
Abstract
The bacterial cell wall presents a barrier to the uptake of unmodified synthetic antisense oligonucleotides, such as peptide nucleic acids, and so is one of the greatest obstacles to the development of their use as therapeutic anti-bacterial agents. Cell-penetrating peptides have been covalently attached to antisense agents, to facilitate penetration of the bacterial cell wall and deliver their cargo into the cytoplasm. Although they are an effective vector for antisense oligonucleotides, they are not specific for bacterial cells and can exhibit growth inhibitory properties at higher doses. Using a bacterial cell growth assay in the presence of cefotaxime (CTX 16 mg/L), we have developed and evaluated a self-assembling non-toxic DNA tetrahedron nanoparticle vector incorporating a targeted anti-blaCTX-M-group 1 antisense peptide nucleic acid (PNA4) in its structure for penetration of the bacterial cell wall. A dose-dependent CTX potentiating effect was observed when PNA4 (0–40 μM) was incorporated into the structure of a DNA tetrahedron vector. The minimum inhibitory concentration (to CTX) of an Escherichia coli field isolate harboring a plasmid carrying blaCTX-M-3 was reduced from 35 to 16 mg/L in the presence of PNA4 carried by the DNA tetrahedron vector (40 μM), contrasting with no reduction in MIC in the presence of PNA4 alone. No growth inhibitory effects of the DNA tetrahedron vector alone were observed.
Introduction
A
The current study developed a potential alternative delivery vehicle for synthetic antisense oligonucleotides; a novel delivery mechanism was based on self-assembling three-dimensional (3D) DNA tetrahedral structures. DNA is inherently non-toxic to cells [8], making it a highly suitable therapeutic delivery vector.
Setyawati et al. reported that a tetrahedral DNA structure was able to traverse the bacterial Gram-negative cell wall and enter the cytoplasm with no toxic effects of the structure alone [9]. Their DNA tetrahedron vector was designed to be self-assembling based on regions of complementarity between single-stranded DNA [10]. This DNA tetrahedron consisted of four 55-base ssDNA strands, each strand incorporating unique regions of complementarity with other strands, resulting in, by Watson–Crick base pairing, the folding of the strands to form a stable 3D equilateral tetrahedron.
Adapting this approach to the design and construction of 3D structures based on ssDNA, the present study developed a DNA tetrahedron vector incorporating a PNA into its structural design. This anti-blaCTX-M-group 1 PNA (PNA4) has been previously shown to translationally inhibit the expression of β-lactamase CTX-M-group 1 in field and clinical Escherichia coli isolates and to partially restore cefotaxime sensitivity in strains with a reduced susceptibility phenotype [11].
Materials and Methods
Strains
E. coli strain LREC461 of human origin was obtained from the Public Health laboratory, the United Kingdom, and had reduced susceptibility to CTX, which could be partially restored by PNA4 conjugated to a cell-penetrating peptide [11].
PNA4 antisense oligomer
The 13-mer anti-blaCTX-M-group 1 PNA4 was synthesized commercially by Cambridge Research Biochemicals (Cleveland, United Kingdom) with the base sequence of: ttccttattctgg (5′-3′).
Design of a 108 bp tetrahedron nanoparticle to carry a single 13-mer PNA (PNA4)
A DNA/PNA tetrahedron, designated PT01 (PNA4-carrying tetrahedron 01), was designed to self-assemble from three 54 nt single-stranded DNA (ssDNA) oligonucleotides (PT01-S1–3), one 41 nt ssDNA (PT01-S4) oligonucleotide, and one 13-mer PNA (Fig. 1A). Sequences of structural PT01 component ssDNA strands are as follows (5′–3′): PT01-S1: cgcgacttaggtccataatcaaggggccggtgagatgggagtgaacgggtctgg; PT01-S2: tagcgttaggacaacggaatctcaccggccccttgatacgtgcgggtctgataa; PT01-S3: ttatggacctaagtcgcgagtccagaataaggaactttatcagacccgcacgta; PT01-S4: actccagacccgttcactccctccgttgtcctaacgctaag.
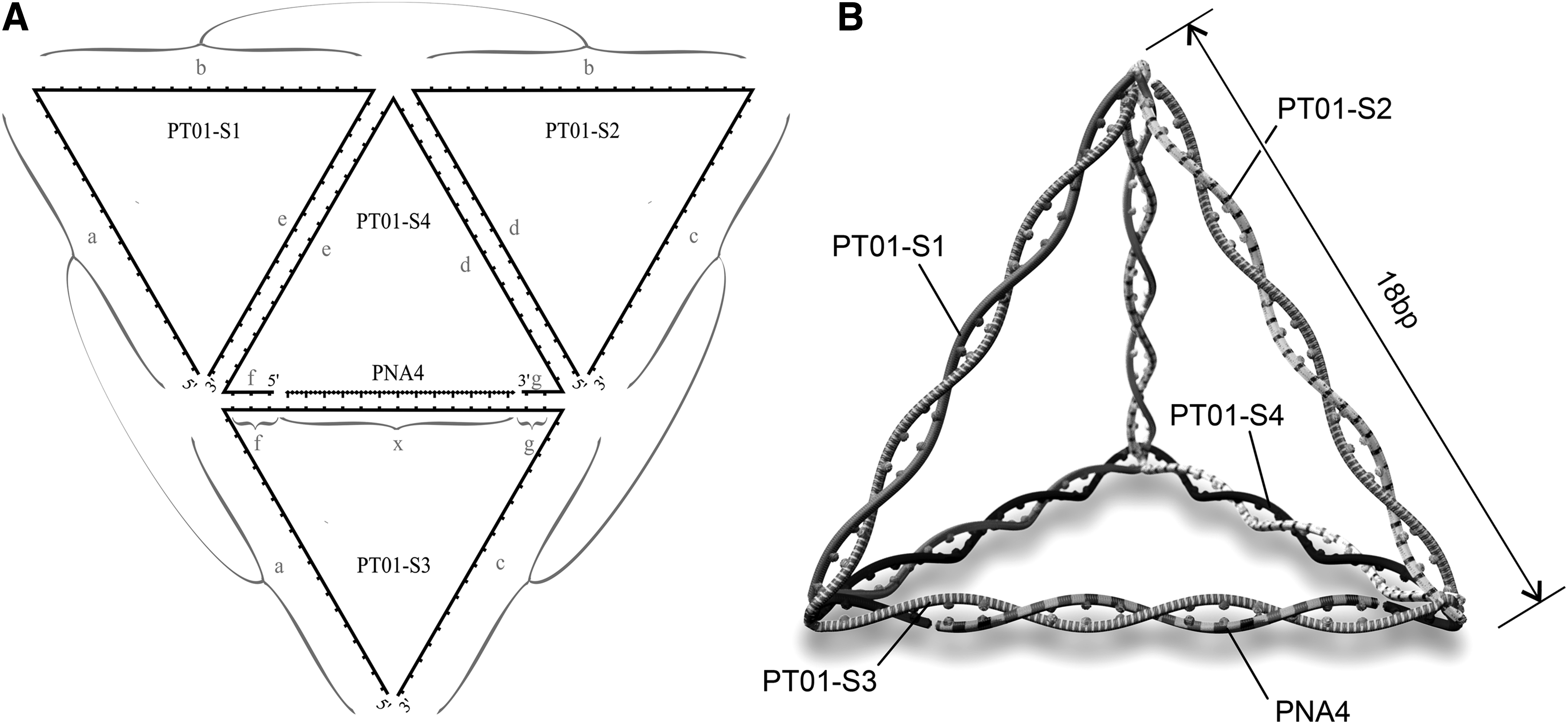
PT01 tetrahedron design to carry one 13-mer PNA.
Tetrahedron component ssDNA strands were checked for unwanted hetero and homo dimerization with Multiple Primer Analyzer [12]. Figure 1b is an illustrative representation of a DNA tetrahedron carrying a PNA after self-assembly.
Assembly of DNA tetrahedral PNA4 delivery vehicle
Using an adapted method described by Pei et al. [10], equimolar concentrations of ssDNA structural oligonucleotides and PNA4 were mixed in an annealing buffer (20 mM Tris, 50 mM MgCl2, pH 8.0). The mixture was heated to 95°C for 4 min and cooled rapidly on ice for 30 min. Assembly of the tetrahedron was verified by agarose gel electrophoresis.
Verification of assembly of DNA tetrahedral PT01 delivery vehicle by agarose gel electrophoresis
Assembly of the tetrahedron was verified on a 2% w/v TBE agarose gel, and 4–5 V/cm was applied until sufficient separation was observed. Individual DNA strands, and combinations of strands, for example, PT01-S1+PT01-S2, PT01-S1+PT01-S2+PT01-S3, and PT01-S1+PT01-S2+PT01-S3+PT01-S4+PNA4, were visualized by agarose gel electrophoresis (Fig. 2), and single discreet bands of increasing size were regarded as evidence of successful annealing and complete tetrahedron formation. A 100 bp DNA ladder (Promega) with markers at 100 bp intervals between 100 and 1000 bp was used to estimate sizes of combinations of tetrahedron component strands. A fully assembled tetrahedron was expected to yield a discreet band of around 108 bp.
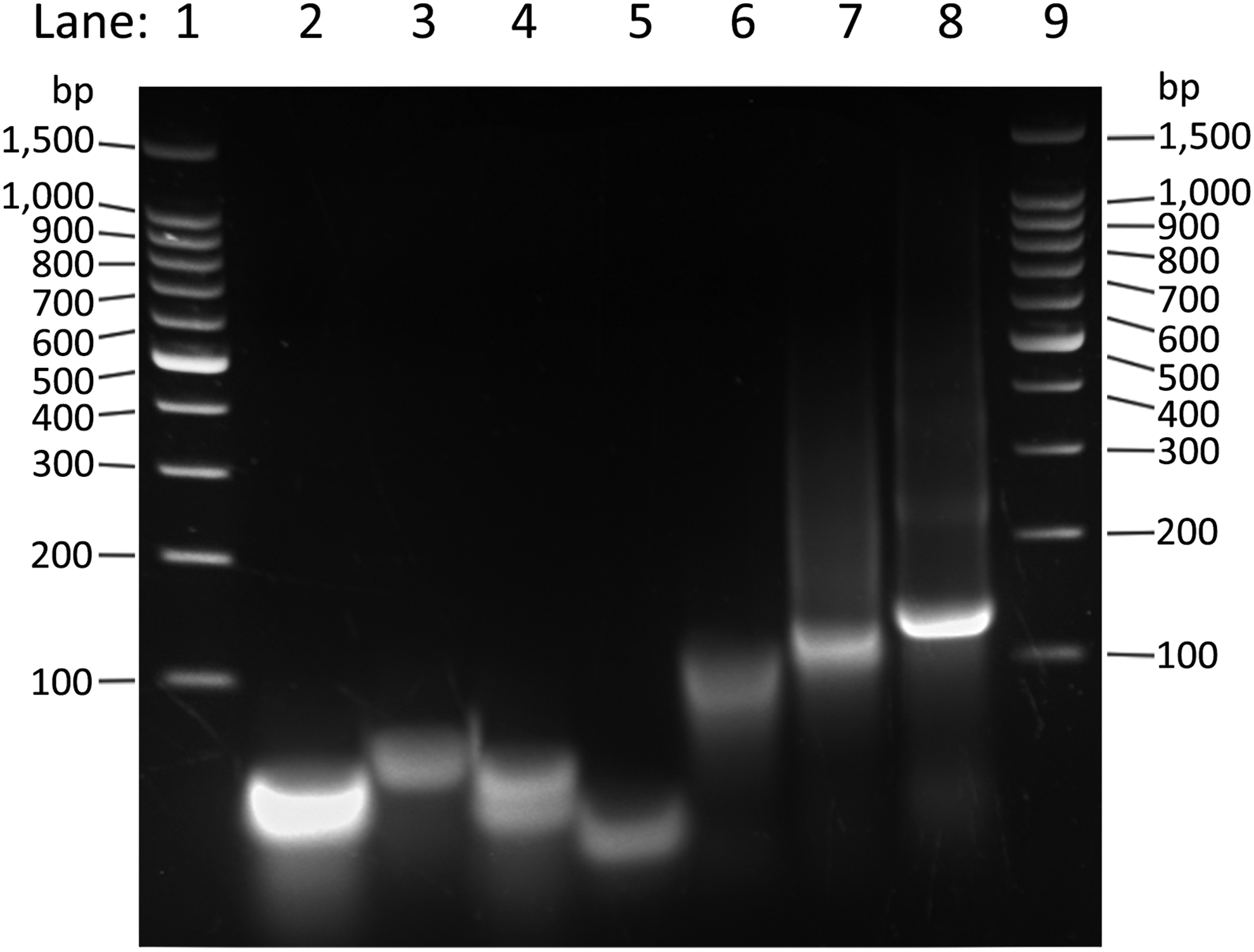
Agarose gel showing relative sizes of tetrahedron (PT01; 108 bp) component parts, partial assembly, and full assembly. Analysis of DNA fragment sizes separated on a 2% w/v TBE agarose gel, stained with SYBR® Safe DNA Gel Stain (Thermo Fisher Scientific, Inc.). Lane 1: 100 bp DNA ladder (Promega). Lane 2: PT01-S1. Lane 3: PT01-S2. Lane 4: PT01-S3. Lane 5: PT01-S4. Lane 6: PT01-S1+PT01-S2. Lane 7: PT01-S1+PT01-S2+PT01-S3. Lane 8: PT01-S1+PT01-S2+PT01-S3+PT01-S4+PNA4 (complete tetrahedron assembly). Lane 9: 100 bp DNA ladder (Promega).
Cell growth assay for assessing the activity of anti-blaCTX-M-15 antisense PNA oligonucleotides
A previously described E. coli cell growth assay in the presence of CTX was used for the evaluation of anti-blaCTX-M-group 1 activity of the PNA delivery DNA tetrahedron vector [11]. Briefly, bacterial cultures were typically grown from glycerol stocks at 37°C in a shaking incubator in MHB media containing CTX (2 mg/L) until early log phase (0.1–0.2 O.D.600nm) was achieved. The culture was diluted to achieve a final cell density of ∼100,000 CFU/mL, previously established empirically to be optimal for assessing the effect of antisense oligonucleotides, and for consistency across experiments. Diluted cell suspension (50 μL) was incubated with MHB (50 μL) growth medium that was supplemented with antibiotics and inhibitory agents where required, and it was transferred to a 96-well microtiter plate (Falcon). Bacterial cell growth was measured in a BMG Labtech FluoStar automated spectrophotometer (BMG LABTECH GmbH, Ortenberg, Germany), and optical density readings (600 nm) were taken over 18–24 h. The O.D.600nm readings were taken over 250 cycles, with 15 flashes per well per cycle approximately every 5 min. All replicates were independent cultures. Growth curves and statistical analyses were produced by using Prism® 6 software (GraphPad).
Establishment of minimum inhibitory concentration values for E. coli strains
E. coli strains were cultured in 96-well microtiter plates and incubated at 37°C. Growth was monitored at O.D.600nm for 18–24 h, and minimum inhibitory concentration (MIC) values were reported as the minimum concentration of an agent, or combination of agents, required to inhibit growth (O.D.600nm < 0.1) in a minimum of 50% of replicates after 18 h of incubation at 37°C.
Experimental controls used for the evaluation of inhibition of β-lactamase activity by unmodified and peptide-conjugated PNA4
Specificity of unmodified and peptide-conjugated PNA4 was demonstrated in previous studies by Readman et al. [11]: Briefly, anti-blaCTX-M-group 1 specific activity was initially demonstrated in a cell-free translation/transcription coupled system followed by cell growth studies with a CTX-resistant strain with a compromised cell wall. In field isolates, peptide-conjugated PNA4 control studies were undertaken with a cell-growth assay with an E. coli strain harboring blaCTX-M-14, with which PNA4 shared 15% (2 out of 13 bases) sequence complementarity. In the current study, control experiments were undertaken to demonstrate the lack of effect on growth of LREC461 to PT01 in the absence of CTX. Any effects of the tetrahedral structure itself were determined by cell-growth assays in the presence of a DNA control tetrahedron assembled from PT01 component ssDNA strands and a 13 nt ssDNA oligonucleotide of the same sequence as PNA4.
Results
Design and assembly of a PNA-carrying tetrahedral DNA nanoparticle
The sequence of ssDNA forming the structural frame of the 3D tetrahedral vector consisted of terminal areas on each strand that were complementary with regions on the other structural strands. These regions of complementarity facilitated the formation of the 3D double-stranded DNA tetrahedron structure. After self-assembly, this was designed to form a tetrahedron with a total size of 108 bp, each edge being 18 bp in length, and PNA4 carried by a region of complementarity with PT01-S3 (Fig. 1A). Assembly of the DNA tetrahedron PT01 was evaluated by visualization of migration and gel-shift patterns of the component oligonucleotides, and intermediate and final products of the tetrahedron annealing process. As shown in Fig. 2, the strands annealed to form progressively larger DNA complexes with slower electrophoretic migration patterns; the final annealed product comprising all components necessary for tetrahedron formation yielded a single discreet band, in a position on the gel that was consistent with a larger DNA molecule than the individual component parts.
Anti-blaCTX-M-3 activity of PT01
E. coli field isolate LREC461 was incubated in the presence of a combination of CTX (16 mg/L—previously shown to be a suitable concentration for observation of CTX potentiation by PNA4) and PT01 (0–30 μM), and a significant synergistic (fractional inhibitory concentration index <0.5) dose-dependent effect was observed (Fig. 3A), evidenced by the increasing time taken for the culture to achieve logarithmic growth. The MIC (to CTX) was reduced in the presence of PT01 (40 μM) from 35 to 16 mg/L (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/nat). Possible inherent toxicity of PT01 was evaluated by culturing LREC461 in the absence of CTX and in the presence of PT01 (10–30 μM), and no significant effects were observed (Fig. 3B). A control DNA tetrahedron was assembled, substituting a 13 nt ssDNA oligonucleotide sharing 100% sequence identity with PNA4 for PNA4, and assembled by using the components PT01-S1–4. No significant effects on growth were observed when field isolate LREC461 was cultured in the presence of the DNA control tetrahedron alone (10–30 μM; Fig. 3C). Small but significant (P < 0.05) negative effects on growth were observed when field isolate LREC461 was cultured in the presence of the DNA control tetrahedron (10 μM) and CTX (16 mg/L). No significant effect was observed with PNA4 without the DNA tetrahedron (Fig. 3D).
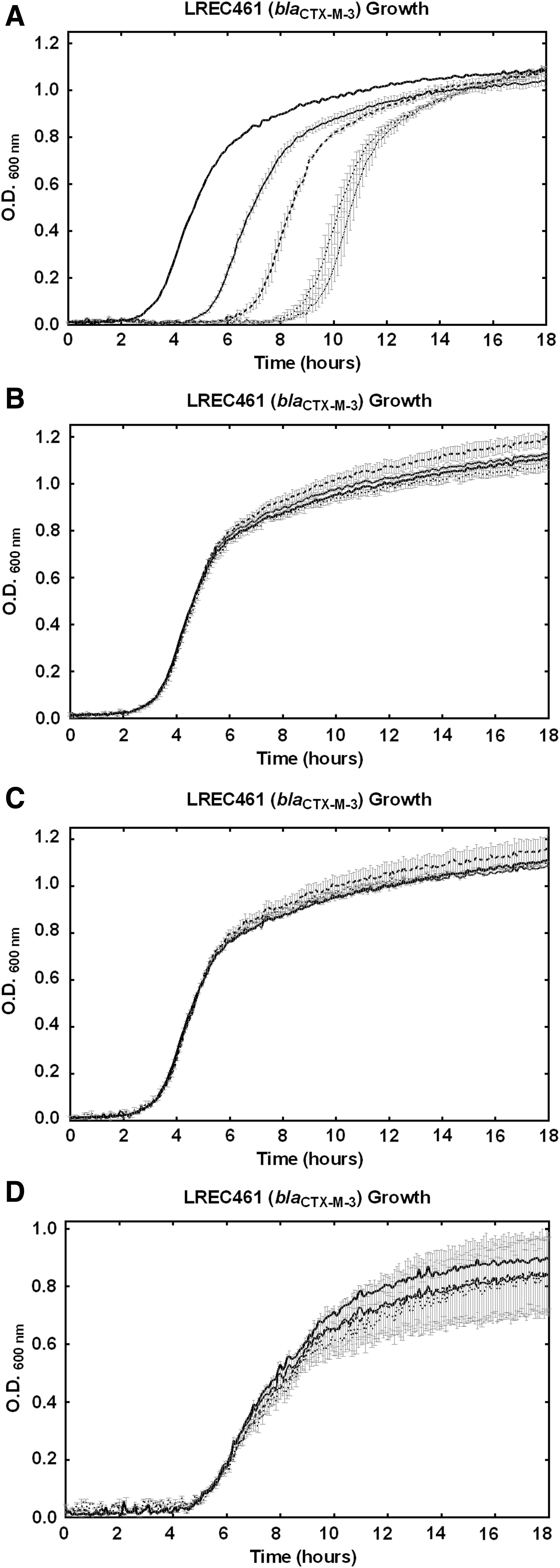
Effects of PNA4-carrying tetrahedron PT01 or DNA control tetrahedron in the presence and absence of CTX (16 mg/L) on growth of Escherichia coli strain LREC461 (harboring blaCTX-M-3).
Discussion
This study demonstrated the application of a DNA tetrahedron vector to deliver a 13-mer PNA antisense oligonucleotide within a bacterial cell by using a self-assembling complementary base pairing assembly approach. The delivery of the active synthetic antisense oligomer component of the tetrahedron relied on post-cell penetration dissociation of the PNA from the DNA structure, or the DNA components of the structure being degraded either wholly or partially by endonucleases on entering the bacterial cytoplasm, releasing the synthetic antisense agent from the DNA tetrahedron [9]. Setyawati et al. found that DNase I was sufficient to degrade a similar dsDNA structure in Staphylococcus aureus [9], whereas Li et al. found that such a structure was relatively stable in bovine serum albumin where DNase I concentrations were typically lower than intracellular concentrations [8]. The specific activity of PNA4 against blaCTX-M-15 had been previously demonstrated by Readman et al. [11]; this included the quantification of β-lactamase activity by an HPLC assay measuring the degradation of CTX in the presence and absence of PNA4 in a cell-free translation/transcription coupled system. Also, the cell-wall compromised mutant strain AS19 was transformed with a blaCTX-M-15-producing plasmid and cultured with CTX and the presence and absence of PNA4, and growth was measured by spectrophotometry over 18–24 h. As blaCTX-M-15 is a non-essential gene, the use of PNA4 in the absence of CTX provided a suitable control for the isolation of the effects of PNA4 on growth. Finally, a field isolate harboring blaCTX-M-14 to which PNA4 was not complementary was cultured with CTX in the presence and absence of PNA4 conjugated to a cell-penetrating peptide. This was the equivalent of a scrambled PNA control [11].
Assembly of the tetrahedral structures were verified by native agarose gel electrophoresis with a series of stepwise incremental DNA structural components. The number of nucleotides will not be related to the calibration standards because of the 3D nature of the partially and fully assembled tetrahedron; however, progressively slower migrating discreet bands represent evidence of successful assembly.
A control DNA tetrahedron was constructed, substituting ssDNA, of the same sequence, for the antisense oligonucleotide. There were no observable effects in field isolates incubated in the presence of a DNA control tetrahedron in the absence of CTX. In the presence of CTX and DNA control tetrahedron combination, a small increase in CTX sensitivity was observed, which was potentially attributable to opportunistic co-translocation of CTX with the control DNA tetrahedron. Control data (Fig. 3C) for the tetrahedron demonstrated no inherent antimicrobial properties of the structure. In contrast, CPPs do have well documented growth inhibitory and toxic effects in bacteria and mammalian cell cultures. CPPs may enable uptake by membrane perturbation, thereby having toxic effects through a loss of structural integrity and opportunistic co-translocation of extra-cellular material; CPPs have been shown to have a synergistic effect with certain antibiotics [4,7]. The observed lack of inherent toxicity or a CTX potentiating effect of the control tetrahedral structure may indicate that membrane perturbation was not the primary mechanism of cell penetration and may, therefore, implicate an alternative uptake pathway. Bacterial natural competence is well known to exist across a range of species, although not well characterized in E. coli. Bacteria commonly express DNA receptor sites at the cell surface, and Gram-negative bacteria such as Haemophilus parainfluenzae and Neisseria gonorrhoeae have been shown to recognize sequence-specific DNA fragments [13–15]. Mechanisms of DNA uptake in E. coli require further study, and it may be plausible to suggest that the shape and size of 3D DNA structures may enable cell penetration via an alternate hitherto unidentified pathway.
The observed re-sensitization of field isolates to previously sub-lethal concentrations of CTX suggest that a DNA tetrahedron is able to penetrate a bacterial cell wall and deliver its cargo, which is able to specifically bind to its target and inhibit CTX-M protein expression. No significant inhibitory effects were observed when field isolate LREC461 was treated with PNA-carrying tetrahedron PT01 in the absence of CTX. Further studies are required to optimize tetrahedron size and isolate the mechanism of penetration.
The CTX-potentiating effects of PT01 were smaller than the previously reported equivalent peptide-conjugated antisense oligonucleotides [11] that yielded a reduction in MIC (to CTX) in strain LREC461 from 35 to 8 mg/L in the presence of PNA4 conjugated to a cell-penetrating peptide with an amino-acid sequence of (KFF)3K (3.2 μM), potentially suggesting a comparatively lower efficient penetration efficiency of the tetrahedron. However, the cell-penetrating peptide portion of the CPP-antisense oligonucleotide conjugate ((KFF)3K) has been previously shown to have synergy with certain antibiotics [4,7], as well as some potential inherent toxicity, contributing to the overall growth inhibitory effect.
Further studies are required to determine the range of bacterial strains that are susceptible to antisense oligonucleotide-mediated protein expression inhibition carried out by DNA tetrahedrons.
Studies that are conducted to investigate the discriminatory potential of tetrahedral delivery vehicles would also be potentially advantageous—cell-penetrating peptides commonly lack specificity for any cell types, and their inherent toxicity to bacterial and mammalian cell cultures is well documented. A similar tetrahedral delivery vehicle was constructed and found to have no inhibitory effects against a mammalian cell culture; however, Walsh et al. reported substantial uptake of a DNA tetrahedral structure in mammalian cells [16]. Further investigations of the mechanism of uptake, and relative uptake efficiency would be required to elucidate this important factor. It is likely that shape, size, and nucleotide sequence of the structure will impact uptake efficacy in different cell types; a delivery vector that was specific to bacterial cells and did not have inherent negative effects on cell growth would be desirable characteristics for a targeted vector carrying antimicrobial agents.
With a demonstrated ability to both penetrate a bacterial cell wall and deliver an active targeted synthetic antisense oligonucleotide, this approach has a large range of optimization options, and a variety of potential applications. The low cost, simplicity, and speed of assembly would appear to make this a potentially viable alternative antisense oligonucleotide delivery vehicle.
Footnotes
Author Disclosure Statement
Research was conducted in the absence of any commercial or financial relationships, which could be construed as a potential conflict of interest; therefore, no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.