
Abstract
RNA interference (RNAi) offers great promise in life science research and therapeutic development, as it easily achieves a potent target gene knockdown with high specificity. Since the conventional small interfering RNA (siRNA) structure, known as 19 bp double-stranded RNA (dsRNA) with 2-nucleotide (nt) 3′ overhang, has been introduced to successfully elicit the RNAi in mammalian cells, a variety of structural variants of RNAi trigger have been developed. Our group previously reported branched, tripodal interfering RNA (tiRNA) structures as a multigene targeting RNA structure inducing RNAi. However, the immune stimulatory effect of branched tiRNA structure has not been thoroughly evaluated. In this study, we show that tiRNA with blunt ends triggers innate immune response in T98G cell and mouse macrophage cells, which is dependent upon the retinoic acid-inducible gene I (RIG-I), a well-known cytoplasmic dsRNA sensor. Interestingly, immune response triggered by tiRNA can be suppressed by the introduction of 2-nt 3′ overhang structure. Our finding expands the structural diversity of RIG-I ligands and provides a guide to develop a safe multitargeting RNA structure for therapeutic application.
Introduction
RNA interference (RNAi) is a sequence-specific gene silencing phenomenon in cells, which is initiated by double-stranded RNA (dsRNA) [1,2]. Abnormal expression of various cellular proteins, including transcription factors, subsets of kinases, and other signaling proteins can be regulated by RNAi [3]. Since the initial reports by Tuschl and colleagues of small interfering RNA (siRNA), which consists of 19 bp dsRNA with 2-nucleotide (nt) 3′ overhang on both ends, showing efficient target messenger RNA (mRNA) degradation in mammalian cells, siRNA has been considered as the standard RNAi trigger for basic life science research as well as therapeutic use [4]. However, a number of follow-up studies have shown that a variety of RNAs, whose structures deviate from original siRNA, can also trigger potent and specific RNAi [5]. In particular, we have previously reported a novel tripodal RNA structure termed tiRNA (tripodal interfering RNA), which simultaneously inhibits expression of three genes based on RNAi mechanism [6].
One of the major problems associated with dsRNA molecules inducing RNAi for therapeutic application is that they are recognized by specific pattern recognition receptors (PRRs) as pathogen-associated molecular patterns (PAMPs), resulting in the activation of innate immune system. Among various PRRs, retinoic acid-inducible gene I (RIG-I) and melanoma differentiation-associated protein 5 (MDA5), which are members of RIG-I-like receptors (RLRs), are responsible for recognizing foreign RNA, such as viral RNA in cytosol [7,8]. They both possess N-terminal caspase recruitment domain (CARD), DExD/H helicase domain, and C-terminal domain (CTD). When the dsRNA produced by viral RNA replication binds to CTD, CTD is dissociated from CARD to make an open conformation of RIG-I. The exposed CARD interacts with CARD of mitochondrial antiviral-signaling protein (MAVS), initiating innate immune activation signaling through the activation of NF-κB and IRF3. Phosphorylation of NF-κB and IRF3 drives the expression of type I interferon and related genes such as interferon stimulating genes (ISGs), including IFIT family [9–11].
Despite the similarity of structure between RIG-I and MDA5, these two receptors respond to different RNA structures. The 5′ triphosphate (5′ ppp) moiety possessing RNA is a primary RIG-I ligand [8]. In addition, chemically synthesized dsRNAs ranging from 21 to 27 bp with blunt ends and short polyinosinic–polycytidylic acid sodium salt (poly I:C) can trigger the immune response by activating RIG-I [12,13]. In contrast to RIG-I, MDA5 recognizes the internal region of longer dsRNAs, and the CTD of MDA5 forms a tighter and closer binding to dsRNA than that of RIG-I [14].
Previously, we reported a tiRNA structure that efficiently inhibited target gene expression by inducing RNAi-mediated mRNA cleavage; however, in that study, we have not thoroughly evaluated the activation of immune response triggered by tiRNA structure [6]. In this study, we assess the activation of innate immune response triggered by tiRNA in T98G cells and mouse macrophage cells. We show that tiRNA structure with blunt ends triggers innate immune response in T98G cells and mouse macrophage cells, initiating the immune response through mainly IRF activation, which is dependent upon the RIG-I. We then show that the immune response triggered by tiRNA can be suppressed by the introduction of 2-nt 3′ overhang structure. We also define the minimal length of dsRNA region within tiRNA structure that is required to activate the RIG-I-mediated immune response.
Materials and Methods
RNAs
Chemically synthesized RNA oligonucleotides were purchased from Dharmacon, and each single-strand oligonucleotide was purified by high-performance liquid chromatography. siRNAs were purchased from Bioneer and Dharmacon. The sequence details of siRNAs and tiRNAs used in these experiments are shown in Supplementary Table S1.
Cells culture and transfection
HeLa (CCL-2) cell line was purchased from ATCC and cultured in Dulbecco's Modified Eagle's Medium (Corning) supplemented with 10% Fetal Bovine Serum (Gibco), 100 U/mL penicillin, and 100 μg/mL streptomycin. T98G (CRL-1690) cell line and Hepa 1–6 (CRL-1830) were also purchased from ATCC and cultured in Dulbecco's Modified Eagle's Medium (Gibco) supplemented with 10% Fetal Bovine Serum (Gibco), 100 U/mL penicillin, and 100 μg/mL streptomycin. HeLa cells were plated in 12-well plate and Hepa 1–6 cells were plated in 24-well plates 24 h before transfection at 30% confluency in complete medium without antibiotics. HeLa cells were transfected with 10 nM of tiSMC, tiLDT, and their variants. Hepa 1–6 cells were transfected with 10 nM of tiApoB1B2B3 and its variants to analyze the target gene knockdown. T98G cells were plated in 24-well plates 24 h before transfection with 10 nM of variants of three different tiRNAs at 30% confluency to assess the activation of innate immune system. To demonstrate the role of RIG-I, T98G cells were plated in 24-well plates 24 h before transfection with 10 nM of siRIG-I (sc-61480; Santa Cruz Biotechnology), control siRNA (sc-37007; Santa Cruz Biotechnology), or siGFP as a control siRNA at 30% confluency. After 24 h recovery, the plated T98Gs were treated with polyinosinic–polycytidylic acid sodium salt (P0913; Sigma), 5′ ppp dsRNA (tlrl-3prna; InvivoGen), siRNA mixture, tiRNA, and tiRNA with three 3′ 2-nt overhangs. Lipofectamine RNAiMax was used for transfection of the described RNAs, following the manufacturer's protocol (Invitrogen). HeLa cells and Hepa 1–6 cells were harvested 24 h after transfection. T98G cells were harvested 12 h for tiSMC and tiApoB1B2B3, and 21 h for tiLDT after transfection for quantitative reverse-transcription PCR (RT-PCR).
Mice and peritoneal macrophage isolation
C57Bl/6 mice were bred and housed at a barrier and specific pathogen-free facility at the OliX Pharmaceuticals, Inc. (Suwon, Korea). All mice were purchased from Orient Bio (Seongnam-si, Korea). Six-week-old mice were used for all experiments. Brewer's thioglycollate broth (3%; Sigma) was prepared, and autoclaved. Then 5 mL of Brewer's thioglycollate broth was injected into the peritoneal cavity of each mouse. After 4 days, peritoneal cells were harvested from each mouse by peritoneal lavage with 1 × Dulbecco's phosphate-buffered saline (Invitrogen) and then centrifuged at 400 g for 20 min at room temperature (RT). After removing the supernatant, the cells were resuspended with 10 mL 1 × RBC lysis buffer (Invitrogen) and incubated for 5 min at RT. After centrifugation with the same protocol described above, the cells were resuspended with RPMI1640 (Gibco) and counted using Hemocytometer. Isolated mouse peritoneal macrophages were plated in 48 well with 5 × 105 cells. The experiment data of each mouse used the peritoneal macrophage harvested from each mouse, respectively. After incubating overnight, the plated cells were treated with polyinosinic–polycytidylic acid sodium salt (P0913; Sigma), 5′ ppp dsRNA (tlrl-3prna; InvivoGen), Lipopolysaccharide (tlrl-3pelps; InvivoGen), and variants of tiRNA. Lipofectamine RNAiMax was used for transfection of the described RNAs, following the manufacturer's protocol (Invitrogen). Mouse peritoneal macrophage cells were harvested 3.5 h after transfection of tiSMC and its variants, and 6 h after transfection of tiLDT and its variants for quantitative RT-PCR. All animal procedures were reviewed and approved by the OLIX Institutional Animal Care and Use Committee (OLIX-IACUC-AP-201805) and conducted in accordance with the Guide for the Care and Use of Laboratory Animals by the National Institutes of Health.
Quantitative RT-PCR
Total RNA was extracted using Tri-RNA Reagent (Favorgen), and then 0.5 mg of total RNA was used for complementary DNA (cDNA) synthesis using the high-capacity cDNA reverse transcription kit (Applied Biosystems) according to the manufacturer's protocol. cDNA was analyzed by quantitative RT-PCR using the StepOne Real-Time PCR System (Applied Biosystems). Primers for the target gene were mixed with SYBR Green PCR Master Mix (TaKaRa). The target genes and internal control mRNA levels were determined by using the relative standard curve quantitation method. The primer sequences for each gene are described in Supplementary Table S2.
Luciferase reporter assay
The 10 ng of ISRE or NF-κB luciferase reporter plasmid (pGL4.45[luc2P/ISRE/Hygro] and pGL4.32[luc2P/NF-κB-RE/Hygro]; Promega) and 1 ng of Renilla luciferase vector (pRL-SV40; Promega) were transfected using Lipofectamine 2000 (Invitrogen) with 2 × 104 T98G cells in 96-well plates, following the manufacturer's protocol. Then, 24 h after the reverse transfection of plasmids described above, the plated T98G cells were treated with polyinosinic–polycytidylic acid sodium salt (P0913; Sigma), 5′ ppp dsRNA (tlrl-3prna; InvivoGen), siRNA mixture, tiRNA, and tiRNA with three 3′ 2-nt overhangs. Lipofectamine RNAiMax was used for transfection of the described RNAs, following the manufacturer's protocol (Invitrogen). After 24 h, luciferase values were measured using the Dual luciferase reporter assay system (E1910; Promega), according to the manufacturer's instructions. The firefly luciferase signal was normalized to the Renilla luciferase signal, and the activity of each response element was expressed as fold induction over the unstimulated control sample.
Results and Discussion
Two-nucleotide overhang at the 3′ ends blocks the immune response of tiRNA by inhibiting the activation of RIG-I
We previously reported a tiRNA structure for RNAi therapeutics, which has three different siRNA units in one molecule (Supplementary Table S3) [6]. However, the immune stimulation induced by tiRNA has not been comprehensively studied. To understand the extent and mechanism of the innate immune activation by tiRNA structure, we synthesized three different tiRNAs, and their variants (Supplementary Fig. S1, S2 and Supplementary Table S3). The tiRNAs and their variants were transfected into T98G cell, and then the mRNA expression of IFIT1 gene was analyzed, which is a sensitive marker for the type I IFN production triggered by exogenous dsRNAs [11]. In the cytosol, it is generally accepted that long polyinosinic–polycytidylic acid (poly I:C) and 5′ ppp RNA are recognized by cytoplasmic innate immune sensors MDA5 and RIG-I, respectively [8,13]. We found that tiRNA with three blunt ends induced the innate immune response in T98G cell line, by increasing IFIT1 mRNA level. However, tiRNA with 3′ 2-nt overhangs induced a weaker immune response than tiRNA did. In particular, tiRNA's variants with 3′ 2-nt overhangs at all three ends rarely activated the innate immune system, which is similar to siRNA mixture (Fig. 1A–C). tiRNA variants with one 3′-overhang showed some level of immune activation, although lower than tiRNAs with no 3′-overhang (Fig. 1A–C). On the contrary, the presence of two 3′-overhangs to tiRNAs reduced immune activation more than tiRNAs with one 3′-overhang (Fig. 1A–C). We analyzed three additional cytokines such as CXL10, TNF-α, and IL-1β, which are related to innate immunity triggered by RNAs [15,16]. CXL10, a sensitive marker for activation of IRF3, showed mRNA expression profile similar to IFIT1, in all three tiRNAs tested (Fig. 1A–C) [17]. In contrast, mRNA levels of TNF-α and IL-1β, which are examples of proinflammatory cytokines, did not show significant changes in two out of three tiRNAs tested, tiApoB1B2B3 and tiLDT (Supplementary Fig. S3B, C). Only in case of tiSMC, mRNA expression of TNF-α and IL-1β increased about 10-fold compared with negative control (Supplementary Fig. S3A), suggesting that TNF-α and IL-1β induction by tiRNA is somewhat sequence dependent. Nevertheless, we note that the fold induction of TNF-α and IL-1β by this tiRNA is much less than that of IFIT1 and CXL10 (∼70-fold), suggesting that tiRNAs mainly stimulate innate immune response via IRF-3 pathway. Importantly, in all three tiRNAs we analyzed, the introduction of 3′ 2-nt overhangs did not cause any increase of the mRNA expression of all four cytokines tested.
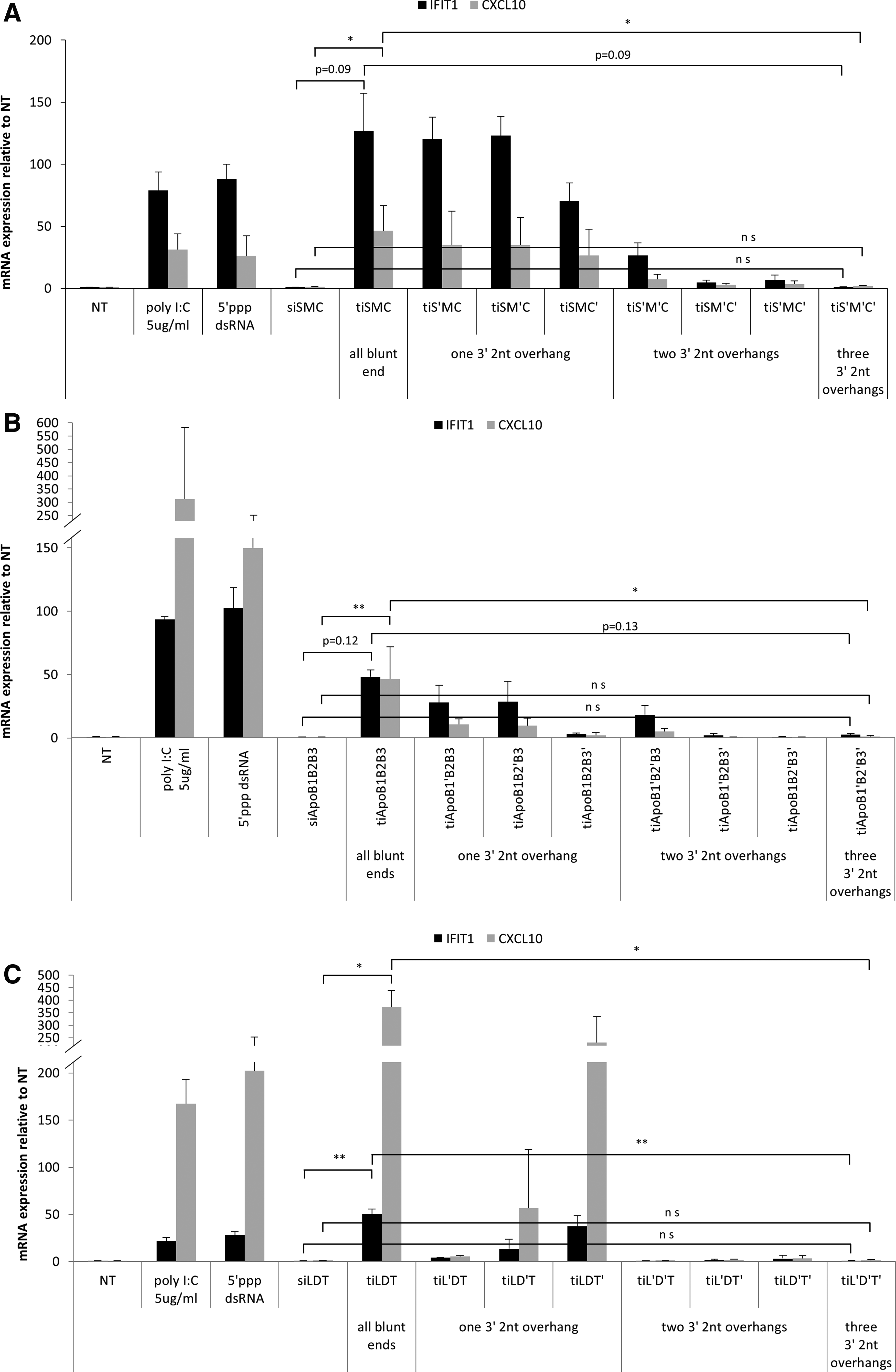
ISG activation triggered by tiRNA with blunt ends and 3′ 2-nt overhangs in T98G cell. Analysis of innate immune activation triggered by transfected three different tiRNAs and their variants in T98G cell line using quantitative reverse-transcription PCR (RT-PCR).
We hypothesized that tiRNA structure might be recognized by RIG-I because it has been reported that the 3′ 2-nt overhang structure prevented the recognition of synthetic dsRNAs by RIG-I [12,18]. To study the role of RIG-I in the recognition of tiRNA and successive immune activation, we assessed IFIT1 mRNA expression in RIG-I knockdown T98G cell line (Supplementary Fig. S2). IFIT1 mRNA expression by tiRNA was strongly suppressed in T98G cell line treated with RIG-I targeting siRNA (Fig. 2A and Supplementary Fig. S4). This confirms our hypothesis that immune activation by tiRNA structure is mediated by RIG-I, and the 3′ overhang structure appended to the tiRNA structure blocks the RIG-I-dependent innate immune response. We also conducted reporter luciferase assay to determine the activation of IRF and NF-κB because it is known that RIG-I signaling is propagated through IRF3 and NF-κB [9]. As expected, IRF3 was activated by all tiRNAs, but not by tiRNAs with three 3′ 2-nt overhangs (Fig. 2B–D). NF-κB was activated by tiSMC, consistent with the moderate activation of TNF-α and IL-1β by this RNA (Supplementary Fig. S3A). Again, the introduction of three 3′ 2-nt overhangs successfully blocked the activation of NF-κB to the level of siSMC (Fig. 2B–D). Taken together, these data suggest that tiRNA is recognized by RIG-I, and mainly activates innate immune response via IRF3 pathways. However, addition of 2-nt overhangs at the 3′ ends of tiRNA efficiently prevents the immune activation.

Analysis of the role of RIG-I in recognition of tiRNA, and reporter assay to verify the down-stream signaling pathway following RIG-I activation in T98G cells.
Analysis of the immunogenic determinant of tiRNA structure in mouse macrophage
RIG-I recognizes the specific RNA structure to discriminate self and nonself RNA [18]. Therefore, identification of the structural feature of tiRNA that is recognized by RIG-I is crucial, because it might unveil novel principle for discrimination between self and nonself RNAs. tiRNA was originally designed to have three fully hybridized 19 bp siRNAs, without any single-stranded region. However, we have previously reported that tiRNA was very sensitive to nuclease, rapidly generating short ∼19 bp fragments, whereas addition of 2′-OMe modifications of 6-nt at the central junction region made tiRNA almost completely resistant to the serum nuclease attack. This observation suggested that the junction of tiRNA structure might possess the single-stranded character, which is vulnerable to the attack of nucleases [6]. Therefore, to understand the core structure of tiRNA which is recognized by RIG-I, we dissected tiRNA structure into two separate parts; central junction and dsRNA arm regions. First, to elucidate the role of a single-stranded junction region of tiRNA in innate immune activation, we designed tiSMCss, which is designed to have a 6-nt long single-stranded bulge on each strand surrounding junction structure. tiRNAss-short is another structural variant of tiRNA; while tiSMCss-short has the central single-stranded junction like tiSMCss, it only has 13 bp-long dsRNA regions, which are shorter than those of tiSMC, which are 16 bp-long. Following the same strategy, we also designed tiLDTss-short (Supplementary Fig. S1, S2 and Supplementary Table S3). Thioglycolate-induced mouse peritoneal macrophage was used to assess the immune-stimulatory potential of tiSMC and tiLDT structural variants. Similar to the result obtained from T98G cells, while tiSMC and tiLDT induced the innate immune response in mouse macrophage, tiS'M'C' and tiL'D'T did not (Fig. 2E, F). tiSMCss showed a similar level of immune activation compared with tiSMC. In contrast, tiSMCss-short showed very weak immune activation compared with tiSMC and tiSMCss (Fig. 2E). tiLDTss-short also showed very weak immune activation which was similar to that of siLDT (Fig. 2F). These results suggest that the dsRNA regions in tiRNA are recognized by RIG-I, and the length of the dsRNA regions is critical for the proper recognition by RIG-I.
Analysis of target gene silencing by variants of tiRNAs
To explore the possibility that tiRNA with three 3′ 2-nt overhangs could be a novel RNAi platform, which is simultaneously targeting three different genes, we conducted quantitative RT-PCR to evaluate the target gene silencing efficiency. First, we tested RNA variants targeting mouse ApoB mRNA. Comparing with the mixture of three siRNAs targeting ApoB, tiApoB1B2B3, and tiApoB1B2B3 variants, including tiApoB1'B2'B3', resulted in the similar reduction of mouse ApoB mRNA (Fig. 3B). tiLDT and its variants, including tiL'D'T', also showed similar or better gene silencing efficiency against three different genes than siLDT (Fig. 3C). tiSMC variants, including tiS'M'C', also showed gene silencing as strong as tiSMC showed against Survivin and c-MET. An exception was the relatively weak gene silencing activity of tiSMC variants against β-catenin (Fig. 3A). In case of tiSMC, β-catenin gene silencing was not observed upon addition of 3′ 2-nt overhang (tiL'D'T') (Fig. 3C). Considering the fact that tiSMC showed a weaker β-catenin gene silencing efficiency than siSMC, we hypothesize that this observation might be due to relatively weaker affinity of β-catenin targeting RNA strand to Ago2, compared with RNA strands in the same tiSMC molecule targeting other genes. This affinity difference might become more significant by attaching 3′ overhang structures [19]. In summary, although careful sequence design should be needed to prevent the reduction of gene silencing activity, tiRNA and its variants with 3′ 2-nt overhang can execute efficient target gene silencing.

Analysis of target gene silencing of tiRNAs and their variants. Gene silencing effect of tiRNAs and its variants is measured by quantitative RT-PCR.
Various RNA structures have been developed to achieve multitargeting RNAi [4–6]. Our group previously developed a novel tiRNA structure, which consisted of three siRNA units, and triggered a sequence-specific gene silencing for three different target genes [6]. In this study, we also showed that the RNAi activity was maintained after adding 3′ overhang structures at all three ends of tiRNA. Although the molecular mechanism by which tiRNA induces RNAi has already been studied, the immunogenic property of tiRNA has not been thoroughly evaluated [6,20]. In this study, the immune stimulating potential of tiRNA was investigated in T98G cell line, and mouse macrophage cells. tiRNA structure with all blunt ends showed immune activation in both cell types tested. Reporter-based assay revealed that tiRNAs mainly activated IRF to initiate the innate immune response; however, one tiRNA molecule, tiSMC, also activated NF-κB, suggesting that the NF-κB activation by tiRNA is sequence-dependent (Supplementary Fig. S3A). We would like to note that, in addition to the activation of NF-κB pathway, the magnitude of IRF-dependent immune response is different among tiRNAs we tested, suggesting that tiRNA-mediated immune activation has sequence-dependent feature (Fig. 2A). Nevertheless, the important finding is that all immune response triggered by tiRNAs can be abrogated by adding 3′ overhangs. This observation strongly pointed out RIG-I as a main culprit of immune sensor, which was confirmed by experiments using RIG-I-specific siRNA.
It was previously reported that 14 bp-long blunt-ended dsRNA can bind to the CTD of RIG-I in vitro [21]. In our result, tiRNA harboring 16 bp-long dsRNA region (tiRNAss) induced the innate immune response, whereas tiRNA with 13 bp-long dsRNA (tiSMCss-short) failed to do so. This indicates that a certain minimum length of dsRNA is required for activation of RIG-I by tiRNA, and that the minimum length of dsRNA would be between 14 and 15 bp, which is consistent with the previous in vitro finding [21]. We tested gene silencing activity of tiSMCss-short, which only has 13-nt RNA region complementary to the target mRNA. This construct did not show significant gene silencing (Supplementary Fig. S5). Similarly, tiSMCss, which has 16-nt sequence complementary to the target mRNA, also showed reduced gene silencing activity. In contrast, in our previous report tiApoB1B2B3ss-short and tiLDTss-short structures with 19-nt sequence complementary to the target mRNA, showed strong target gene silencing [22,23]. Combined together, we believe that tiRNAss-short structure can be potentially developed into another multiple target gene silencing RNA structure without immunostimulation, as long as 19-nt long RNA sequence complementary to the target mRNA is present. In conclusion, our study extends the diversity of RIG-I ligand structures by adding tiRNA to the list. Also, considering the tiRNA with 3′ overhangs structure maintains RNAi, controlling the immune stimulation by tiRNA with the addition of 3′ overhang structure is important in future development of multitargeting RNAi therapeutics based on tiRNA structural platform.
Footnotes
Acknowledgment
This work was supported by a Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2017R1D1A1B03036001).
Author Disclosure Statement
D.-k.L., and J.Y.C. are employees and shareholders of OliX Pharmaceuticals, Inc. The other authors have no competing financial interests to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.