
Abstract
The species sensitivity and mechanism of complement pathway activation by a phosphorothioate oligonucleotide were investigated in monkey and human serum. Increasing concentrations of a phosphorothioate oligonucleotide, ISIS 2302, were incubated in either monkey or human serum. Complement activation in monkey serum was selective for the alternative pathway and occurred at concentrations ≥50 μg/mL ISIS 2302. By comparison, complement activation in human serum was absent. A similar difference in sensitivity for activation was also observed for a representative 2′-methoxyethyl (MOE)–modified oligonucleotide. The absence of oligonucleotide-induced complement activation was also observed in dogs. Protein binding with ISIS 2302 and enzyme competition studies suggested that factor H was important in oligonucleotide-mediated complement activation process, and addition of factor H to serum effectively prevented the activation in monkey serum. Furthermore, based on the immunoassay for factor H, there was an apparent decrease in factor H concentration as the ISIS 2302 concentration increased. This result suggests that ISIS 2302 binds to factor H and interferes with the factor H antibody from the immunoassay. Factor H is a regulatory protein that limits alternative pathway activation. Disruption of factor H interaction with C3 convertase by oligonucleotide could promote activation in this pathway.
Introduction
P
Despite the specificity derived from their base-pairing properties, these compounds do have the potential to produce toxicities through hybridization-independent mechanisms [4,5]. Phosphorothioate oligonucleotides are stable, water-soluble polyanions that are generally 6,800 to 7,200 Daltons. These physical properties favor protein interactions and promote a number of beneficial properties of phosphorothioate oligonucleotides such as favorable pharmacokinetic profiles and tissue distribution [6,7]. However, oligonucleotide–protein interactions may also lead to toxicity or unintended physiologic effect. Since these protein interactions are dependent on the biophysical properties of the oligonucleotides, rather than the specific nucleotide sequence, they typically produce similar effects from one sequence to another [5].
Hybridization-independent effects of oligonucleotides are thought to occur through interactions with soluble proteins leading to either altered enzyme functions or cell binding that may result in cellular activation. Sequence-independent toxicities observed in monkeys resulting from oligonucleotide interactions with specific plasma proteins include prolongation of clotting times and activation of complement [8,9]. These effects on serum enzymatic cascades are reversible and transient due to the rapid clearance of oligonucleotide from blood. As oligonucleotide is cleared from blood, normal function of these serum proteases returns.
Complement activation by phosphorothioate oligonucleotides shared properties similar to the effects on the coagulation cascade as described above. Complement activation in monkeys was also sequence independent, the effect correlated with elevated plasma concentration of oligonucleotide, and the activation was transient [8,10]. Existing data demonstrate a threshold concentration of 70 to 80 μg/mL total oligonucleotide after a short-term 1- or 2-hour intravenous infusion (i.v.) is required for activation [8]. However, a more complete understanding of the molecular interactions responsible for complement activation and a more thorough understanding of the interspecies sensitivity to these compounds is needed. It appears that interaction between the oligonucleotides and certain components of the alternative pathway disrupted the normal regulation process. While complement activation by oligonucleotides has been confirmed in monkeys, a number of other species have been thoroughly studied with no physiologic evidence that complement activation by oligonucleotides occurs [11–13]. There are currently no data to explain the basis for these species differences.
An in vitro model for complement activation is presented and was used to elucidate the mechanism of oligonucleotide-induced complement activation. The in vitro assay provides a basis for comparison of complement activation by phosphorothioate oligonucleotides between monkey and man. Data are also presented to highlight differences between species and sensitivity to complement activation by oligonucleotides.
Materials and Methods
Oligonucleotides
Oligonucleotides used in this study were synthesized at Isis Pharmaceuticals, Inc. using a Milligen model 8800 DNA synthesizer using solid-phase triester chemistry as described in the literature [14]. The phosphorothioate internucleotide linkage was generated by use of Beaucage reagent as the P(III)→P(V) oxidant. The oligonucleotide was purified by reverse phase high-performance liquid chromatography (HPLC) to a final purity of >92% full-length oligomer (determined by capillary electrophoresis and strong anion exchange HPLC). There were no detectable levels of endotoxin in the final lyophilized product as determined by limulus amebocyte lysate analysis (Associates of Cape Cod, East Falmouth, MA). Lyophilized bulk drug substance was formulated as a stock concentration of 50 mg/mL in phosphate-buffered saline (pH 7.4) at the concentrations used in this study.
Measurement of serum complement activation in vitro
The effects of antisense oligonucleotides on in vitro complement activation were measured in monkey (rhesus or cynomolgus) and human serum. Dilutions of oligonucleotides were added to normal human or monkey (rhesus or cynomolgus) serum at a 1:10 to 1:20 ratio, v/v. Samples were incubated at 37°C and aliquots removed at the indicated time points. Activation was terminated by placing the aliquots in a container of crushed ice and then prepared for the determination of the complement split products (C3a, C4a, C5a, and Bb). C4a and C5a were measured using a radioimmunoassay kit from Amersham Life Sciences (Little Chalfont, Buckinghamshire, England). C3a and Bb were measured using enzyme-linked immunosorbent assay (ELISA) kits from (Quidel, San Diego, CA). In some experiments, zymosan A (500 μg/mL; Sigma, St. Louis, MO) or cobra venom factor (CVF) (2 U/mL; Quidel) were used to activate the alternative pathway in the presence of the oligonucleotide. Each was added at a final volume of 1:20. Factor H level was measured by radial immunodiffusion, largely as previously described [15].
Alternative pathway reconstitution
Segments of the alternative pathway were reconstituted with purified human proteins essentially as described by Keil et al. [16]. C3 convertase activity was assayed by combining C3 (125 μg/mL), factor B (20 μg/mL), and factor D (0.2 μg/mL) in Hank's balanced salt solution buffered with 5 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), pH 7.2. In some experiments, factors H and I were also included at 25 μg/mL and 2 μg/mL, respectively. Incubations were carried out under ambient conditions in the presence of a range of antisense oligonucleotide concentrations. Aliquots were removed at selected intervals and immediately diluted 50-fold in ice-cold ELISA dilution buffer. Complement split products were measured by ELISA (Quidel) according to the manufacturer's instructions.
Protein binding measurement
The analytical method utilized to measure protein–oligonucleotide interactions was surface plasmon resonance performed on a BIAcoreX instrument (Biacore, Inc., Piscataway, NJ). Immobilization of ISIS 2302 was conducted at 5 μL/minute and 25°C. HEPES buffered saline (10mM HEPES, 150 mM NaCl, 3 mM EDTA, 0.005% polysorbate 20) was utilized as the immobilization running buffer. The temperature was maintained at 25°C for the duration of the experiment. Research grade CM5 sensor chips were activated with N-hydroxysuccinimide and N-ethyl-N′-(3-dimethylaminopropyl) carbodiimide (Biacore, Inc.), then immobilized with 200 μg/mL streptavidin (Pierce, Rockford, IL) in 10mM acetate pH 4.5 (Sigma). Ethanolamine hydrochloride (Biacore, Inc.) was added to block unreacted N-hydroxysuccinimide esters. ISIS 2302 was biotinylated using 5′-biotin phosphoramidite (Glen Research, Baltimore, MD). Biotinylated ISIS 2302 was captured onto the streptavidin-coated chip over a single flow cell. The free streptavidin sites of the ISIS 2302 flow cell and the control flow cell were capped with 2.5 mM D-biotin in HEPES buffered saline.
Kinetic binding experiments of ISIS 2302–factor H and ISIS 2302–protein C3 (Advanced Research Technologies, San Diego, CA) interactions were conducted at a flow rate of 40 μL/min, utilizing Dulbecco's phosphate-buffered saline (Life Technologies, Inc., Baltimore, MD) as the running buffer. Factor H and protein C3 were dissolved in Dulbecco's phosphate-buffered saline at concentrations of 10 nM to 5 μM and 25 nM to 1 μM, respectively. Binding surfaces were regenerated with pulses of 4–8 M Urea. Sensorgrams were evaluated using BiaEval Software version 3.0 after subtracting the values from the streptavidin-biotin only surface from the ISIS 2302 surface.
Measurement of complement activation in dogs
Dogs (4 males) were treated with a single dose of 10 or 20 mg/kg ISIS 2105 administered as a 2-minute i.v. infusion. Blood samples were collected predose, and at 2, 10, 30, 60, 120, 240, and 360 minutes after end of infusion. The status of the complement system was evaluated by determination of total hemolytic complement (CH50), and comparison of changes relative to baseline values. Plasma was collected at the same time points for measurement of oligonucleotide concentration by capillary gel electrophoresis as described by Leeds et al. [17]. All animal husbandry procedures were in full compliance with AAALAC guidelines.
Results
Oligonucleotide-induced complement activation in monkey and human serum
It is critically important for the therapeutic use of phosphorothioate oligonucleotides to fully understand their potential interactions with the complement cascade in human serum. Toward this end, an in vitro model has been developed to study the mechanisms and species sensitivity of complement activation by phosphorothioate oligonucleotides. Incubation of normal monkey serum with various concentrations of ISIS 2302 for 30 minutes at 37°C selectively activated the alternative pathway of complement. Rapid increases in Bb, C3a, and C5a were observed in the absence of any change in the C4a concentration after incubated 100 μg/mL ISIS 2302 in monkey serum (Fig. 1). This profile of split product generation is the same as that seen in vivo for treated monkeys [8]. The concentration-response and kinetics of complement by ISIS 2302 activation in vitro also agreed with in vivo monkey results. In the in vitro model, 50 μg/mL of ISIS 2302 produced a low and somewhat variable level of complement activation, but higher concentrations of 100 to 500 μg/mL markedly increased alternative pathway activation (Fig. 2). By comparison, the threshold plasma concentration for total oligonucleotides causing activation in vivo was approximately 70 to 80 μg/mL for the first generation phosphorothioate oligonucleotides with short-term i.v. infusion [8]. Rapid complement activation is also characteristic of the in vivo response. The kinetics of activation generally reached a plateau by 15 to 20 minutes with 1-hour i.v. infusion (data not shown).
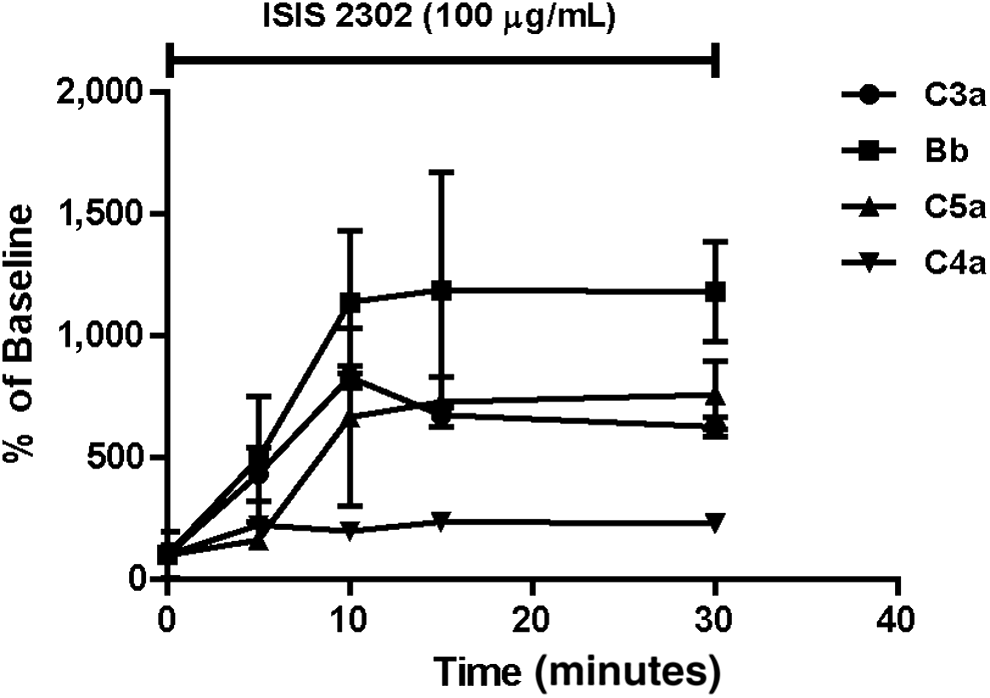
Activation of alternative complement pathway by ISIS 2302 in isolated monkey serum. Monkey serum from 3 individual animals was incubated in the presence of 100 μg/mL ISIS 2302 at 37°C. Samples were obtained at the indicated times, the reaction terminated, and complement split products measured as indicated in Materials and Methods. Expressed values are the mean and standard deviation of the relative increase in split product compared with baseline concentrations.
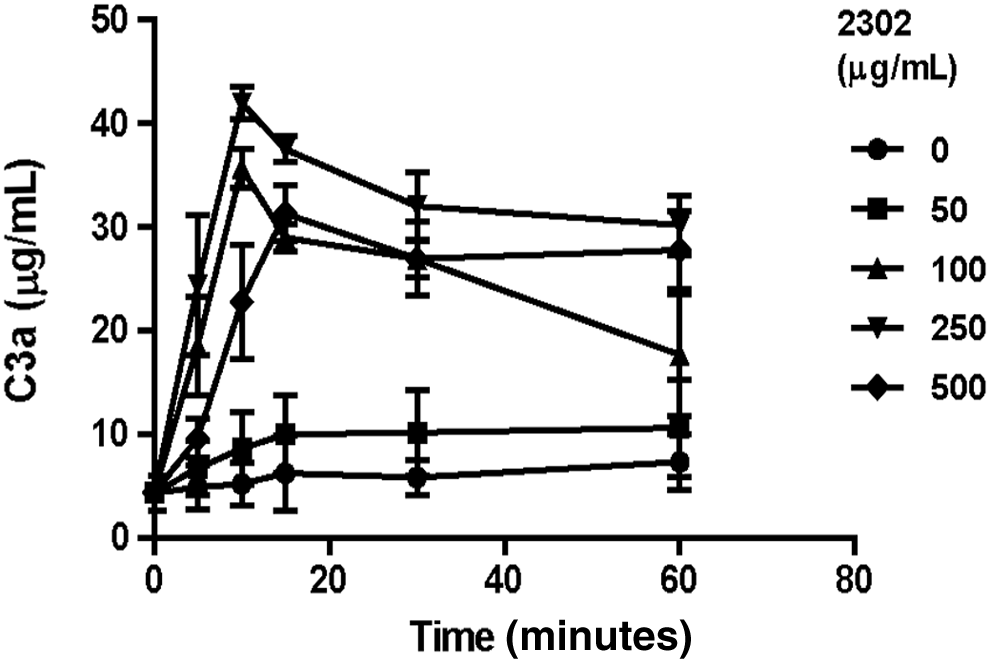
ISIS 2302 produces a concentration- and time-dependent increase in C3a production in monkey serum. Monkey serum from 3 individual animals was incubated in the presence of increasing concentrations of ISIS 2302 at 37°C. Samples were obtained at the indicated times, the reaction terminated, and complement split products measured as indicated in Materials and Methods. Expressed values are the mean and standard deviation of C3a concentrations.
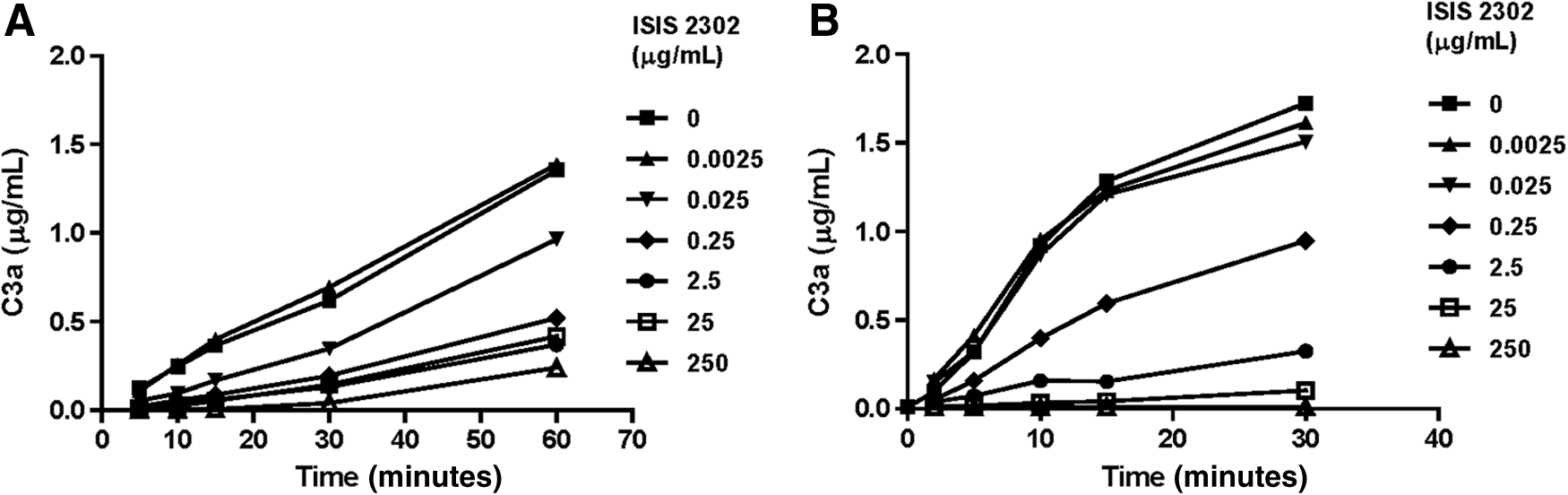
Interestingly, the pattern of complement activation in human serum was distinct from that in monkey (i.e., less activation over a more narrow concentration range in human serum). Although there were minimal increases in C3a at low concentrations (25 and 50 μg/mL) of oligonucleotide in human serum, there was no increase in split product formation at concentrations ≥100 μg/mL, and often there was even a decrease relative to baseline values (Fig. 3A). A similar difference in sensitivity for oligonucleotide-mediated activation was observed from a representative second generation 2′-methoxyethyl (MOE)–modified phosphorothioate oligonucleotide, ISIS 420476. In this case, similar concentration-dependent increase in C3a was observed in serum from cynomolgus monkeys, but there was no increase in the split product in human serum over the same concentration range (Fig. 3B). This is representative of many sequences studied to date.
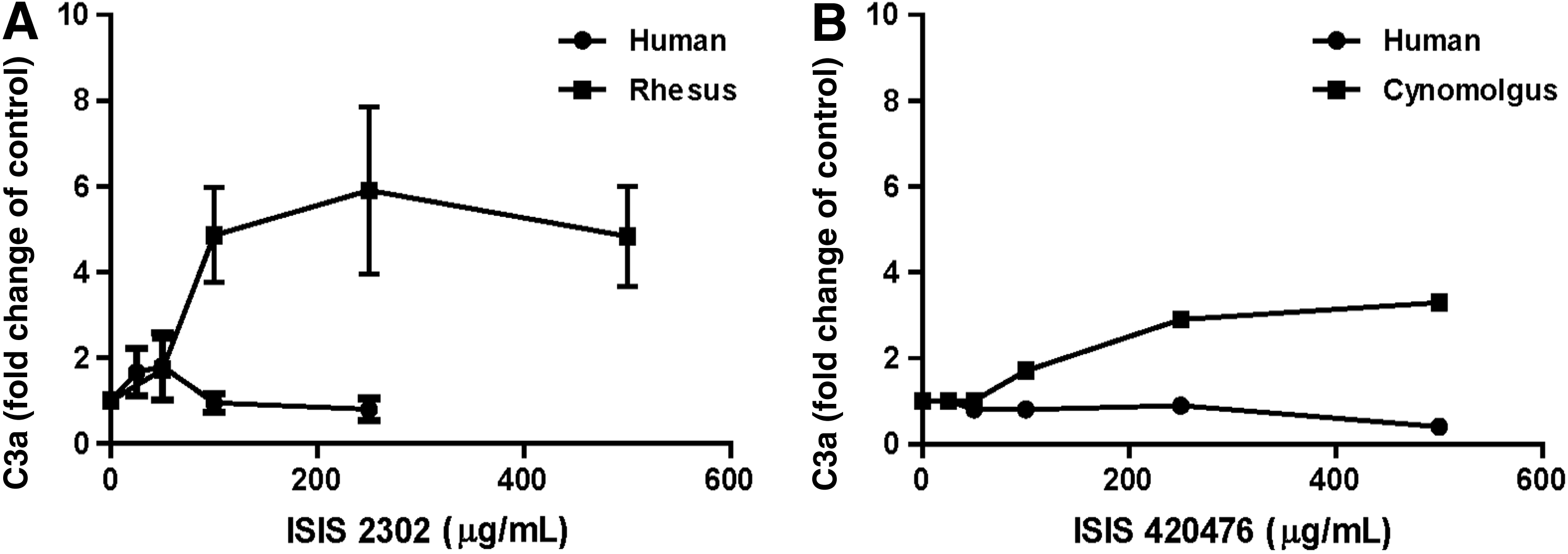
Comparison of C3a production in monkey serum or human serum following treatment with ISIS 2302
Inhibition of zymosan and CVF-induced complement activation by ISIS 2302
Other polyanions have been shown to either activate or inhibit the alternative complement pathway. These effects appear to depend on the concentration of the polyanion as well as the presence of an activation surface [16,18]. Evidence of a biphasic concentration-response was observed for ISIS 2302 (Fig. 3A). The ability of ISIS 2302 to inhibit activator-induced complement activation was investigated in both monkey and human serum. Zymosan was chosen as the surface-activating agent for the alternative pathway. CVF was chosen to selectively investigate ISIS 2302 effects on the C3 convertase. The concentrations of zymosan or CVF used to stimulate complement activation had been titrated to a level that produced approximately 50% of full activation. Complement activation was enhanced in monkey serum in the presence or absence of zymosan or CVF at concentrations of ISIS 2302 up to 500 μg/mL. At ISIS 2302 concentration >500 μg/mL, complement activation induced by either zymosan or CVF was inhibited (Fig. 4). By comparison, in human serum, ISIS 2302 did not enhance activation, and complement activation by zymosan or CVF was suppressed by concentrations of oligonucleotide as low as 25 to 50 μg/mL (Fig. 5). Thus, a 10- to 20-fold higher concentration of ISIS 2302 was required to inhibit complement activation in monkey serum compared to human serum.

Inhibition of complement pathway by ISIS 2302 in monkey serum following activation by either zymosan or cobra venom factor (CVF). Zymosan or CVF were used to activate either the alternative pathway or C3, respectively, and concentrations were titrated to produce approximately 50% of maximum complement activation. Monkey serum from 3 individual animals was added to increasing concentrations of ISIS 2302 in the presence or absence of each activating agent and incubated at 37°C for 30 minutes. The reaction was terminated and complement split products measured as indicated in Materials and Methods. Expressed values are the mean and standard deviation of C3a concentrations.
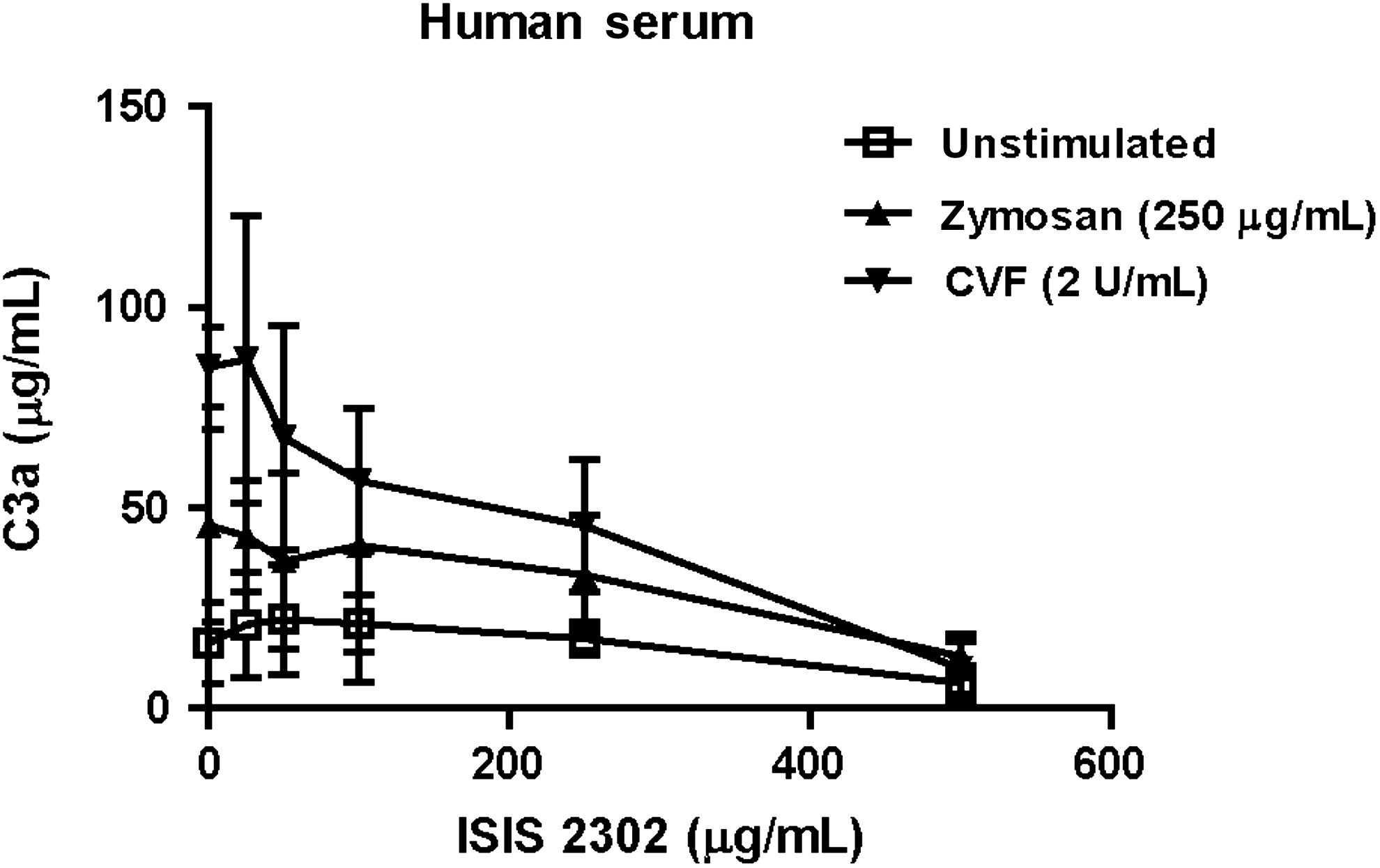
Inhibition of complement pathway by ISIS 2302 in human serum following activation by either zymosan or CVF. Zymosan or CVF were used to activate either the alternative pathway or C3, respectively, and concentrations were titrated to produce approximately 50% of maximum complement activation. Human serum from 3 individual donors was added to increasing concentrations of ISIS 2302 in the presence or absence of each activating agent, and incubated at 37°C for 30 minutes. The reaction was terminated and complement split products measured as indicated in Materials and Methods. Expressed values are the mean and standard deviation of C3a concentrations.
Disruption of alternative pathway regulation by ISIS 2302
One potential mechanism for complement activation by ISIS 2302 may be its ability to interfere with factor H function. In monkey serum, ISIS 2302 induced an apparent concentration-dependent decrease in factor H detected by immunoassay (Fig. 6). An apparent decrease in circulating factor H concentrations was also reported following intravenous injection of ISIS 2302 in monkeys [8]. The apparent decrease in factor H caused by ISIS 2302 might be due to oligonucleotide protein interaction, which prevents the identification of factor H by the immunoassay antibody. Addition of human factor H to monkey serum reduced the effect of ISIS 2302 on alternative pathway, further implicating an interaction between ISIS 2302 and factor H. Human factor H at concentrations as low as 3 μg/mL showed inhibition of complement activation by ISIS 2302 in monkey serum (Fig. 7). Higher concentrations (e.g., at 30 μg/mL) of human factor H prevented all spontaneous and activator-induced activation of complement in monkey serum. In this regard, human factor H appears to be a potent inhibitor of the alternative pathway in monkey complement.
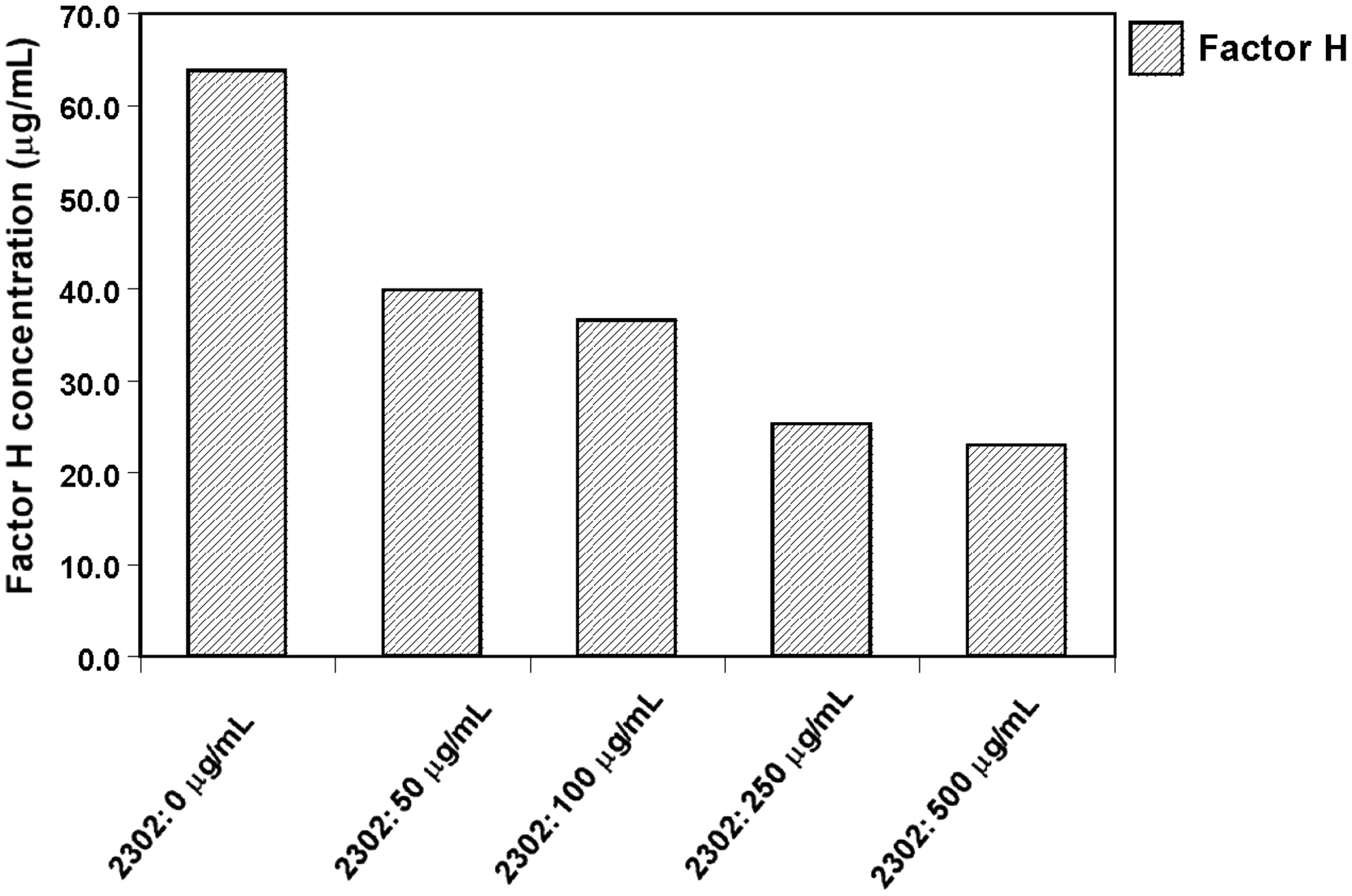
Effect of increasing ISIS 2302 concentration on apparent concentration of Factor H in monkey serum. Serum from 3 individual monkeys was incubated in the presence of increasing concentrations of ISIS 2302 at 37°C for 30 minutes. The reaction was terminated, and factor H levels were measured by radial immunodiffusion as indicated in Materials and Methods. Expressed values are the mean of factor H concentrations.

Addition of purified human factor H prevented alternative pathway complement activation by ISIS 2302 in monkey serum. Factor H concentrations are low relative to physiologic concentration in human (500 μg/mL). Monkey serum from 3 individual animals was added to increasing concentrations of ISIS 2302 in the presence of the indicated concentration of human factor H and incubated at 37°C for 30 minutes. The reaction was terminated and complement split products measured as indicated in Materials and Methods. Expressed values are the mean and standard deviation of C3a concentrations.
The inhibitory effects of ISIS 2302 on the human complement cascade were confirmed by reconstituting the complement cascade with purified human proteins. In these experiments, there was no evidence of activation of either the reconstituted C3 convertase (factors D, C3, and B) or the alternative pathway (factors D, C3, B, H, and I) over a broad range of oligonucleotide concentrations (0.0025 to 250 μg/mL). There was, however, a concentration-dependent inhibition of both reconstituted C3 convertase and alternative pathway activity by ISIS 2302 (Fig. 8A, B). The concentration-response in both reconstituted systems was similar, with approximately 0.25 μg/mL ISIS 2302 producing 50% inhibition of C3a production. These observations using ISIS 2302 and purified human proteins were consistent with the effects observed in human serum, which favored inhibition over activation of the alternative pathway of complement. Similar examination of these effects on reconstituted monkey C3 convertase or alternative pathways were not performed because the purified factors are not readily available.
Effect of ISIS 2302 on the activity of C3 convertase
Binding of ISIS 2302 to complement protein factors
Protein binding studies of ISIS 2302 and complement factors were performed using surface plasmon resonance. Equilibrium binding constants (KD) were calculated using association (ka) and dissociation (kd) rate constants determined from the curves generated by fitting data to kinetic models. Both C3 and factor H appeared to have a high affinity for ISIS 2302 with binding constants in the low (KD1) to mid nM (KD2) range (Table 1). Similar high affinity binding has been characterized for proteins of the coagulation cascade (thrombin) and α2-microglobulin. However, the protein binding affinity of other abundant plasma proteins such as albumin are an order of magnitude less (KD=mid-μM range; data not shown) [19]. Demonstration of high affinity binding between ISIS 2302 and key components of the alternative pathway such as factor H is consistent with the proposed mechanism of activation. Inhibition of complement activation and C3 convertase activity can be explained by binding of the oligonucleotide to C3.
KD1, low-range equilibrium binding constant; KD2, mid-range equilibrium binding constant.
Comparison of complement activation in different animal species
Differences in the species sensitivity to complement activation by phosphorothioate oligonucleotides was clearly evident in the various animals models tested. In dogs, intravenous infusion of 10 and 20 mg/kg ISIS 2105 over 10 minutes did not decrease CH50 (Fig. 9). Due to lack of available species specific reagents, the complement split products in dogs were not measured. The resulting peak plasma concentration of oligonucleotide in dogs (295 μg/mL) exceeded the threshold concentration previously shown to cause complement activation in monkeys (i.e., 70 to 80 μg/mL total oligonucleotide with short-term i.v. infusion). Based on these results, complement activation by phosphorothioate oligonucleotides appears not to occur in dogs. There is also evidence that oligonucleotide-induced complement activation does not occur in mice, rats, or rabbits (data not shown). This conclusion is supported by the measurement of CH50 in rats and the absence of acute anaphylactoid-like reactions in these animal species when treated with high doses of phosphorothioate oligonucleotides. Thus, cynomolgus and rhesus monkeys appear to be unique in their relative sensitivity to complement activation by phosphorothioate oligonucleotides.

Intravenous injection of ISIS 2105 in dogs does not activate complement pathway. The activation state of complement pathway was assessed by measuring total hemolytic complement activity (CH50) at the indicated time points (see Materials and Methods). Plasma oligonucleotide peak concentration (Cmax) were determined by measuring concentration at the 2-minute time point by capillary gel electrophoresis. Data are the mean of 4 dogs at each dose level.
Effects of chemically modified oligonucleotides on complement activation
The nature of the interaction of the complement cascade and oligonucleotides was further explored by synthesizing oligonucleotides of the same sequence, but with various chemical modifications known to influence pharmacology [20–22]. Incorporation of 2′-MOE derivative on the ribose sugars of the oligonucleotide increases hybridization affinity to target mRNA but further stabilizes the compound against nuclease degradation of both phosphorothioate and phosphodiester linkages. Ribose sugar units with 2′-MOE were also incorporated into an oligonucleotide that contained a mixed backbone of phosphorothioate and phosphodiester linkages. The presence of phosphodiester linkages further reduces serum protein binding, while maintaining the negative charge state of the molecule [20]. Complement activation in monkey serum appeared to be reduced by 2′-MOE modification of either full phosphorothioate or mixed phosphodiester and phosphorothioate oligonucleotides relative to unmodified oligonucleotides (ISIS 5132 and ISIS 2302). The full phosphorothioate 2′MOE oligonucleotide (ISIS 13650 and ISIS 15839) produced slightly less alternative pathway activation at all concentrations tested (Fig. 10). The mixed backbone 2′-MOE oligonucleotide (ISIS 12854 and ISIS 14725) produced significantly less complement activation, consistent with a decrease in protein-binding affinity compared to ISIS 5132 or ISIS 2302. Similar structure activity relationships have been observed with oligonucleotides having different sequences as well (data not shown).
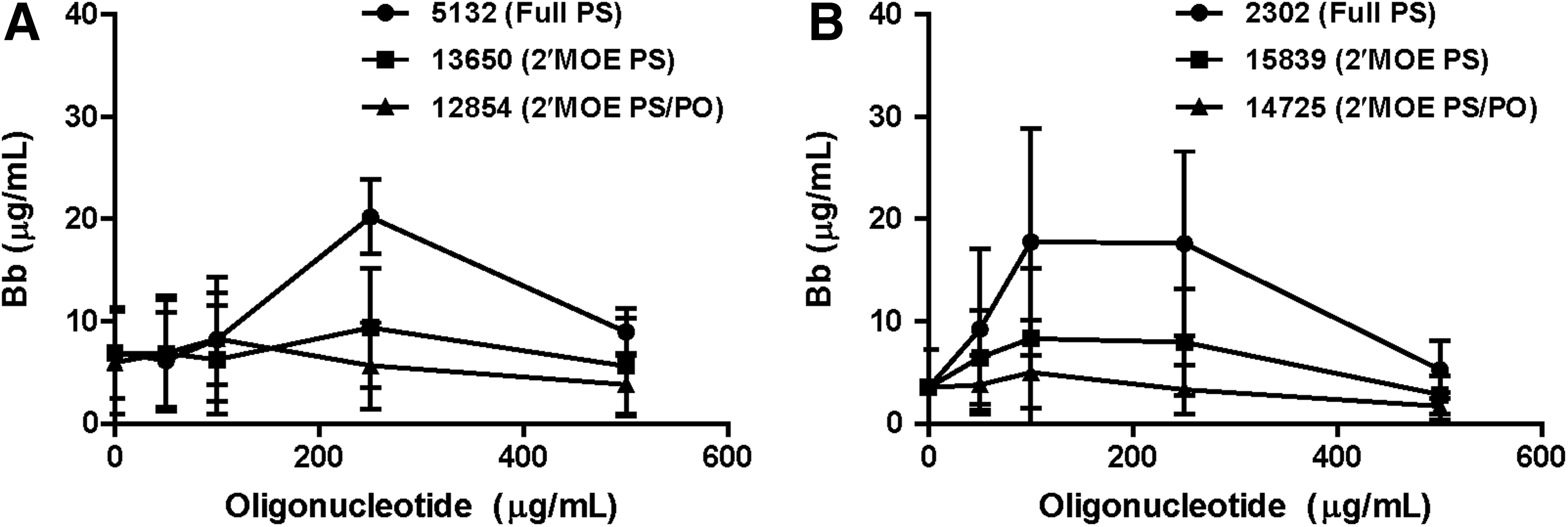
Chemical modification of phosphorothioate oligonucleotide can modulate the ability to activate complement in monkey serum. Two series of oligonucleotides were examined to study the effects of chemical modification, each series of three oligonucleotides share the same sequence. Full phosphorothioate oligonucleotides (ISIS 2302 and ISIS 5132) were compared with oligonucleotides that contained phosphorothioate (PS) linkage and MOE substituents on the 2′-position of ribose (ISIS 15839 and ISIS 13650) or oligonucleotides that have mixed PS and phosphodiester (PO) linkages with 2′-MOE substituents (ISIS 14725 and ISIS 12854). Monkey serum from 3 individual animals was incubated in the presence of increasing concentrations of oligonucleotide at 37°C. Samples were obtained at the indicated times, the reaction terminated, and complement split products measured as indicated in Materials and Methods. Expressed values are the mean and standard deviation of Bb concentrations.
Discussion
Complement activation in monkeys occurs as a consequence of administration of high doses of antisense oligonucleotides and is an undesirable side effect. To better characterize why complement activation is induced by oligonucleotides, studies have been performed to examine both the effects and concentration-response relationships. Activation of the alternative complement pathway in monkeys occurs as a consequence of intravenous infusion of phosphorothioate oligonucleotides and is largely independent of sequence, thus it is considered a class effect of these compounds [5,8,10]. A precise relationship between dose, rate and duration of infusion, plasma oligonucleotide concentration, and complement activation has been described in monkeys. Complement activation by ISIS 2302 in cynomolgus monkeys required a minimum threshold concentration in plasma of 70 to 80 μg/mL of total oligonucleotide with short-term i.v. infusion [8]. Examining the effects of three additional oligonucleotides of similar length but different sequences revealed that their concentration thresholds for activation were similar to ISIS 2302, indicating that the interactions with complement components are predominantly independent of sequence, however the magnitude of response could be varied by the sequences [5]. These observations also clearly indicate that complement activation is hybridization-independent, and suggest that oligonucleotide–protein interactions are most likely central to the underlying mechanism of action.
Complement activation in clinical trials has been avoided by administering dose regimens that produce peak plasma oligonucleotide concentrations below the established threshold for activation. Minimizing peak plasma concentrations of oligonucleotide by prolonging infusion time reduces or prevents complement activation in monkeys [10,23]. Doses as high as 6 mg/kg delivered by a 2-hour i.v. infusion in patients did not produce significant or consistent complement activation [24]. A similar absence of complement activation in clinical trials has been documented for a number of the 2′-MOE modified phosphorothioate oligonucleotides at doses up to 6 mg/kg administered by subcutaneous injection or 1-hour i.v. infusion. Although complement activation has not been observed in clinical applications of antisense therapeutics, the relative sensitivity of humans is difficult to assess because plasma exposure is relatively low compared with that attained in monkey studies.
Although phosphorothioate oligonucleotides have been studied in a number of species including mice, rats, guinea pigs, rabbits, and dogs, complement activation has only been confirmed in rhesus and cynomolgus monkeys. Aside from monkeys, none of these other species have exhibited the acute hemodynamic or hematologic alterations associated with complement activation. Data in this report demonstrate the absence of complement activation in dogs using oligonucleotide dose regimens sufficient for complement activation in monkeys. Possible explanations include differences in protein-oligonucleotide binding affinities that exists in different species or possible differences in stringency of the alternative complement pathway regulation. The absence of available complement cofactors from species other than human has restricted the investigation of the underlying mechanisms of complement activation in monkeys. However, use of the human complement proteins has provided some insights into these mechanisms and the relative sensitivities of human and monkey complement to be activated by oligonucleotides.
The in vitro model for complement activation described in this report will help further understand the basis of species differences and mechanisms of complement activation by phosphorothioate oligonucleotides. ISIS 2302 was used in these studies as a representative of the chemical class for 15- to 25-nucleotide phosphorothioate oligonucleotides. The experimental tools used in this model system, specifically an ELISA assay for human complement split products, were the same as those used in the monkey studies to simplify interpretation of results between models. Measurement of the split products assures an unambiguous interpretation of the status of the complement system. Total hemolytic activity (CH50) was not used to measure complement activation because it does not distinguish between inhibition of enzymatic processes from consumption of complement components. The characteristics of complement activation induced by ISIS 2302 in monkey serum in vitro were consistent with those observed in treated monkeys. Specifically, the profile of split products indicated a selective activation of the alternative pathway (increased Bb and no change in C4a) and confirmed activation of the full cascade (i.e., increases in both C3a and C5a). Furthermore, serum concentrations of ISIS 2302 that activated complement in vitro were similar to blood levels leading to activation in treated monkeys. Complement split products increased within minutes of exposure to ISIS 2302 in both models. Thus, it appears that in monkey complement activation in the in vitro model can confidently be interpreted as representative of that observed in vivo.
One interesting aspect of complement activation by ISIS 2302 in monkey serum was the biphasic profile of the concentration-response curve. Activation of the alternative pathway was concentration dependent up to approximately 500 μg/mL. Higher concentrations appear to inhibit the pathway. In fact, at a concentration of 1,000 μg/mL ISIS 2302 even inhibited complement activation by CVF or zymosan in monkey serum.
Human serum was relatively insensitive to complement activation by either ISIS 2302 or a 2′-MOE oligonucleotides. A biphasic concentration-response curve was also observed with increasing concentrations of ISIS 2302 in human serum. However, human serum was more sensitive than monkey serum to inhibition of complement activation by ISIS 2302. Concentrations below 50 μg/mL resulted in a low level of alternative pathway activation, but no activation was observed at higher concentrations. Consistent with reduced levels of complement activation compared with monkeys, ISIS 2302 inhibited CVF and zymosan-activated complement in human serum at concentrations as low as 50 μg/mL. High concentrations of oligonucleotide also inhibited classical pathway activation by aggregated immunoglobin G in human serum (data not shown). Thus, inhibition can occur for either the classical or alternative pathways.
Other polyanions such as heparin and dextran sulfate have been reported to alter regulation of the alternative complement pathway [25–27]. These interactions are complex, and both activation and inhibition of the alternative pathway have been documented [16]. However, the greater tendency is for polyanions to inhibit complement activation in human serum [26]. Recognition of polyanions on the surface of cells (such as membrane sialic acid) is one manner by which the alternative pathway distinguishes mammalian cells from foreign pathogens, such as bacteria [28]. These surface polyanions promote the interaction of factor H with C3b bound to the surface. In the same manner, soluble polyanions such as dextran sulfate and heparin have also been shown to promote the interaction of factor H with surface-bound C3b [29,30]. These interactions lead to inhibition of the alternative pathway via accelerated conversion of C3b to C3bi by factor I [31]. However, reports on the ability of heparin to enhance this molecular interaction between factor H and fluid-phase C3b are mixed [16,30]. Occupation of a specific site on factor H by heparin is thought to increase the affinity for C3b. Factor H is a 150 kDa cationic protein present in human serum at approximately 500 μg/mL containing a specific binding site for heparin [32]. In addition to having polycationic regions that could bind ISIS 2302 through ionic interactions, factor H has also been identified as a plasma DNA-binding protein [33].
Polyanions, such as heparin, can also inhibit alternative pathway directly by disrupting the interaction between factor B and C3, thus preventing formation of the C3 convertase [16,29,34]. The ability of high concentrations of ISIS 2302 to inhibit CVF-induced complement activation is consistent with this type of interaction. Based on our reconstitution experiments, ISIS 2302 was a potent inhibitor of C3 convertase, and this appears to be the predominant mechanism for inhibition of zymosan and CVF-induced complement activation by ISIS 2302 in human serum. ISIS 2302 may interfere with C3 convertase by binding to C3 and preventing Bb binding. Interaction of ISIS 2302 with human C3 could also explain the inhibition of CVF-induced activation, since this process depends on a CVF–C3 interaction. The relatively lower inhibition of CVF-induced complement activation by ISIS 2302 in monkey serum may reflect a lower affinity of oligonucleotide for monkey C3. Currently, purified monkey C3 is not available to test this hypothesis.
Although seemingly paradoxical, the interaction between factor H and a polyanionic substance can also lead to complement activation. Factor H is an important regulatory component of the alternative pathway that works in conjunction with factor I to inactivate the alternative pathway C3 convertase, which is constitutively active at a low level in serum (i.e., the so-called tick-over mechanism) [31]. If factor H function is impaired, then C3 convertase becomes unregulated and fluid-phase activation of alternative pathway proceeds, consistent with the observed response to ISIS 2302 treatment [27]. Complement activation by heparin and other polyanions is thought to result from sequestration of factor H or prevention of its interaction with C3 convertase, leading to unchecked amplification of the alternative pathway [16,18,25]. The polyanionic nature of phosphorothioate oligonucleotides is different from sulfated polysaccharides such as heparin or dextran. However, protein-binding studies indicate that factor H does interact with ISIS 2302, and factor H immunoassay results (in vitro and in vivo) suggest that this interaction either sequesters factor H or blocks the interaction with the factor H antibody [8]. This disruption of the C3b–factor H interaction is believed to lead to an activation of the alternative complement pathway in monkeys.
A functional role for factor H in alternative pathway activation in monkey serum was suggested by the ability of human factor H to prevent complement activation by ISIS 2302. Addition of human factor H at concentrations as low as 3 μg/mL started to show inhibition of ISIS 2302-induced complement activation in monkey serum. Higher concentrations of factor H (e.g., 30 μg/mL) completely inhibited any complement activation in monkey serum, whether spontaneous or ISIS 2302 induced. By comparison, human factor H had only modest effects in human serum on either zymosan or CVF-induced complement activation at concentrations as high as 250 μg/mL. Although not thoroughly investigated, there may be a marked difference in potency of factor H between these species. Human factor H added to monkey serum appeared to provide very potent inhibition of alternative pathway at concentrations approximately 100-fold below physiologic concentrations (e.g., 500 μg/mL). There could be several explanations for this species difference, including potency differences between monkey and human factor H. Another possible explanation for the greater sensitivity of monkey complement to ISIS 2302 may be differences in factor H concentrations between the species. Alternatively, intrinsic differences in factor H between species may make it more or less susceptible to the inhibitory effects of phosphorothioate oligonucleotides. These observations provide insight into both the nature and mechanism of complement activation by oligonucleotides in monkeys, as well as confirm significant differences in species sensitivity to these molecules.
Footnotes
Author Disclosure Statement
Authors affiliated with Isis Pharmaceuticals, Inc. (SPH, SM, RSG, AAL) are or have been employees and shareholders of the company that is submitting the paper.