
Abstract
Herein we described the synthesis of siRNA-NES (nuclear export signal) peptide conjugates by solid phase fragment coupling and the application of them to silencing of bcr/abl chimeric gene in human chronic myelogenous leukemia cell line K562. Two types of siRNA-NES conjugates were prepared, and both sense strands at 5′ ends were covalently linked to a NES peptide derived from TFIIIA and HIV-1 REV, respectively. Significant enhancement of silencing efficiency was observed for both of them. siRNA-TFIIIA NES conjugate suppressed the expression of BCR/ABL gene to 8.3% at 200 nM and 11.6% at 50 nM, and siRNA-HIV-1REV NES conjugate suppressed to 4.0% at 200 nM and 6.3% at 50 nM, whereas native siRNA suppressed to 36.3% at 200 nM and 30.2% at 50 nM. We could also show complex of siRNA-NES conjugate and designed amphiphilic peptide
Introduction
N
siRNA is a double stranded RNA with 21 nt length and can efficiently suppress mRNA or lncRNA by RISC dependent cleavage of the target in a sequence specific manner. Synthetic miRNA can enhance or suppress the function of endogenous miRNA by regulating mRNA or lncRNA in a RISC dependent manner. Ribozyme and DNAzyme function as sequence specific endonucleases against target primary transcript, mRNA and lncRNA. DNA/RNA aptamers and DNA/RNA decoys are usually screened through in vitro selection and function as competitive ligands or inhibitors for receptors, enzymes, transcription factors, RNA, DNA, and so on.
In the past decade and a half, a number of novel noncoding RNAs have been discovered by comprehensive analysis of transcripts and their crucial functions have been revealed step by step. It has been found that upregulation or downregulation of a specific miRNA expression is seriously related to various diseases, including cancers and neural disorders. lncRNA is another novel noncoding RNA involved in transcriptional and posttranscriptional regulation of genes, including epigenetic regulation. Therefore, the structure and the intracellular location of the target molecules for nucleic acid drugs are getting diverse more and more. The target molecules include genomic DNA, lncRNA, nuclear miRNA, and pre-mRNA in the nucleus and mRNA and miRNA in the cytoplasm. Proteins which contribute genetic regulation, metabolism, and signal transduction can be targets for DNA/RNA aptamers and decoys all over the cell.
Although medical applications of synthetic nucleic acids are extremely promising, a number of hurdles still need to be cleared to exert the full potential of them. Nucleic acid drugs should (1) be stable enough chemically and biologically, inside and outside the cells, (2) be delivered to the targeted tissue, (3) be taken up into cells effectively, (4) be localized in the proper intracellular site, (5) bind to the target molecule with high affinity and specificity, (6) not be toxic, (7) not cause undesirable effect (off-target effect), (8) not induce immune response, and (9) not interact with intracellular and extracellular molecules in a nonspecific manner [8–12].
To clear these hurdles, chemical conjugation of oligonucleotide with biological and synthetic functional molecules by a covalent linkage has been attracting intensive attention more and more and now is recognized as one of the most powerful strategies along with chemical modification and noncovalent complexation [13–16]. Oligonucleotide conjugates have been verified to improve the pharmacokinetic properties, the stability in vivo and in cells, the specificity of targeting tissues, the permeability into cells, the controlled intracellular trafficking with an aid of covalently linked peptides, lipids, sugars, protein ligands, RNA aptamers, antibodies, polyethylene glycol, and so on. Especially, since peptides derived from a great variety of proteins have extremely attractive and diverse structures and functions, oligonucleotide-peptide conjugates have been studied most intensely from a synthetic, a biological, and a medical point of view.
In our previous study, we reported the synthesis of oligonucleotide conjugates with nuclear localization signal (NLS) peptides, nuclear export signal (NES) peptides, designed peptides, and galactosamine by our original solid phase fragment coupling (SPFC) method [17]. SPFC is a convenient and versatile method and enables us to prepare DNA/RNA conjugates with peptides bearing arbitrary amino acid components and sequences. We successfully showed that oligoDNA-NLS peptide conjugates were taken up into cells without any transfection agent and localized in the cellular nucleus of human T lymphocyte cell line Jurkat. In contrast, oligoDNA-NES peptide conjugates were also taken up into Jurkat cells and localized in the cytoplasm outside the nucleus (Fig. 1). The controlled subcellular localization of oligonucleotides largely affected its antisense inhibition effect against human telomerase template RNA, that is, phosphorothioate DNA-NLS conjugate containing a peptide derived from Simian Vacuolating Virus 40 large T antigen (SV40 Tag) suppressed 99.6% of telomerase activity in 24 h and 95.3% in 48 h at 5 μM, whereas oligoDNA-NES conjugate derived from HIV-1 REV showed absolutely no inhibitory effect [18, 19].
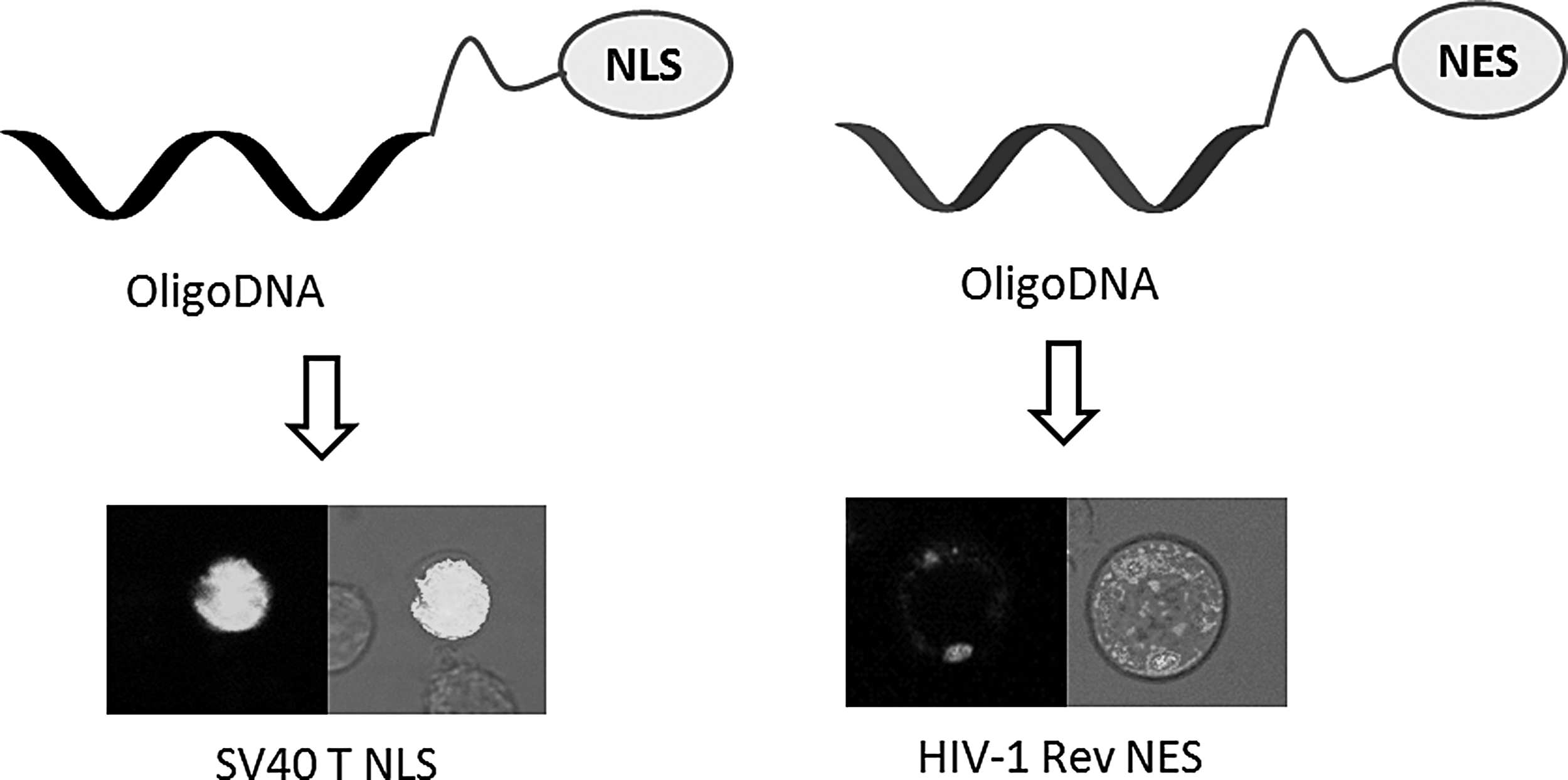
Controlled intracellular localization of oligoDNA-signal peptide conjugates.
In the present study, we prepared two types of siRNA-NES conjugates by our original SPFC and evaluated silencing efficiencies of siRNA-NES conjugates against bcr/abl chimeric gene in human chronic myelogenous leukemia cell line K562.
Materials and Methods
General remarks
Oligonucleotides were assembled on controlled pore glass (CPG) support by automated DNA/RNA synthesizer nS-8 II (Gene Design, Inc.) using a standard cyanoethylphosphoramidite chemistry. Modification of 5′ end of oligonucleotides was performed with N-MMT-2-(2-aminoethoxy) ethyl phosphoramidite (Glen Research 5′-Amino Modifier 5). Fmoc protected amino acids for peptide synthesis were purchased from Nova Biochem and Watanabe Chemical Industries, Ltd. Peptides were synthesized by Biotage® Initiator+ Alstra microwave assisted peptide synthesizer using a standard Fmoc chemistry on Wang-Resin (Nova Biochem). Oligonucleotides and conjugates were purified and analyzed by reversed phase high performance chromatography (RP-HPLC) on Hewlett-Packard HP-1100 system. RNA-NES conjugates and peptides were characterized by MALDI-TOF MS on Voyager-DE (Applied Biosystem) and ESI-MS on amaZon SL (Bruker Co.). Other chemicals for the synthesis of RNA-NEA conjugates were purchased from Wako, Kishida, Dojindo, and Aldrich. Total RNA extraction was performed with RNeasy Mini Kit (QIAGEN). Quantitative reverse transcription polymerase chain reaction was performed using SuperScript III Platinum One-step Quantitative RT-PCR System™ purchased from Life Technologies Co. on MX3005P (Agilent Technologies).
Peptide synthesis
Preparation of peptides was carried out by solid phase fmoc chemistry. The following side chain protected amino acids were used to obtain sufficient yield: Fmoc-Arg(Mtr)-OH, Fmoc-Lys(Boc)-OH, Fmoc-Asn(Trt)-OH, Fmoc-Glu(OtBu)-OH, Fmoc-Thr(tBu)-OH, and other neutral amino acids were used as Fmoc-AA-OH. HOBt (N-hydroxybenzotriazole), TBTU (2-(1H-benzotriazole-1-yl)-1,1,3,3-tetra-methyluronium tetrafluoroborate), and DIEA (diisopropylethylamine) were used as coupling reagents for peptide elongation in dry N, N-dimethylformamide (DMF).
Peptide synthesis was carried out on Biotage Initiator+ Alstra microwave assisted peptide synthesizer, using Wang resin with a 0.05 mmol scale. The protocols of peptide synthesis included the following cycle: deprotection of Fmoc was carried out by 20% piperidine for 5 min (this procedure was repeated twice); the external piperidine was removed by DMF; five equivalents of amino acid, activated with TBTU and HOBt in 1 M DIEA, were coupled with amino acid assembled on the resin for 30 min; nonreacted amino acid was capped by anhydrous acetic acid. Cleavage and side chain deprotection (except for Lys(Tfa)) were accomplished with a mixture containing 1 M TMSBr/thioanisole in TFA with m-cresol/EDT for 2–8 h (depending on the peptide sequence), at room temperature with constant stirring. The crude peptide fragment was obtained by precipitation in cold ether. The crude peptide fragments were purified by RP-HPLC on oligoR3 (4.6/100 mm) peek column (PerSeptive Biosystems), at linear gradient from 100% A to 100% B for 45 min at a flow rate of 1.0 mL/min. Eluent A contains 0.1% trifluoroacetic acid in water, and eluent B contains 0.1% trifluoroacetic acid in acetonitrile. The products were characterized by RP-HPLC and MALDI-TOF MS. The data of RP-HPLC profiles and MALDI-TOF MS spectra of NES peptides
Synthesis of RNA-NES peptide conjugates C1 and C2
The RNA-NES conjugates were synthesized by SPFC (Fig. 2). Oligonucleotides assembled on CPG (0.2 μmol scale) support in column using a standard cyanoethylphosphoramidite chemistry were reacted with 0.1 M N-MMT-2-(2-aminoethoxy)ethyl phosphoramidite in dry CH3CN containing 1H-Tetrazole. 1H-Tetrazole was used as an activator to attach an amino group to the 5′ ends of the oligonucleotides. The column was washed five times by dry CH3CN. MMT was removed by treatment of the column with 3% trichloroacetic acid (TCA) in dichloromethane. Then the amine was reacted with 50 eq of carbonyl diimidazole (CDI) in CH3CN containing 4 eq of DIEA for 2 h at room temperature in dry conditions. CDI was chosen as a most appropriate linker in this study. The column was washed carefully with dry CH3CN to remove external linker. Twenty equivalent amounts of peptide fragments, dissolved in 300 μL of dry DMF containing 10 eq of DIEA, were added to the CPG support, activated at 5′ end by CDI.

Synthesis of RNA-NES conjugates by solid phase fragment coupling. NES, nuclear export signal.
The CPG column, containing peptide fragment, was shaken for 24 h at 30°C under dry conditions and then washed thrice with 1 mL of DMF and five times with 1 mL of CH3CN. The CPG column was dried by N2-gas, and RNA-NES conjugates were cleaved and deprotected by treatment with concentrated aqueous ammonia for 4 h at 55°C in one step. RP-HPLC purification of RNA-NES conjugates was performed on an ODS column (5 μm, 4.6 × 150 mm) at 60 min linear gradient from 10% to 100% B at a flow rate of 0.2 mL/min. The mobile phase A contains 20 mM TEAA in water and phase B contains 20 mM TEAA in CH3CN/H2O (70/30).
In SPFC chemistry, the functional side chain of amino acid in peptide fragment could be free during conjugation, except for Lys(Tfa). Lys(Tfa) could be deprotected during the treatment of DNA/RNA-peptide conjugates assembled on CPG column with concentrated aqueous ammonia used for DNA cleavage and deprotection.
The products were characterized by RP-HPLC and MALDI-TOF MS. The data of RP-HPLC profiles and MALDI-TOF MS spectra of RNA-NES conjugates are shown in Supplementary Figs. S3–S6.
Cell culture and transfection
K-562 derived from chronic myelogenous leukemia was cultured in RPMI-1640 medium, supplemented with 10% heat-inactivated fetal bovine serum, streptomycin (100 μg/mL), and penicillin (100 U/mL) in a humidified atmosphere at 37°C over 5% CO2. The cells used for transfections were at ∼70% confluence.
For transfection of siRNA-NES conjugates: 24–48 h before the treatment, the cell suspensions were cultured in 24-well plates (Iwaki, 1 mL cell suspension per plate) at a density 5 × 105 cells/mL in RPMI-1640 medium without antibiotics. siRNA-NES conjugates were added to the plates in final concentrations of 1 and 5 μM, respectively. The mixtures were incubated for 24 and 48 h at 37°C in humidified atmosphere.
For transfection of siRNA-NES conjugates by Lipofectamine 2000™ (Invitrogen): siRNA-NES conjugates were diluted in 50 μL Opti-MEM (Invitrogen) and were incubated for 5 min at room temperature. Two microliters Lipofectamine 2000 were diluted in 48 μL Opti-MEM and incubated for 5 min at room temperature. Then both siRNA-NES conjugate and Lipofectamine 2000 solutions were combined together and incubated for 30 min at room temperature.
A hundred microliters of the RNA-NES conjugate-Lipofectamine 2000 complexes was added to 24-well plates, containing 900 μL of K562 cells (5 × 105 cells/mL) in RPMI-1640 medium without antibiotics. The incubation was carried out following the manufacturer's instruction. The final concentrations of RNA-NES conjugates were 1 or 5 μM.
Quantitative analysis of BCR/ABL gene expression by real time reverse transcription polymerase chain reaction
Total RNA was extracted using RNeasy Plus Mini Kit (QIAGEN). Single step RT-PCR was performed using SuperScript III One-Step RT-PCR with Platinum® Taq Kit (Invitrogen Life Technologies Corp.). The conditions for RT-PCR were as follows: 15 min at 50°C, 2 min at 94°C, and then followed by 40 cycles of amplification for 15 s at 94°C, 20 s at 55°C, and 20 s at 72°C. GAPDH was used as an internal control to ensure accuracy. The assay used an instrument capable of measuring fluorescence in real time (MX3005P; Invitrogen).
BCR/ABL mRNA Primers
Forward 5′-TGCAGATGCTGACCAACTCG-3′
Reverse 5′-GTTCCAACGAGCGGCTTCAC-3′
TaqMan Probe 5′-FAM-CAGTAGCATCTGACTTTGAGCCTCAGGGTCT-BHQ-3′
The results in Figs. 3–8 are the means of triplicate determinations. Each determination was calculated from triplicate measurements. Each error bar indicates a standard deviation of the triplicate determinations.

Silencing of BCR/ABL in K562 by 5′-modified siRNA. siRNA mediated reduction of BCR/ABL mRNA expression in K562 cells. siRNAs were transfected by Lipofectamine 2000™. Normalized BCR/ABL/GAPDH mRNA levels were measured 24 h after transfection. The results are the means and standard deviations of triplicate determinations.

Silencing of BCR/ABL mRNA by

Silencing of BCR/ABL mRNA by
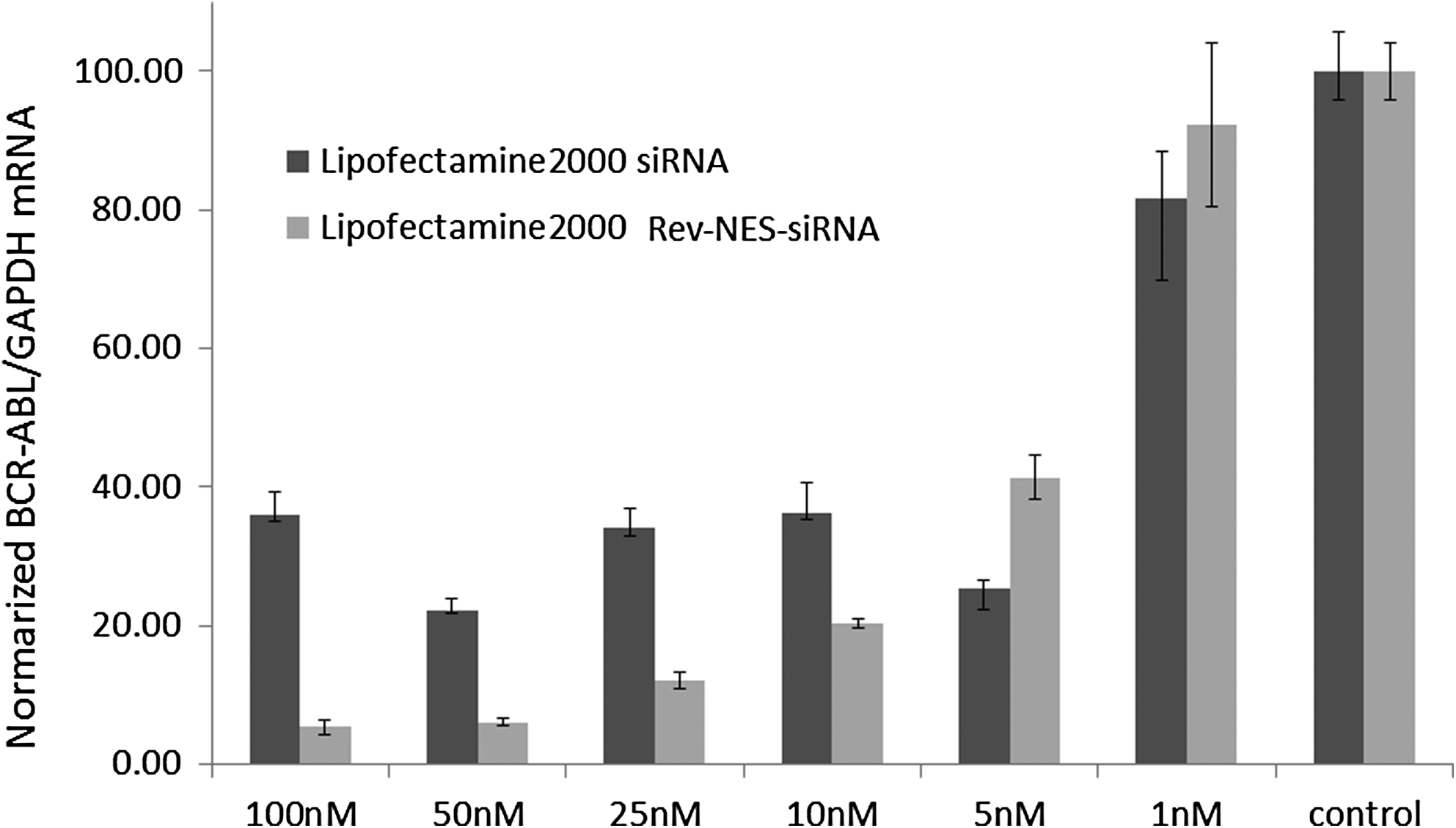
Silencing of BCR/ABL mRNA by

Silencing of BCR/ABL mRNA by
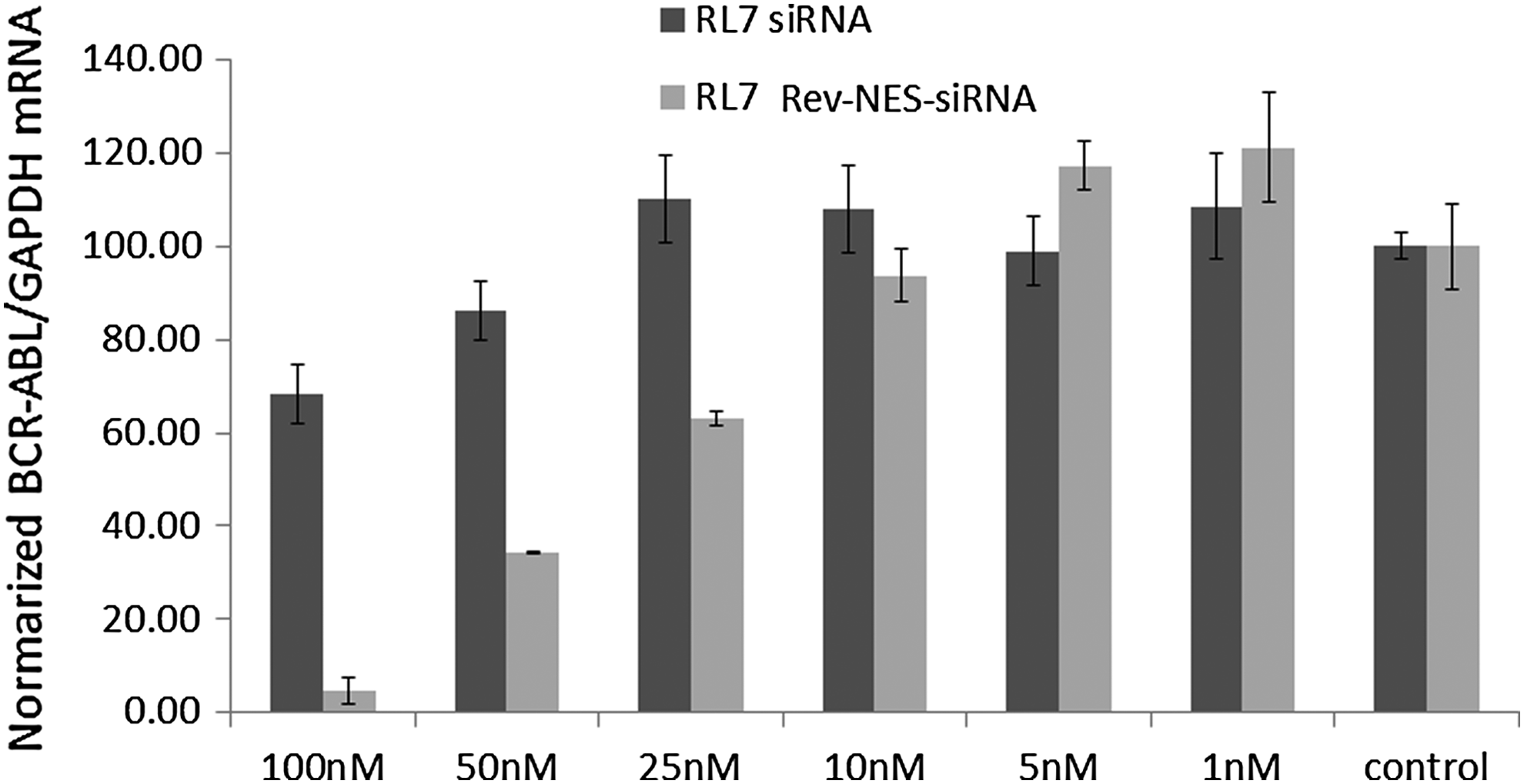
Silencing of BCR/ABL mRNA by
Results and Discussion
Silencing of BCR/ABL chimeric gene by siRNAs bearing 5′-Amino Modifier 5
As a preliminary study to find out the desirable site for conjugation, before the synthesis of siRNA-NES conjugates, silencing efficiencies against bcr/abl chimeric gene were evaluated using
Target sequence of BCR/ABL mRNA (355–390)
5′-ggauuu
antisense 3′-TTAGAGCGAACCCGCUCUCAUU-5′
antisense 3′-TTCGUCUCAAGUUUUCGGGAA-5′
antisense 3′-TTCGUCUCAAGUUUUCGGGAA-5′
antisense 3′-TTCGUCUCAAGUUUUCGGGAA
antisense 3′-TTCGUCUCAAGUUUUCGGGAA
X
Silencing efficiencies of

5′-Terminus of the guide strand modified with X in the cationic pocket of MID domain of human Argonaute2.
Synthesis of RNA-NES conjugates C1 and C2
According to the results mentioned above, two types of NES peptides were covalently attached to the 5′ end of the sense strand of siRNA by SPFC as shown in Fig. 2.
The sense strand of siRNAs targeting to BCR/ABL mRNA (361–381) with two thymidines at 3′ end over hang region was assembled on CPG by standard automated protocols of cyanoethylphosphoramidite chemistry and modified at 5′ end with 5′-Amino Modifier 5 (Glen Research). After removal of MMT protection by 3% TCA, solution of N, N-diisopropylethylamine (DIEA) and CDI in absolute acetonitrile was added to the CPG column using a syringe at room temperature and left the column placed on a shaker for 2 h. Solution of 10 equivalents of a peptide fragment which was separately prepared by solid phase synthesis and purified by HPLC and 10 equivalents of DIEA in absolute acetonitrile was added to the CPG column using a syringe at room temperature and left the column stand for 24 h. The TFIIIA NES peptide fragment (NH2CH2CH2CH2CO-LPVLENLTL-OH) derived from Xenopus laevis [22–24] used in the present synthesis has γ-aminobutyric acid (GABA) moiety with free reactive amino group at its N-terminus, and TFIIIA NES peptide fragment was linked to the 5′ end of RNA through GABA amino group. The HIV-1 REV NES peptide fragment (Ac-LPPLERLTL-KG-OH) [25, 26] has acetyl cap on N-terminal leucine and extra unprotected lysine and glycine at its C-terminus. HIV-1 REV NES peptide fragment was linked to RNA through ɛ-amino group of lysine with an adequate nucleophilic reactivity. The side chain of arginine contained in the NES peptide fragment derived from HIV-1 REV protein was set free during deprotection procedure using TFA before the conjugation step. The pKa value of DIEA is 10.76, which is slightly higher than the pKa value 10.67 of amino group of lysine and much lower than the pKa value 12.50 of guanidinyl group of arginine. That is why, it can be supposed that arginine side chain is protonated and has a decreased nucleophilicity, whereas lysine side chain is not protonated to undergo nucleophilic attack to isocyanate group in aqueous solution. In fact, HPLC profile of the isolated product was very sharp indicating a pure sole product. Cleavage from CPG support was performed with 28% aqueous ammonium hydroxide at 55°C for 4 h. The crude product was purified by reversed phase HPLC, and pure products were obtained in 26.8% for
Silencing of BCR/ABL chimeric gene by siRNA-NES conjugates
Using thus obtained two types of RNA-NES peptide conjugates
antisense; 3′-TTCGUCUCAAGUUUUCGGGAA-5′
NES; -HNCH2CH2CH2CO-LPVLENLTL-OH (TFIIIA NES)
antisense; 3′-TTCGUCUCAAGUUUUCGGGAA-5′
NES; Ac-LPPLERLTL-K(ɛNH-)G-OH (HIV-1 REV NES)
As in Fig. 4 for
Transfection of siRNA-NES conjugates by designed peptides
We understand that nontoxic transfection of siRNAs into cells is one of the most crucial technologies to be established for in vivo use and medical applications. Next we explored transfection of siRNA-NES conjugates by designed peptides which could bind to and internalize RNA into cells. In our previous studies, we found out amphiphilic designed peptides which could bind to double stranded DNA and RNA [26–29]. We investigated various types of designed peptides for the transfection of siRNA. We could find out that two peptides efficiently internalized the siRNA-NES conjugates into cells. One was named
Conclusions
Herein we successfully showed that siRNA-NES conjugates suppressed bcr/abl chimeric gene in human chronic myelogenous leukemia cell line K562 very efficiently. It can be reasonably speculated that NES peptide linked to 5′ end of sense strand of siRNA played a crucial role for the remarkable enhancement of silencing efficiency. Moreover we could show combination of siRNA-NES conjugate and
Footnotes
Acknowledgments
This work was financially supported by JSPS KAKEN grant number 22550159. D.A.S. is supported by RSF grant no. 14-44-00068. A.A.F. acknowledges support from RFBR grant no. 16-03-01055.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.