
Abstract
The highly conserved and ubiquitous molecular chaperone heat shock protein 70 (Hsp70) plays a critical role in protein homeostasis (proteostasis). Controlled by its ATPase activity, Hsp70 cycles between two conformations, Hsp70-ATP and Hsp70-ADP, to bind and release its substrate. Chemical tools with distinct modes of action, especially those capable of modulating the ATPase activity of Hsp70, are being actively sought after in the mechanistic dissection of this system. Here, we report a conformation-specific RNA aptamer that binds only to Hsp70-ATP but not to Hsp70-ADP. We have refined this aptamer and demonstrated its inhibitory effect on Hsp70's ATPase activity. We have also shown that this inhibitory effect on Hsp70 is independent of its interaction with the Hsp40 co-chaperone. As Hsp70 is increasingly being recognized as a drug target in a number of age related diseases such as neurodegenerative, protein misfolding diseases and cancer, this aptamer is potentially useful in therapeutic applications. Moreover, this work also demonstrates the feasibility of using aptamers to target ATPase activity as a general therapeutic strategy.
Introduction
T
There are many pathologically stressful conditions, of which an interesting case is found in cancer cells adapting to adverse microenvironments through specific mutations in proteins that deregulate cell growth. To maintain an increased proliferation rate compared to nontransformed cells, they exhibit an increased requirement for nutrients and accelerated metabolic processes in a hypoxic environment, resulting in growth under highly stressful conditions. To tolerate the higher levels of free radicals and the increased protein damage, transformed cells further adapt by increasing the requirements for molecular chaperones. Elevated levels of heat shock proteins, especially Hsp70 and Hsp90, are observed in nearly every type of cancer cells [3]. More specifically, these chaperones have been shown to stabilize the folding of mutated proteins critical for cancer maintenance, such as the dominant negative forms of the p53 tumor suppressor, various hormone regulated receptors, and non-receptor tyrosine kinases [4]. In light of these findings, specific inhibitory reagents to different heat shock proteins will not only make useful tools in mechanistic studies, but also have potential therapeutic applications.
Hsp70 is composed of a 45-kDa N-terminal nucleotide binding domain (NBD) and a 30-kDa substrate binding domain (SBD) with a C-terminal “lid.” The NBD binds and hydrolyzes adenosine triphosphate (ATP), while the SBD binds a hydrophobic peptide segment of the substrate protein. The conformation of Hsp70 changes between its different nucleotide bound forms [5–7]. In the ATP bound form, the NBD and SBD are docked together with the “lid” domain in open position, resulting in a high off rate for substrates [5,7,8]. In the adenosine diphosphate (ADP) bound form, NBD and SBD become uncoupled, moving independent of each other [6,9], and the “lid” domain closes, decreasing the off rate for substrates. Driven by its ATPase activity, Hsp70 cycles between these two distinct conformations, which regulates its substrate binding and release. By modulating its ATPase activity, different aspects of Hsp70 function can be regulated. For example, the Hsp70 co-chaperone Hsp40 stimulates the intrinsic ATPase activity of Hsp70 and accelerates its substrate binding and release cycle [10,11]. For this reason, specific reagents that inhibit ATPase activity are useful in elucidating the mechanism of Hsp70 function. Moreover, targeting ATPase activity has also been proposed as an attractive and widely applicable therapeutic strategy [22].
Compared with another key heat shock protein, Hsp90, fewer small molecule ligands have been developed for Hsp70 [12]. While they have been used to study Hsp70 mediated pathway regulation [13–16,18], most of them bound and stabilized Hsp70's high substrate affinity states: Hsp70-ADP or the structurally similar apo-state [14,17,19,20]. In addition, small molecules identified through screening often exhibit low specificity and affinity [12]. Therefore, we believe there is a need to develop alternative chemical tools to dissect biological processes involving this important protein. Peptide aptamers inhibiting Hsp70 function had been isolated, but their mechanism of action and their effect on Hsp70 ATPase activity are not known [21]. In this contribution, we describe our effort to develop an RNA aptamer that inhibits Hsp70 ATPase activity. An essential requirement for such an aptamer is that it should bind to the Hsp70-ATP conformation. With that requirement in mind, we intentionally designed a special in vitro selection scheme through which we isolated an RNA aptamer referred to as AptHsp70-1. The aptamer was found to be specific for Hsp70-ATP and did not recognize Hsp70-ADP. Interestingly, it inhibited Hsp70's ATPase activity both in the presence or absence of the Hsp40 co-chaperone and hence indicated the existence of a novel regulatory site with potential therapeutic utility.
Materials and Methods
Proteins and plasmids
The expression plasmid for human Hsp70-1 (hHspA1A) with a C-terminal His-tag was purchased from Genecopoeia (cat. no. EX-Z5704-B31). The protein was expressed in Escherichia coli strain BL21-AI and purified by nickel-affinity column (Sigma) according to the manufacturer's instructions. Purified protein fractions were pooled together and concentrated using protein concentrators (Pierce Scientific, 9 kDa molecular weight cutoff) and buffer-exchanged using the same concentrators into protein storage buffer (50 mM HEPES/pH 7.4, 100 mM NaCl, 3 mM DTT, 2 mM MgCl2, 45% glycerol, 100 μM ATP). Human Hsp40 (Hdj1) and E. coli Hsp40 (DnaJ) were purchased from Enzo Life Sciences. Human Hsc70 (hHspA8) was purchased from Sino Biological.
In vitro selection
The initial RNA pool contained ∼1.8×1015 different sequences, each having a 50-nt randomized region in the middle flanked by 25-nt constant regions on either side [23]. Selection was performed as described previously [23] with minor modifications. The binding buffer contained 20 mM HEPES/pH 7.4, 150 mM NaCl, and 10 mM MgCl2. ATP (final concentration 100 μM) was also added in the protein storage buffer to maintain Hsp70 in the ATP bound state during selection. Bound and unbound RNA fractions were separated by nitrocellulose filters. After six cycles of selection and amplification, the final pool in the form of DNA was cloned into pGEM-3Z vector (Promega), and aptamers were identified by sequencing and binding assays.
Aptamers sequences and secondary structure
The sequences of AptHsp70-1, its deletion constructs and derivatives are listed in Tables 1 and 2.
Apt, aptamer; Hsp, heat shock protein.
Secondary structures were predicted using the Vienna RNA server [24]. The secondary structure of AptHsp70-1 was predicted by preserving the minimum free energy structure of the minimized aptamer, AptHsp70-1-61. Secondary structures of all other aptamer constructs are their minimum free energy structures.
Electrophoretic mobility shift assays
Binding reactions were performed in 15 μL 1×binding buffer (same as the binding buffer used for in vitro selection). A typical binding reaction contained ∼30 nM of 32P-labeled RNA aptamer and various amounts of Hsp70. For binding reactions used to measure dissociation constant (Kd), 0.2 nM of 32P-labeled RNA aptamer was used to ensure an excess protein concentration. The RNA aptamers were labeled with [α-32P]ATP (Perkin Elmer) using the MAXIscript in vitro transcription kit (Applied Biosystems). The labeled aptamers were heated to 65°C for 10 minutes and cooled to 37°C for 10 minutes before mixing with the protein. The binding reactions were incubated at 37°C for 30–40 minutes, mixed with gel loading buffer (50% glycerol, 0.01% bromophenol blue, and 0.01% xylene cyanol) and run on 2.5% agarose gel or 4.8% polyacrylamide gel (37.5:1 Acrylamide:Bis) in 0.5×TBE buffer. The gels were run for 3 hours at 4°C, dried, exposed to phosphor screen, and imaged using a Typhoon scanner (GE Healthcare). The protein-bound and unbound RNA bands were quantified using the ImageQuant software (GE Healthcare). Binding curves were constructed using KaleidaGraph 4.1. The data points were fitted using Hill equation [y=xa/(ba+xa), where a represents the Hill coefficient and b represents the apparent dissociation constant Kd].
In-line probing
In-line probing was performed as described previously [25,26], with minor modifications. Radio-labeled RNA aptamer was transcribed using the MAXIscript in vitro transcription kit (Applied Biosystems). As AptHsp70-1 begins with a G nucleotide at the 5′ end, the transcription reaction was supplemented with [γ-32P]GTP (Perkin Elmer) along with all four unlabeled nucleotides. The labeled full-length RNA aptamer was gel purified and incubated in binding buffer in the presence or absence of ATP or ADP at room temperature for 65 hours. The samples were mixed with RNA gel loading buffer 2 (Ambion) heated to 65°C for 1 minute and then run on 12% polyacrylamide gel [19:1 Acrylamide:Bis with 50% (w/v) urea] for 2.5 hours, exposed to phosphor screen, and imaged using a Typhoon scanner (GE Healthcare).
ATPase assay
Single turnover ATPase assay was performed as described previously [27]. Briefly, 15 μL reaction with 200 nM Hsp70, 200 nM Hdj1 or DnaJ, and 30 nM [α-32P] ATP in buffer (20 mM HEPES/pH 7.4, 50 mM KCl, 10 mM MgCl2, and 2 mM DTT) was incubated at 37°C for the indicated time. From the reaction 0.8 μL was spotted onto a thin layer chromatography (TLC) plate and run in 0.5 M LiCl and 0.5 M formic acid. For the kinetic assay, a 0.8 μL sample was spotted on to a TLC plate at various time points. Result was visualized and quantified by phosphor-imager analysis (GE Healthcare). Kcat values were determined from the slope of the best fit line plotted using KaleidaGraph 4.1.
Results
Identification of an aptamer for a predetermined Hsp70 conformation
The stress inducible Hsp70s and their constitutive cognates Hsc70s are highly conserved in evolution [28]. We used the predominant stress inducible human isoform HspA1A (a.k.a. Hsp70-1) as the target to generate RNA aptamers for its ATP-bound form. Since Hsp70 has an intrinsic ATPase activity, simply adding ATP to the protein would not keep the target preparation in a conformationally pure and stable Hsp70-ATP state required during aptamer selection. However, ATP hydrolysis is absolutely dependent on a pair of K+ ions at the bottom of the nucleotide binding cleft [29,30]. Replacing K+ by Na+ would inhibit the ATPase activity of Hsp70 [29,30], and as a result the cyclic conformational change would be stopped at the ATP-bound state. Based on this reasoning, we avoided K+ and used Na+ in the buffers for protein purification, storage, and aptamer selection. One hundred micromolars of ATP was also added to the protein preparation to maintain the ATP bound state. As shown in Fig. 1A, the Hsp70 we prepared was active in ATP hydrolysis when K+ is present and the ATPase activity was inhibited by substituting K+ with Na+. An alternate strategy to “freeze” the Hsp70-ATP conformation would be to use non-hydrolyzable ATP analogs to displace ATP. We adopted K+ substitution because K+ also favors the formation of RNA G-quadruplex structures found in filter binders, which may cause high background in the SELEX process [31,32].

Generation of an RNA aptamer for Hsp70-ATP.
Using this Hsp70-ATP preparation as the target, six cycles of selection and amplification were performed starting from an initial pool of∼1.8×1015 RNA molecules. DNA pool from the sixth cycle was cloned into pGEM-3Z plasmid vector and transformed into E. coli. Fifty clones were randomly selected and sequenced, and individuals were tested for binding to identify aptamers. One unique aptamer sequence (AptHsp70-1) was identified that bound to Hsp70 in electrophoretic mobility shift assay (EMSA) (Fig. 1B). Interestingly, AptHsp70-1 was not able to bind the closely related isoform Hsc70 in EMSA (Fig. 1C). The sequence and predicted secondary structure of the aptamer is shown in Fig. 1D. The remaining 49 clones were represented by three sequences that might be selected nonspecifically by unknown components in the binding or amplification reactions.
Conformational specificity and affinity of AptHsp70-1 binding to Hsp70
EMSA was used to verify whether AptHsp70-1 binds to Hsp70-ATP conformation. As expected, although the aptamer bound Hsp70 avidly in the presence of excess ATP, the binding was not observed in the presence of ADP (Fig. 2A). However, there are two possible explanations for this result. First, AptHsp70-1 was specific to Hsp70-ATP and could not recognize Hsp70-ADP. Second, the aptamer structure was affected by ATP, ADP, or both, so that only in the presence of ATP, the aptamer folded into an active conformation that was able to bind Hsp70. As free ATP was present in the binding mixture during the process of selection, it was necessary to exclude the second possibility by an independent assay. For this purpose we performed in-line probing for the aptamer in the presence of ATP or ADP and compared the patterns of bands with those generated in the absence of any nucleotide. If either nucleotide interacted with the aptamer and changed its folding, different in-line cleavage patterns would be revealed [25].
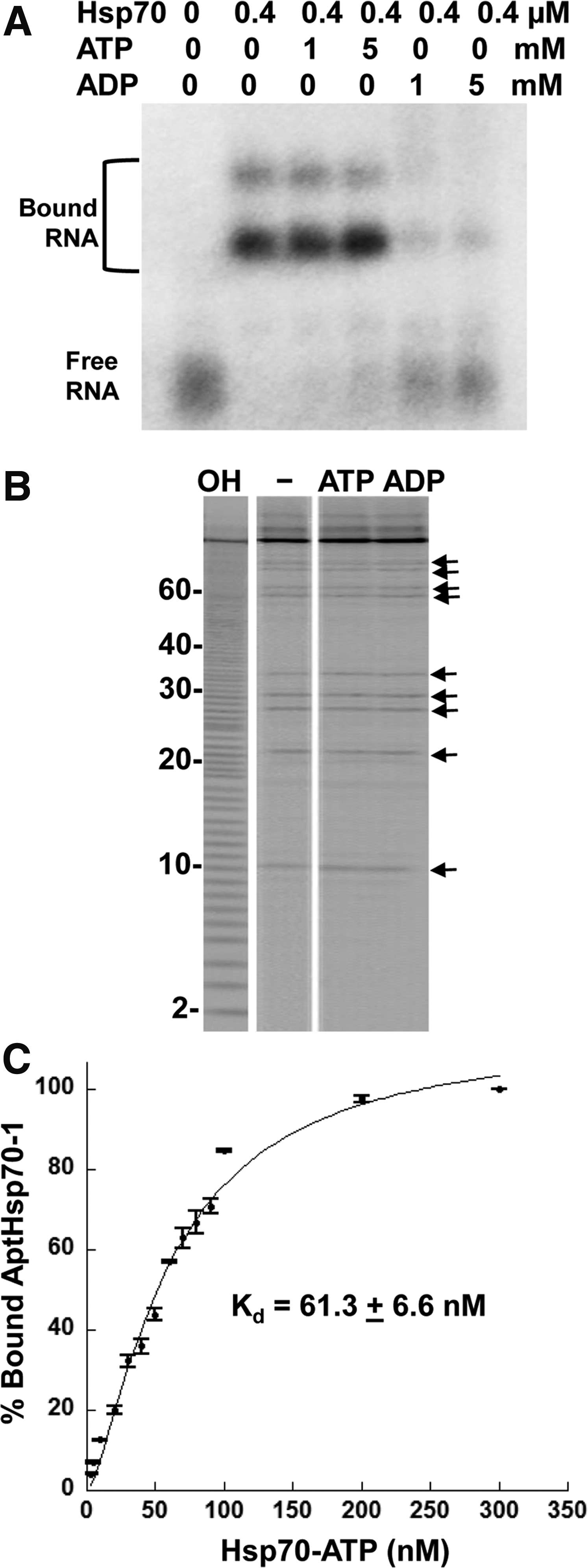
Conformation specificity of AptHsp70-1.
However, as shown in Fig. 2B, an identical pattern was observed in the absence or presence of ATP or ADP, indicating that ATP or ADP did not bind to AptHsp70-1 and affect its structure. Therefore, in the binding assay shown in Fig. 2A, ATP or ADP only affected Hsp70 but not the aptamer, and AptHsp70-1 is indeed a conformation-specific aptamer for Hsp70-ATP. In addition, in-line probing of AptHsp70-1 also supported the predicted secondary structure of the aptamer, with cleavages occurring mostly in the single stranded regions (bands in Fig. 2B are mapped to the positions on the structure shown in Fig. 1D). To measure the affinity of aptamer binding we performed EMSA with different concentrations of Hsp70, ranging from 2.5 nM to 300 nM. As shown in Fig. 2C, the apparent dissociation constant (Kd) for AptHsp70-1 binding to Hsp70 was 61.3±6.6 nM.
Sequence and structural refinement of AptHsp70-1
To find the critical regions of AptHsp70-1 involved in binding, we performed deletion analysis. The aptamer was first sequentially deleted from the 3′ end and the various constructs were tested for Hsp70 binding by EMSA (Fig. 3A, B). The aptamer retained binding activity until more than 35 bases were deleted from the 3′ end (Fig. 3B). Further deletions from either the 3′ (right panel of Fig. 3B) or the 5′ end (Fig. 3C) resulted in negligible binding. Although AptHsp70-1-61, the 61-nt minimized aptamer (Fig. 3D), was able to bind Hsp70, the binding detected in EMSA was considerably weaker than the full-length aptamer (right panel of Fig. 3B). The reduction seemed to be caused by structural instability of the active conformation, as most of the RNA was found in a protein-indifferent band (box 1 in Fig. 4A) that migrated faster on the native gel than the bands that shifted in the presence of protein (box 2 in Fig. 4A). To increase the fraction of the active conformation, we added a “GC-clamp” stem to fortify the secondary structure of AptHsp70-1-61 and a slightly longer version, AptHsp70-1-66 (Fig. 4B). This addition resulted in a marginal improvement in binding for AptHsp70-1-61 but a significant increase for AptHsp70-1-66 (Fig. 4C and Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/nat).

Deletion analysis of AptHsp70-1 to minimize the aptamer.
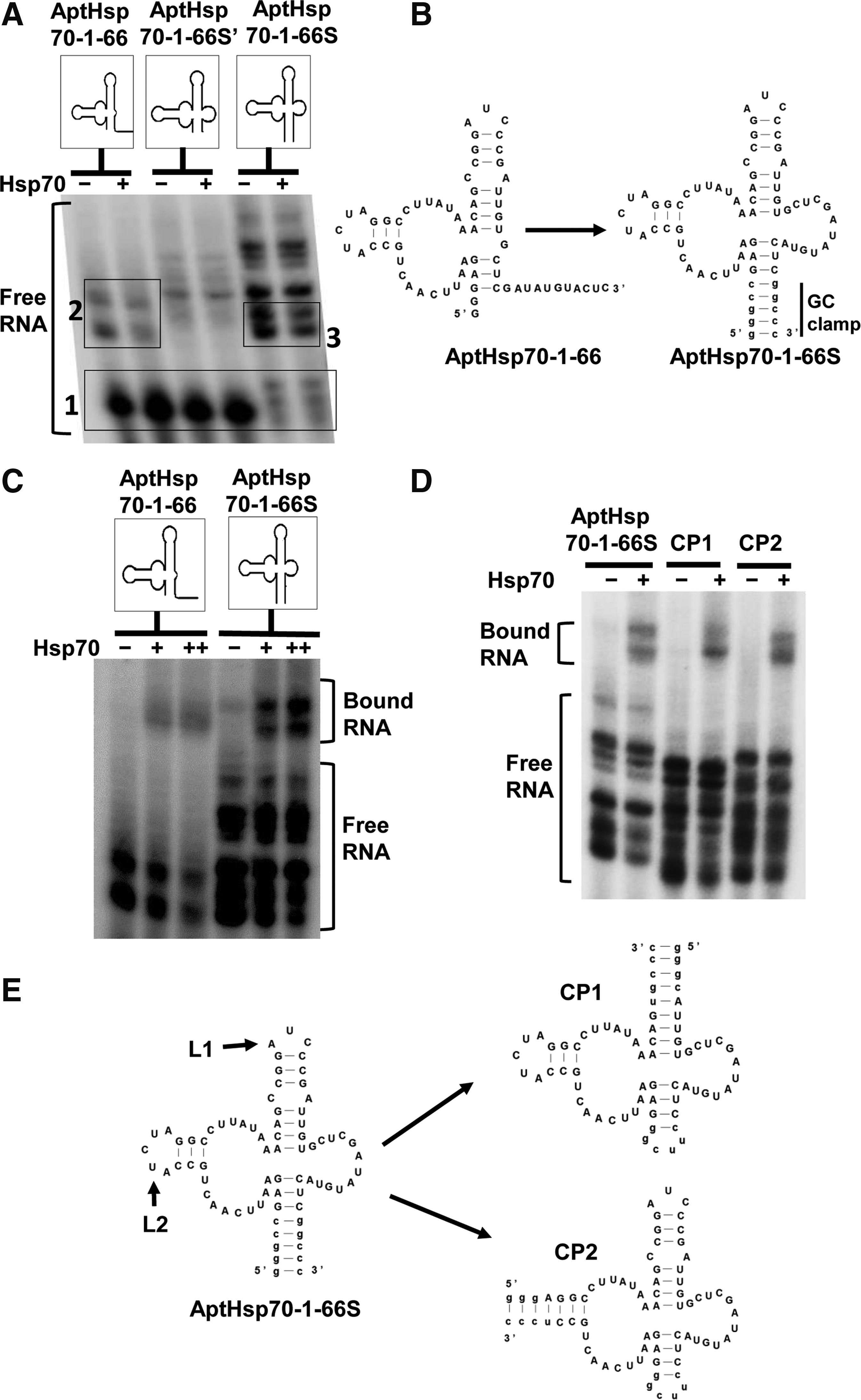
Refinement of AptHsp70-1.
When the new construct, AptHsp70-1-66S, was used in EMSA, there was an increase in the relative intensity of the RNA bands that bound to the protein (box 3 in Fig. 4A) and a corresponding decrease in the intensity of the faster migrating, protein-indifferent band (box 1 in Fig. 4A). Apparently, the improved binding of AptHsp70-1-66S was due to stabilization of the active conformation of AptHsp70-1-66. We further tested the predicted secondary structure of AptHsp70-1-66S by circular permutation. As shown in Figure 4D and 4E, introducing the chain breaking 5′/3′ ends to loop 1 or 2 (L1 or L2) resulted in the constructs CP1 and CP2, both of which retained binding to Hsp70, indicating that the aptamer adopted a complex 3-way junction structure. These mutational analyses not only yielded a compact version of the aptamer that can be used individually to perturb Hsp70 function, but also generated a structural interface with another aptamer to form composite aptamers [33]. Importantly, AptHsp70-1-66S retained conformational specificity to Hsp70-ATP (Supplementary Fig. S2).
Inhibitory effect of the aptamer on ATPase activity of Hsp70
We targeted Hsp70-ATP conformation for aptamer generation to find an RNA aptamer capable of influencing the ATPase activity of Hsp70. After confirming the conformation specificity of AptHsp70-1, we used single turnover ATPase assays to detect its effect on Hsp70's ATPase activity. In this assay, the ATP hydrolysis and nucleotide exchange steps can be separated and the step in the catalytic cycle affected by the aptamer can be revealed. Although Hsp70's ATP hydrolysis is essential for its chaperone function, the intrinsic ATPase activity is generally too low (ranging between 3×10–4 and 1.6×10–2 per second) to drive the functional cycle [34]. Tight coupling of ATP hydrolysis with substrate association often requires the stimulation of ATPase by J domain proteins (JDP, a.k.a. Hsp40), which are a diverse group of cofactors sharing a highly conserved ∼70 amino acid signature region, the J domain [35].
When we included a human Hsp40 (Hdj1) in our ATPase assay, we observed that AptHsp70-1 inhibited Hsp70 ATPase activity (Fig. 5A). For further characterization of aptamer activity we used the founding member of J domain protein family, E. coli Hsp40 (DnaJ) for two reasons. First, there are about 50 human J domain proteins [36], and among them, although Hdj1, Hdj2, and Hdj3 are the major isoforms, it is not clear which one is the physiologically right partner of human Hsp70 [37–40]. Second, the mechanism of Hsp40 is best understood for DnaJ [35], which is the only J domain protein in E. coli [39] and has been used as a generic Hsp40 in in vitro studies of human Hsp70 [15]. As shown in Fig. 5B, when AptHsp70-1 was incubated with Hsp70 and DnaJ in a single turnover ATPase assay, there is a significant decrease in ATPase activity, while nonspecific yeast tRNA had no effect. A dose response curve for this inhibition indicated an IC50 of 500 nM for the aptamer when 200 nM Hsp70 and 200 nM DnaJ was used in the assay (Fig. 5C). Since there was only one ATP turnover in the single turnover assay, and the aptamer bound only to Hsp70-ATP but not Hsp70-ADP, it must have specifically inhibited the ATP hydrolysis step in the ATPase cycle [27,41].
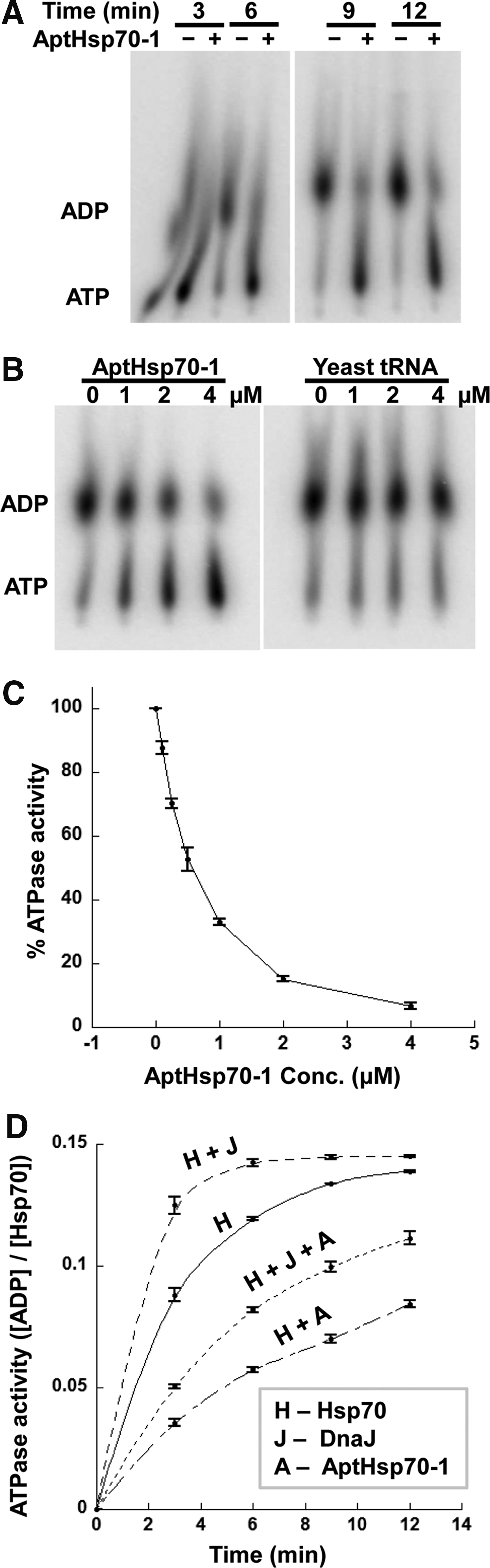
Inhibitory effect of AptHsp70-1 on Hsp70 ATPase activity.
We further studied the kinetics of this inhibitory effect and found that the aptamer mediated inhibition was not dependent on DnaJ. The aptamer decreased turnover rate of Hsp70 by 2.22 fold at 1:5 (Hsp70:AptHsp70-1) ratio in the absence of DnaJ and by 2.58 fold in the presence of DnaJ at 1:1:5 (Hsp70:DnaJ:AptHsp70-1) ratio (Fig. 5D and Table 3). Thus, the aptamer decreased the rate of both intrinsic and DnaJ stimulated ATPase activity of Hsp70, whereas DnaJ increased the rate of ATPase activity even in the presence of the aptamer (Fig. 5D and Table 3). This mutual independence also suggested that the aptamer binding site on Hsp70 was likely to be different from that occupied by DnaJ (or JDP/Hsp40 in general).
Discussion
Hsp70 is a major hub in the complex network responsible for the quality control of the proteome [42]. Chemical tools with distinct modes of action are necessary to complement genetic and biochemical methods in the mechanistic dissection of this network. The potential of Hsp70 as a therapeutic target has also motivated the search for its ligands, especially in the form of small molecules that could inhibit its function [12]. An outstanding and elegant example of this approach is the rational design of a small molecule interacting with an allosteric pocket discovered through extensive computational modeling based on detailed structural information [18]. Incidentally, a global druggability survey of this structural model also revealed the paucity of suitable sites for the development of small molecules. Of the five potential sites identified on the surface of the full-length human Hsp70 model, only one was deemed highly druggable [18], which partly explained the difficulty in finding drug leads for this target.
In contrast, we sought after a macromolecular ligand for Hsp70, which does not necessarily require a deep pocket to fit and has higher affinity than most small molecule ligands. In the approach we took, the only structural information utilized explicitly was the requirement of potassium ions for ATPase activity. With this information we designed a condition to fix the Hsp70-ATP conformation during in vitro selection and acquired a conformation-specific aptamer. Hsp70 undergoes extensive structural changes from its ATP-bound state to its ADP-bound state [5–7]. So, targeting one conformation (Hsp70-ATP) for aptamer selection was sufficient to obtain an aptamer specific for that conformation, which did not recognize the alternative conformation (Hsp70-ADP). Even though no negative selection was performed against Hsp70-ADP, no aptamer was isolated for this conformation. To our knowledge this is the first instance for such a selection and it would be interesting to see if this could be repeated for other proteins that undergo extensive conformational changes.
Hsp70 is a non-Walker ATPase, with its NBD and SBD conformations allosterically coupled [43,44]. The aptamer inhibited Hsp70 ATPase activity in single turnover ATPase assays, which indicates it inhibited the ATP hydrolysis step specifically in the ATPase cycle. The binding of the aptamer seems to stabilize the ATP bound conformation by slowing the catalytic step. Because it inhibited Hsp70's ATPase activity without preventing stimulation of the ATPase activity by Hsp40, the aptamer should have interacted with Hsp70 through a different site than the one recognized by Hsp40. This mechanistic insight may be useful in the development of novel drug leads that target this particular site on Hsp70. Since the ATPase activity is commonly used to drive ordered conformation changes of molecular machines that could function as motors [43,45–52], inhibiting this activity is an efficient strategy for drug targeting and therapy [22,53–55]. However, small compounds targeting ATPase often exhibit low specificity [56,57] and the use of reagents with larger surface area like aptamers may help overcome this problem. Therefore, aptamer mediated inhibition of Hsp70 ATPase activity can be an effective therapeutic strategy for treating Hsp70 associated diseases.
The heat shock response promotes cancer cell survival [58], and eliminating Heat Shock Factor 1 (HSF1), the transcription activator for the major heat shock genes including Hsp70, has been shown to be protective against cancer [58]. We had previously developed an RNA aptamer that interferes with the DNA binding of HSF1 [59] and inhibits HSF1 in human cancer cells [60]. Hsp70 also has oncogenic potential and its elimination has been shown to induce cancer cell death [61]. The Hsp70 aptamer described here inhibits Hsp70 ATPase activity and may work in synergy with the aptamer for HSF1 to suppress the proliferation of cancer cells. In addition, heat shock proteins are also linked to protein misfolding disorders such as cystic fibrosis and neurodegenerative diseases such as Alzheimer's [62–64]. Thus, specific inhibitory reagents to different proteins functioning at different stages of heat shock response will be useful tools to dissect the intricacies of this pathway and they will also have potential therapeutic applications.
Footnotes
Acknowledgment
This work was supported by a grant (R01CA140730) from the National Institutes of Health.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.