stable nucleic acid lipid particles (SNALP) (Zimmermann et al., 2006) and RNA/oligonucleotide nanoparticle delivery (RONDEL) (Heidel et al., 2007; Davis et al., 2010). These technologies have demonstrated to be effective in vivo (Zimmermann et al., 2006; Judge et al., 2009; Davis et al., 2010), and significant progress is being achieved in some clinical trials (PRO-040201, TKM-080301, ALN-TTR01, ALN-PCS02, ALN-VSP02, CALAA-01) (Table 1).
Structural Modifications of siRNA
To improve the silencing efficiency of siRNAs, researchers tried different ways to modify structures of siRNAs. A variety of structural siRNA modifications has been reported for gene silencing (Table 2).
Bold line: sense strand, thin line: antisense strand.
aiRNA, asymmetrical siRNA; dgRNA, double-guide siRNAs; shRNA, short hairpin RNA; sisiRNA, small internally segmented interfering RNA; ss-siRNA, single-stranded siRNA.
Shorter siRNAs mimicking Dicer cleavage products
The structure of most canonical siRNAs mimics features of the products from Dicer natural cleavage, which comprises two annealed strands, one is a 21-nucleotide (nt) guiding strand antisense to a target RNA and the other is a complementary passenger strand. These two strands form an siRNA duplex upon annealing with a 19-bp dsRNA stem and 2-nt 3′-overhangs at both ends (Elbashir et al., 2001b). Depending on sequence features, some studies reported that siRNAs shorter than canonical siRNAs could efficiently trigger RNAi in mammalian cells and sometimes led to better silencing effect even than that of canonical siRNAs. Reduction of nonspecific responses and immune stimulation as well as enhanced internalization with shorter siRNAs has also been reported. Some variations in the shorter modification on siRNAs include blunt 19-bp siRNA (Czauderna et al., 2003; Prakash et al., 2005; Hogrefe et al., 2006; Ghosh et al., 2009), short hairpin siRNA (Siolas et al., 2005; Ge et al., 2010), 16-mer siRNA (Chu and RANA, 2008), asymmetrical shorter-duplex siRNA (Sun et al., 2008), and asymmetrical siRNA (Sun et al., 2008). Other modified siRNAs used in research and drug development are single-stranded siRNAs (Martinez et al., 2002; Holen et al., 2003; Hall et al., 2006), double-guide siRNAs (Hossbach et al., 2006), small internally segmented interfering RNAs (sisiRNA) (Bramsen et al., 2007), fork siRNAs (HOHJOH, 2004; Petrova Kruglova et al., 2010), dumbbell-shaped circular siRNA (or dumbbell siRNA) (Abe et al., 2007), and bulge-siRNA (Dua et al., 2011).
Long dsRNA designed as Dicer-substrate for gene silencing
RNAi effectors longer than standard siRNA were reported in 2005 by Kim et al. and Hannon et al. These longer siRNAs that structurally mimic various Dicer substrates to enhance incorporation into RNAi pathways, are termed “Dicer-substrate siRNA” (Kim et al., 2005; Rose et al., 2005; Siolas et al., 2005; Amarzguioui et al., 2006; Collingwood et al., 2008; Hefner et al., 2008; Tanudji et al., 2009). An investigation of cellular interferon induction caused by in vitro transcribed siRNAs showed that limited concentrations of some 25- to 27-mer siRNAs had greater potency than all of the possible synthetic 19-bp duplex and 2-base 3′-overhangs siRNAs that could be generated from the larger duplex (Kim et al., 2005). Hannon et al. (2005) also found that synthetic dsRNA with 25- to 30-mer lengths could be up to 100-fold more potent than corresponding conventional 19-bp duplex and 2-base 3′-overhangs siRNAs and would not activate protein kinase R or induce interferon (IFN). The enhanced potency of the longer duplexes is attributed to the fact that they are substrates of Dicer, linking the production of siRNAs to incorporation in the RISC. Foster et al. (2012) evaluated the in vitro and in vivo activities of siRNAs and dual-target siRNAs (dsiRNAs) targeting phosphatase and tensin homolog and factor 7 with over 250 compounds representing both siRNA and dsiRNA structures. They identified highly active compounds from 25-/27-mer dsiRNAs both in vitro and in vivo (Foster et al., 2012). This study further demonstrated that in vitro processing by Dicer is direct, starting predominantly from the open end of the stem and generating a mixture of 19-bp duplex and 2-base 3′-overhangs cleavage products. Increased potency could be attributed to Dicer processing, which might promote more efficient incorporation into RISC through physical association of Dicer with Ago proteins, the effectors of RNAi. This interpretation is supported by biochemical evidence in Drosophila melanogaster, indicating a role for Dicer in the initial stages of RISC assembly (Pham et al., 2004).
Although structurally modified siRNAs have been utilized successfully, more investigations and development may be required to further validate such modified and functionalized siRNAs as therapeutic agents. Some of these siRNAs could be loaded directly into RISC whereas others serve as substrates for Dicer and can be processed into shorter species before loading into the RISC. Doré-Savard et al. (2008) demonstrated that a low dose of 27-mer Dicer-substrate siRNA resulted in effective silence of target genes that are related to central nervous system disorders.
Multi-targeting siRNAs for Therapeutics
Drugs designed to act on individual molecular targets cannot usually address multigenic or multi-factor diseases such as cancer, or diseases that affect multiple tissues or cell types such as diabetes and immune-inflammatory disorders (Zimmermann et al., 2007). Thus, multi-targeting therapeutics has been considered to be an attractive approach toward complex diseases. Such an approach as addressing multiple targets has been used to regulate multiple nodes of disease network resulting in a synergistic effect (Bolognesi et al., 2009; Efferth and Koch, 2011) and to design medications against atherosclerosis (Lahoute et al., 2011), cancer (Lu et al., 2012), depression (Maes et al., 2012), and psychosis (Kroken et al., 2014) as well as neurodegenerative diseases (FLIGHT, 2013). Another approach focused on “systemic” drugs suggested that development of novel computational and mathematical concepts be certainly required for suitable modeling of complex data (Schrattenholz et al., 2010).
Combination of multi-target siRNAs for disease therapeutics
Various design strategies with multiple-target siRNAs have been used in many gene-mediated disorders or viral infectious research (SHARP, 2001; Holen et al., 2002). Use of multiple siRNA duplexes in targeting CXC chemokine receptor type 4 (CXCR4) and fatty acid synthase ligand (FASL) was first reported to prevent human immunodeficiency virus-1 (HIV-1) infection. An enhanced gene silencing with the multiple siRNAs was demonstrated not only at protein or mRNA expression level, but also at functional level by evoking greater inhibition of HIV infection as well as more reduction of FASL-mediated apoptosis than that using single siRNA (Ji et al., 2003). Menendez et al. (2004) identified a molecular link between fatty acid synthase (FAS) and human epidermal growth factor receptor 2 (HER2) oncogene by using concurrent RNA-mediated silencing of FAS and HER2 genes, they found that simultaneous inhibition of FAS and HER2 genes by corresponding siRNAs led to apoptosis synergistically in HER2 overexpressing cancer cells. In an anti-prostate cancer study, multi-target siRNAs based on the homologous region of the DNA methyl transferases 3A and 3B (DNMT3A/B) family were designed and their effects on proliferation, migration, and invasion of TSU-PR1 prostate cancer cells were investigated in vitro (Du et al., 2012). The results showed that DNMT3B alone apparently played a key role in maintaining unfavorable behavior of prostate cancer cells, thereby implying the potential significance of DNMT3B as a promising therapeutic target, while DNMT3A was simply a helper. To evaluate the effect of dual gene silencing from heat shock protein 27 (HSP27) and cellular Fas-associated death domain protein (FADD)-like interleukin-1β-converting enzyme inhibitory protein (C-FLIP) on doxazosin-induced apoptosis of prostate cancer cell, Kim et al. (2013) used a mixture of siRNAs corresponding to HSP27 and C-FLIP. They showed that dual silencing of C-FLIP and HSP27 enhanced apoptosis even under 1 μM—a rather low concentration—of doxazosin in prostate cancer PC-3 cells. In another study (Tai et al., 2010), dual silencing of HER2 and vascular endothelial growth factor (VEGF) genes by multi-target siRNAs exhibited significant changes in cell morphology and substantial suppression on migration, spreading, cell adhesion, and proliferation in vitro breast cancer model. These results suggested that HER2 positive breast cancer could be treated more effectively by inhibition of both HER2 and VEGF genes simultaneously. Also in gastric cancer, Liu et al. (2014) demonstrated that co-downregulation of VEGF and HER2 by a siRNA cocktail resulted in significant inhibition of gastric cancer growth and migration in vitro. Thus, inhibition of VEGF and HER2 expression could induce apoptosis of gastric cancer cells. Other examples of applying combinations or cocktails of siRNAs for multiple genes targeting are presented in Table 3. These investigations showed that two or more siRNAs could be synthesized and mixed for therapeutic purposes.
BCL-X (L), B-cell lymphoma-extra large; C-FLIP, cellular Fas-associated death domain protein-like interleukin-1β-converting enzyme inhibitory protein; CXCR4, CXC chemokine receptor type 4; DNMT3A, DNA methyltransferases 3A; DNMT3B, DNA methyltransferases 3B; EREG, epiregulin; EZH2, enhancer of zeste homolog 2; FAK, focal adhesion kinase; FAS, fatty acid synthase; FAS-ligand (FASL), FAS ligand; HER2, human epidermalgrowth factor receptor-2; HIV, human immunodeficiency virus; HSP27, heat shock protein 47; IP, intraperitoneal; IT, intratumoral; MDM2, mouse double minute 2 homolog; P110α, phosphatidylinositol-4,5-bisphosphate 3-kinase, catalytic subunit alpha; XIAP, X-linked inhibitor of apoptosis protein).
Dicer substrate-based designs for multi-target siRNAs
Alternatively to the above modification methods, conventional double-stranded siRNAs can be structurally modified to bear multiple targeting RNA sequences. In a number of studies, such siRNAs with quite unique structures have been designed and used as Dicer substrates for therapeutic evaluations (Table 4).
▵, point mutations in sense (mhRNA) or antisense strand (e-shRNAs).
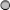 , DTME, a cleavable crosslinker.
, DTME, a cleavable crosslinker.
○, dT or RNA, DNA nucleosides.
DBP, D site of albumin promoter (albumin D-box) binding protein; ddRNAi, DNA-directed RNAi; DGT siRNA, dual gene targeting siRNA; DNA, deoxyribonucleic acid; dsiRNA, dual-target siRNA; dT, deoxythymidine; DTME, dithiobismaleimidoethane; EGFP, enhanced green fluorescent protein; e-shRNAs, extended short hairpin RNAs; hpRNA, hairpin RNA; lhRNA, long hairpin siRNA; mhRNA, modified hairpin RNA; multi-siRNA, multimeric siRNA; RNA, ribonucleic acid; tiRNA, tripartite-interfering RNA; tsiRNA, tandem siRNA; T-tiRNAs, tripodal interfering RNAs.
Short hairpin siRNAs
Anderson et al. (2003) reported that bispecific short hairpin siRNAs containing an 8-nucleotide intervening spacer targeted against either CXCR4 and cluster of differentiation 4 or C-C chemokine receptor type 5 (CCR5) and CXCR4, which are known HIV-1 cell surface receptors. After being transfected with the bispecific siRNAs, MAGI-CXCR4 and CCR5 cells exhibited significant downregulation of their respective coreceptors. In other approach, a double siRNA structure with a single construct called hairpin RNA (hpRNA)—produced by a cytomegalovirus promoter-driven DNA template—was used to block expression of matrix metalloproteases (MMP)-9 and cathepsin B genes (Lakka et al., 2004). The hpRNAs were found to reduce the invasive behavior of glioblastoma cell line, which was attributed to the inhibition of the MMP-9 and cathepsin B expression. Direct intratumoral (IT) injections of plasmid DNA expressing hpRNA inhibited growth of established glioma tumor model significantly and reduced invasion of intracranial tumors in in vivo models. On the other hand, intraperitoneal (IP) injections of plasmid DNA expressing hpRNA not only repressed preestablished tumors greatly without any indication of tumor cells present but also kept its effect for a period of 4 months. Gondi et al. (2004) showed that, in both in vitro and in vivo models, glioma cell invasion and angiogenesis could be inhibited when an hpRNA targeting the receptor of urokinase-type plasminogen activator (UPAR) and cathepsin B was used, IT injections of plasmid vectors expressing the hpRNA for UPAR and cathepsin B resulted in the suppression of preestablished intracranial tumors. Other hpRNAs designed for targeting UPAR, MMP-9, and UPAR were also tested for treatment of meningioma (Tummalapalli et al., 2007; Kargiotis et al., 2008) and breast cancer (Kunigal et al., 2007). All these studies showed satisfactory inhibition effects of multi-target hpRNAs on cancers in vivo.
Long hairpin siRNAs
Long siRNAs with hairpins for multiple gene targets were designed to achieve sustained gene silencing. DNA-directed RNAi (ddRNAi) technology, by which long siRNA with hairpin transcripts were expressed after introduction of ddRNA constructs into cells, had recently been developed (Rice et al., 2005) and successfully applied for anti–hepatitis-C (HCV) therapeutics in a clinical trial (Denise et al., 2014). Others also reported various designs of long siRNAs with hairpin structures (Akashi et al., 2005; Liu et al., 2007; Sano et al., 2008). Novel modified hairpin RNAs (mhRNAs) more than 100 bp in length were designed (Akashi et al., 2005) and featured by multiple specific point mutations in the sense strand. Such mhRNAs induced RNAi and suppressed replication of multiple HCVs effectively without inducing the IFN pathway. Liu et al. (2007) introduced the so-called extended short hairpin RNAs (e-shRNAs) that encode two effective siRNAs against HIV-1 variants in a construct. The study indicated that, from activity assays and RNA processing analyses, positioning of the two siRNAs on the hairpin stem was critical to generation of the two corresponding functional siRNAs. Efficient processing of the e-shRNAs into two effective siRNAs could be achieved, which led to better inhibition of virus reproduction. A similar form of long hairpin siRNAs (lhRNAs), termed promoter-expressed long hairpin RNAs, was evaluated (Sano et al., 2008). Long hairpin RNA could be processed into multiple siRNAs that are deemed as effective agents for treatment of rapidly mutating viruses such as HIV. Human U6 promoter-driven lhRNAs of 50, 53, and 80 bp targeting contiguous sequences within the tat and rev genes of HIV-1 have been generated and the results showed that such lhRNAs could be stably and functionally expressed for a long time (48 days) in HIV-1 susceptible T cells, where they provided potent inhibition of HIV replication against both nonmutant and mutant variants of HIV-1.
With rapid advancement in chemical synthesis and modification methods, chemically synthetized siRNAs other than vector-based siRNAs have been widely used for investigations of RNAi-based therapeutics. In addition, synthetic multi-target siRNAs could also be conjugated with other moieties by linkages. In one case, Lee et al. (2011) reported that use of conjugates formed from dual gene targeting multimeric siRNA (DGT multi-siRNA) provided more effective inhibition of the two corresponding target genes simultaneously than that from a cocktail of mixed multimerized siRNA conjugates. The DGT multi-siRNA conjugates, which were chemically cross-linked via cleavable disulfide linkages, exhibited significantly higher gene silencing efficiency with negligible stimulation of immune response against VEGF and green fluorescent protein (GFP) genes at expression levels of both mRNAs and proteins than a physical mixture of naked siRNAs. DGT multi-siRNAs against SURVIVIN and BCL2 genes were shown to induce enhanced apoptotic effect in a similar fashion.
Branched siRNAs
Branded siRNAs are another class of novel structural designs that could be obtained by using symmetric doubler phosphoramidites (Frieden et al., 2004) to form branches with two or four strands from RNAs. The branched siRNAs had similar inhibitory capacity to those of unmodified siRNA duplexes, as evidenced by tumor necrosis factor-α (TNF-α) gene silencing experiments (Aviñó et al., 2011). A variant of branched siRNAs, called tripodal interfering RNAs (T-tiRNAs) (Chang et al., 2012a), was constructed on a trebler-phosphoramidite core structure (Shchepinov et al., 1997) and extended with short DNA linker. Such T-tiRNAs could silence simultaneously up to three different mRNAs or miRNAs by harboring three siRNA or antagomir units. T-tiRNAs could be incorporated into RISC efficiently and trigger efficient RNAi in mammalian cells and achieve greater gene silencing and delivery efficiency than conventional siRNAs. There were also other branched RNAs with three- or four-way junctions (Nakashima et al., 2011) designed by assembling single-stranded RNA for RNA interference. These unique RNAs could be cleaved by Dicer as well into about 20-bp conventional siRNA species that lead to a potent silencing effect. Thus, a branched, nonlinear tripartite-interfering RNA (tiRNA) structure was designed by Chang et al. (2012b), and the study showed that the tiRNA could induce better silencing potency on multiple target genes than the corresponding classical siRNAs. Surprisingly, the gene silencing observed on the use of tiRNA did not require Dicer-mediated processing into smaller RNA units and was thought to be triggered by the 38-nt-long guide strands through the RNAi machinery in mammalian cells.
Linear multi-target siRNAs
In contrast to the above hairpin and branched structures of siRNAs, linear multi-target siRNAs have also been designed and evaluated for silencing of various gene targets. Tandem siRNA (tsiRNA) (Shin et al., 2009), which was composed by a duplex RNA harboring two siRNA units by using an expression system with convergent H1 and U6 polymerase 3 promoters, was used to target HCV (Lavender et al., 2012; Suhy et al., 2012). TsiRNAs were optimized to be processed by the intracellular ribonuclease Dicer into functional siRNAs targeting different sequences in a given gene or different genes. In vitro, inhibition of HCV protein could be achieved by tsiRNAs as efficiently as a single 19-bp duplex and 2-base 3′-overhangs siRNA without affecting miRNA maturation or induction of an IFN response. Chang et al. (2009) showed that synthetic dsiRNAs with a length as long as 38 bp could induce specific silencing of two target genes simultaneously without causing nonspecific antiviral IFN responses. Long double-stranded dual-target or multiplex siRNAs (DGT-siRNAs or multi-siRNAs) (Wu et al., 2013; Peng et al., 2013) were reported for gene silencing through a sequence-specific RNAi process without inducing significant immune responses. A gap feature structurally designed in either of the nucleotide strands of the multi-siRNAs was essential toward silencing target genes and avoiding immune responses.
Conclusion and Perspectives
Here, we have summarized the current states of RNAi therapeutics and development of siRNA structural modifications, in particular multi-target siRNAs. By using these structurally modified siRNAs, gene silencing efficiency has been improved with low or even no induction of innate immune responses. Some of them can even enhance delivery efficiency compared with canonical siRNA. With rapid growth of RNAi therapy applications, many RNAi-based drug candidates have moved into clinical trials in recent years. Although significant progress has been made toward delivering siRNAs for therapeutic applications as evidenced by the clinical trials, much more effort is still needed to ensure safe and efficient means of delivery of siRNAs in vivo. siRNA agents have been successfully delivered into many organs in clinical trial studies, such as eyeball, skin, liver, and lung which may be easier targets than solid tumors in internal organs.
With better understanding of disease mechanisms, novel multi-target drugs have been developed against multiple sites within the pathological process. The RNAi-based therapeutics has also been investigated for multi-target therapies by many researchers. A number of applications using multi-target siRNAs for antiviral and antitumor therapeutics have achieved substantial efficacy and safety outcomes. Some of them have moved into the clinic trials (Table 1). Multi-target-based siRNA therapeutics opens up new therapeutic possibilities and may help to improve treatment effectiveness. Additionally, single- or multi-target siRNA could be used as a pharmaceutical ingredient by combining with other biomedical and/or chemical agents such as monoclonal antibody drugs and/or chemotherapy agents for diseases treatments.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Acknowledgments
This work was supported by Foundation of the Production–Study Research Prospective Joint Research Programs of Jiangsu Province, China (2013042-06), the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), the Science Foundation of Nantong City, Jiangsu Province, China (HS2012070), the Postgraduation Advanced Project Program of Nantong University (2012-29 and 22292679), the National Science and Technology Major Project from the National Natural Science Foundation of China (2011ZX09401-012), and the National High Technology Research and Development Program of China (863 Program) (2012AA022501).