
Abstract
The increasing incidence of various cancers indicates the urgent need for finding accurate early diagnostic markers and more effective treatments for these malignancies. MicroRNAs (miRNAs) are small noncoding RNAs with great potentials to enter into cancer clinics as both diagnostic markers and therapeutic targets. miR-21 is elevated in many cancers, and promotes cell proliferation, metastasis, and drug resistance. In recent years, many studies have shown that targeting miR-21 combined with conventional chemotherapeutic agents could enhance their therapeutic efficacy, and overcome drug resistance and cancer recurrence both in vitro and in animal models. In this review, we first summarize the effects and importance of miR-21 in various cancers, and explore its function in drug resistance of cancer cells. Next, the challenges and prospects for clinical translation of anti-miR-21, as a therapeutic agent, will be discussed in combination cancer therapy.
Introduction
Despite significant breakthroughs in cancer screening and treatment strategies, various malignancies are still accounting for the second leading cause of death worldwide [1]. One major obstacle in cancer treatment is drug resistance, which is associated with high rates of tumor recurrence and metastasis. Two types of drug resistance in cancer cells include primary and acquired resistance, and major causes of this phenomenon are tumor heterogeneity and cancer stem cells (CSCs). Numerous studies have shown several mechanisms for drug resistance such as prevention of drug uptake, drug inactivation and efflux by ATP-dependent pumps, resistance to apoptosis, increased repair of DNA damage/tolerance, and epigenetic alterations.
MicroRNAs (miRNAs) are a group of evolutionary conserved small noncoding RNAs, which are involved in some vital biological processes; and thus, changes in their expression may disrupt normal cell functions leading to tumor formation [2]. Studies have indicated that miRNAs are major regulators of drug resistance by targeting genes related to promoting and development of this natural phenomenon [3].
miR-21 is a notable miRNA with a significant overexpression in most cancer types. Interestingly, aberrant expression of miR-21 is involved in tumor initiation, development and metastasis, decreased survival and resistance to apoptosis and therapeutic agents. This miRNA is also secreted by multiple components of tumor microenvironment, including cancer-associated fibroblasts and myeloid-derived suppressor cells, which stimulate tumor cell migration and invasion [4,5]. Furthermore, there is a significant positive correlation between miR-21 elevated levels and TNM stages of different tumors with poor prognosis. An increased level of this oncomiR has also been reported as a diagnostic as well as prognostic marker in various cancer types [6,7]. Exosomal miR-21, which is secreted from tumors into the blood, can also be used as a potential noninvasive diagnostic marker [8,9]. An increasing number of studies indicate that CSCs are responsible for therapeutic resistance. Consistently, miR-21 plays important roles in stemness regulation and cancer drug resistance; therefore, it seems that targeting this oncomiR sensitizes tumors to therapeutic agents [10].
Chemotherapeutic drugs can also damage most normal dividing cells in the body, so they result in adverse effects on the blood producing cells in the bone marrow, increased susceptibility to various diseases, gastrointestinal toxicity, hepatotoxicity, and reproductive health concerns. miR-21 antisense therapy together with chemotherapeutic agents can exhibit markedly synergistic anticancer effects, and therefore using a lower therapeutic dosage of these agents can lead to a significant inhibition of tumor growth with less side effects [11,12].
Furthermore, there is evidence that both miR-21-5p and miR-21-3p isoforms contribute to cancer drug resistance; therefore, depending on cancer type, targeting either isoform would elicit better responses. In human, 30 and 10 isoforms have been reported for miR-21–5p and miR-21–3p, respectively. In a study, in 2020 it was shown that monoadenylated isoform of miR-21-5p was enriched in cancer samples, which might reflect its critical roles during tumor development [13]. In another study conducted by Jiao et al., they revealed that different miR-21-3p isoforms target different genes and have variable effects on the ability of colon cancer cell migration [14]. Interestingly, among major isoforms of miR-21–3p, miR-21-3p 0 | 2 had favorable adverse effects through targeting the messenger RNA (mRNA) of some oncogenes in colon cancer [14]. Therefore, it seems that upregulation of miR-21-3p 0 | 2 along with downregulation of monoadenylated isoform of miR-21-5p may synergically sensitize cancer cells to therapeutic agents to have promising results in combination cancer therapy.
Here, we review the available data on regulation and function of miR-21 in cancer cells, particularly in CSCs. Then, the importance of this carcinogenic miR in different mechanisms of cancer drug resistance will be summarized.
Regulation of miR-21 Expression in Cancer Cells
miR-21 is located in a fragile site and is frequently amplified at DNA level in different cancers. Expression of this oncomiRNA can be activated by binding different transcription factors (TFs) to several conserved elements on its promoter region (Fig. 1). Activator protein 1 (AP-1) is a heterodimeric TF, which is closely related to various cellular events such as differentiation, proliferation, and apoptosis. Chromatin immunoprecipitation (ChIP) and DNA electrophoretic mobility shift assays have revealed that activated AP-1 can bind to miR-21 core promoter and triggers upregulation of this miRNA [15]. Then, miR-21 exerts its oncogenic effects by suppressing programmed cell death 4 (PDCD4), which leads to enhanced AP-1 expression by a feedback loop between miR-21/PDCD4/AP-1 [16].
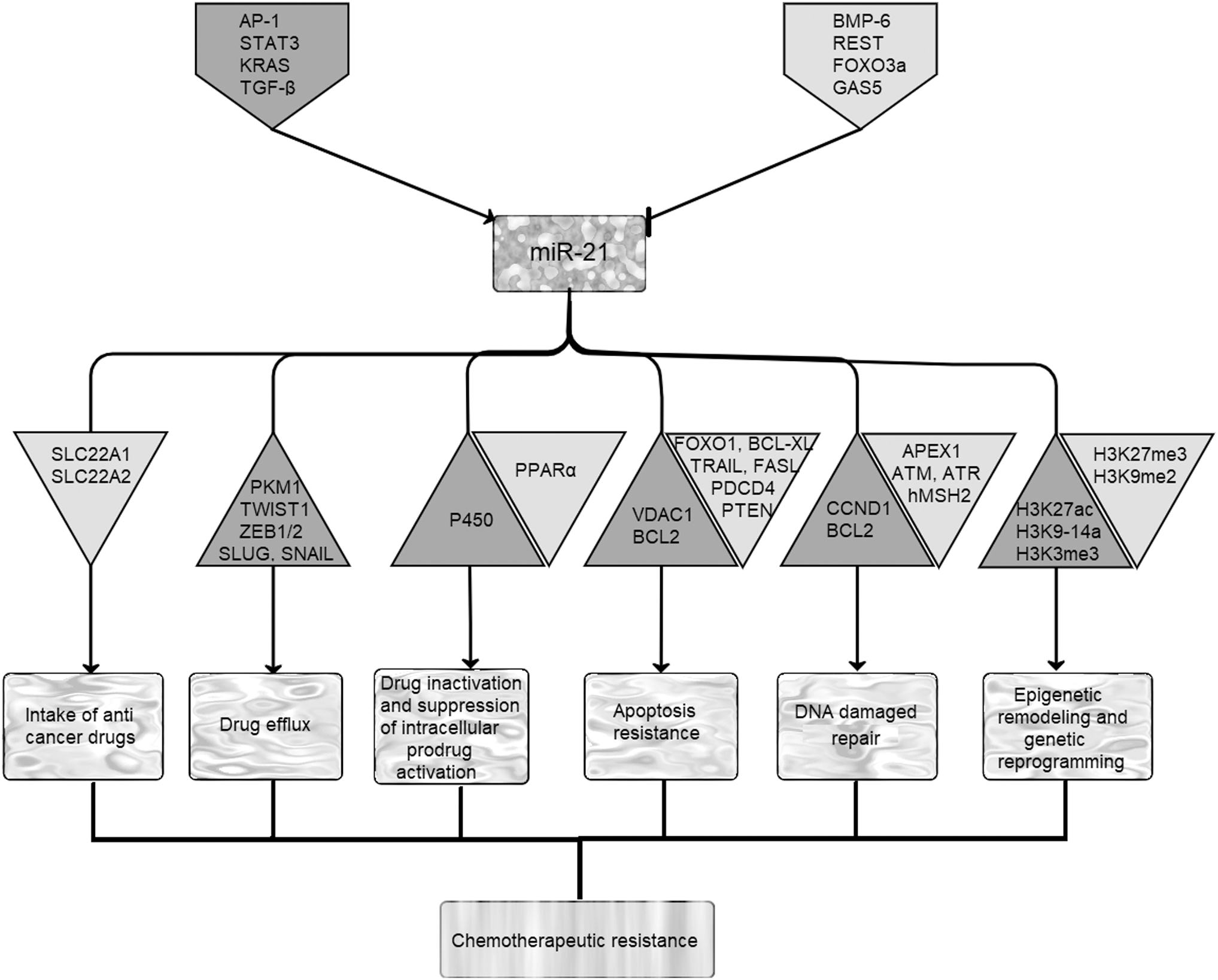
Regulation of miR-21 expression in tumor cells and its subsequent effects on cancer multidrug resistance. Expression of miR-21 can be activated or repressed by binding different proteins/noncoding RNAs, which are represented in gray and white pentagons, respectively. Gray and white triangles, respectively, represent upregulated and downregulated proteins after overexpression of miR-21. AP-1, activator protein 1; APEX1, apurinic/apyrimidinic endodeoxyribonuclease 1; ATM, ataxia telangiectasia mutated; ATR, ATM- and Rad3-related; BCL2, B-cell lymphoma 2; BCL-XL, B-cell lymphoma-extra large; BMP-6, bone morphogenetic protein-6; CCND1, cyclin D1; FASL, Fas ligand; FOXO1, forkhead box protein O1; FOXO3a, forkhead box O3; GAS5, growth arrest-specific transcript 5; hMSH2, human DNA MutS homolog 2; KRAS, Kirsten rat sarcoma; P450, cytochromes P450; PDCD4, programmed cell death 4; PKM1, pyruvate kinase muscle 1; PPARα, peroxisome proliferator-activated receptor α; PTEN, phosphatase and tensin homolog; REST, RE1-silencing transcription factor; SLC22A1, solute carrier family 22 member 1; STAT3, signal transducer and activator of transcription 3; TGF-β, transforming growth factor beta; TRAIL, TNF-related apoptosis-inducing ligand; TWIST1, twist family BHLH transcription factor 1; VDAC1, voltage-dependent anion channel 1; ZEB1/2, zinc finger E-box-binding homeobox 1/2.
Interleukin-6 (IL-6), an important tumor promoting factor, is mainly secreted by the malignant cells or other cells in the tumor microenvironment, and stimulates proliferation and development of cancer cells [17]. After an increase in IL-6, signal transducer and activator of transcription 3 (STAT3), a master TF, which plays an important role in tumorigenesis, translocates to the nucleus and mediates induction of miR-21 through binding to an upstream enhancer region. Afterward, miR-21 enhances NF-κB activity, which leads to upregulation of NF-κB and IL-6. In addition, miR-21 expression is induced by NF-κB through a positive feedback loop in response to an inflammatory stimulus or DNA damage [18].
In Her2/neu overexpressing breast cancer, lung cancer, and thyroid carcinomas, activation of mitogen-activated protein kinase (MAPK) signaling, targets miR-21 overexpression by oncogenic Kirsten rat sarcoma (KRAS), and induces cell migration, invasion, and drug resistance through tumor suppressors' inhibition [19–21]. As reported earlier, transforming growth factor beta (TGF-β)-activated SMAD-3/4 promotes epithelial-to-mesenchymal transition (EMT) 1through increasing miR-21 expression, and miR-21 can also reciprocally amplify the TGF-β signaling pathway through forming a feedback circuit between them [22,23]. However, bone morphogenetic protein-6 (BMP-6), RE1-silencing transcription factor (REST), and forkhead box O3 (FOXO3a), which are downregulated in cancer cells, act as negative regulators of miR-21 expression [24,25]. Studies have also demonstrated that long noncoding RNA (lncRNA) growth arrest-specific transcript 5 (GAS5), a pivotal tumor suppressor that serves an essential role in normal growth arrest, could directly bind to miR-21 and negatively regulate this oncomiR expression, which modulates growth and apoptosis in cancer cells [26–28].
Downstream Pathways of miR-21 in Cancer
Insights into the roles of this small RNA revealed that miR-21 negatively regulates a network of key tumor-suppressive pathways. Phosphatase and tensin homolog (PTEN)/PI3K/AKT is an important signaling pathway, which is activated in several cancers. It has been shown that upregulation of miR-21 can lead to abnormality in survival, proliferation, angiogenesis, invasion, migration, and resistance of tumor cells to chemotherapeutic agents in most cancer types through targeting PTEN, a major regulator of PTEN/PI3K/AKT pathway [29]. However, in some cancers with mutated or loss of PTEN, another tumor suppressor protein, like PDCD4, was inhibited by miR-21 at post-transcriptional level. Downregulation of PDCD4 aberrantly hyperactivates MAP4K1/JNK/AP-1 and JAK/STAT3 pathways, which increase cancer cell proliferation and migration.
On the other side, miR-21 induces angiogenesis and suppresses apoptosis through activating MAPK/ERK pathway. A recent research indicated that miR-21-mediated hepatoblastoma progression and resistance to apoptosis happened through ASPP2/P38 signaling pathway. Liu et al. also revealed that miR-21 suppresses the expressions of ASPP2, a key regulator of apoptosis, while it upregulates P38, a member of the MAPK family [30]. Besides, miR-21 activates oncogenic pathway RAS/RAF-1/MEK1/ERK through regulation of RAS P21 GTPase activating protein 1 (RASA1) and promotes the malignant phenotypes in colon cancer [31]. It has been indicated that miR-21 may contribute to deregulation of extracellular matrix in tumor microenvironment, which was mediated by matrix metalloproteinases (MMPs). Downregulation of TIMP3, a tissue inhibitor of MMPs and a direct target of miR-21, is mediated through TGF-β pathway, and promotes tumor cell infiltration and angiogenesis [32,33]. In addition, Fas ligand (FASL), a type-II transmembrane protein as a member of the TNF family involved in apoptosis induction, is another important target of miR-21; its inhibition contributes to increased cell survival and apoptosis resistance through suppressing caspase 8 activation [34]. Notably, miR-21 targeted TNF signaling pathway, through inhibition of TNF receptor 1 (TNFR1) while activating TNFR2, which result in suppression of apoptosis in cervical cancer [35].
miR-21 and CSCs
CSCs constitute a small subpopulation of cells within tumors with self-renewal ability that attribute to tumor multidrug resistance. They are involved in cancer initiation and progression, and have the potential to differentiate into various cell lineages. Aldehyde dehydrogenase-1, CD24, CD44, and CD133 are among the most common signatures for solid tumor stem cell markers.
Publications have addressed the function of miR-21 in CSCs by downregulation of TGF-βR2 [36], which augments Wnt/β-catenin signaling pathway and induces stemness in colon and, head and neck CSCs [36,37]. In addition, it has been revealed that miR-21 contributes to CSC regulation with miR-145 in a negative feedback loop mediated by KRAS. In a study, Yu et al. reported that upregulation of miR-21 along with downregulation of miR-145 induces drug resistance and suppresses differentiation in CSCs. Overexpression of miR-21 enhances KRAS, which in turn causes a decrease in miR-145 levels and finally results in upregulation of pluripotency factors SOX2, NANOG, and OCT4 in colon CSCs. Besides, promotion of Wnt/β-catenin signaling pathway by miR-21 supports these CSCs growth and dedifferentiation simultaneously [38]. In breast CSCs, miR-21 contributes to epithelial–mesenchymal transition, metastasis, and angiogenesis through increased levels of hypoxia-inducible factor-1α (HIF-1α). In these CSCs, miR-21 activates AKT/ERK1/2 pathways through suppression of PTEN [39]. An increasing number of studies have shown that the combination of miR-21, miR-9, miR-128, miR-218, miR-146a, and miR-326 promoted cell proliferation and self-renewal properties in glioblastoma stem cells (GBMSCs) [40]. On the contrary, Aldaz et al. reported that miR-21 and its network are key regulators in differentiation of CSCs in GBM, and suppression of this miRNA causes GBM recurrence through decreased expression of a stem cell marker, Nestin [41].
CD133, a pentaspan membrane glycoprotein, is commonly upregulated in CSCs and has been extensively used as a marker for isolation of stem cells from cancerous tissues. Overexpression of miR-21 enhances the expression of CD133 marker, which results in increased sphere formation capacity of ovarian CSCs [42]. Overexpression of CD44, another transmembrane glycoprotein marker in CSCs, is also related to miR-21 expression in colon CSCs, and led to sphere formation and proliferation of these cells [36]. In addition, interaction of CD44 with c-Jun results in transcriptional activation of miR-21 and consequently increased chemotherapeutic resistance in cancer cells [43]. Thus, these data indicate that miR-21 plays an essential role in cancer by regulating CSC properties [42].
Another target of miR-21 in GBM stem cells is FASL. Actually, miR-21 contributes to GBMSCs resistance to apoptosis through downregulation of FASL in TNFα pathway [44]. Recently, Zhu et al. have revealed that miR-21-3p, the passenger strand of miR-21, can suppress TNF-related apoptosis-inducing ligand (TRAIL). TRAIL, a death ligand, induces apoptosis through interaction with death receptor 4 (DR4) and DR5. In liver CSCs, miR-21-3p promotes PI3K/AKT/BAD pathway by inhibiting PTEN and suppression of TRAIL expression, which result in cell survival [45]. In this regard, in another study it has been reported that miR-21-3p may promote proliferation of esophageal CSCs and exerts its antiapoptotic effects through regulating TNF receptor-associated factor 4 (TRAF4). In fact, miR-21-3p inhibits TRAF4 expression, which leads to the stemness maintenance in these CSCs [46].
Role of miR-21 in Drug Resistance Mechanisms in Cancer Cells
Intake of anticancer drugs
The intake of anticancer drugs occurs through solute carrier family (SLC) proteins, which are localized on the cell membrane. Downregulation of SLCs or reducing the tendency of these proteins to drugs intake leads to reducing the intracellular concentration and efficacy of therapeutic agents [86]. OCT constitutes a large family of transporters including OCT1, OCT2, and OCT3 [also known as solute carrier family 22 member 1 (SLC22A1)–SLC22A3], which enable the drugs to enter the cells by passive diffusion [87].
It has been revealed that downregulation of miR-21 in doxorubicin-resistant human hepatocellular carcinoma cells [HepG2/ADR (adriamycin)] resulted in an increased cellular uptake and accumulation of this anticancer agent, which promoted apoptosis in tumor cells [88]. Gaudelot et al. have reported a negative correlation between miR-21 overexpression and upregulation of SLC22A1/OCT1 and SLC22A2/OCT2, in renal carcinoma (Fig. 1 and Table 1) [84]. They showed that anti-miR-21 reverses paclitaxel (PTX), 5-fluorouracil (5-FU), oxaliplatin, and dovitinib resistance in renal carcinoma. Therefore, conjugation of anti-miR-21 and anticancer drugs significantly induced apoptosis in these cancerous cells [84].
Importance and Involvement of miR-21 in Response to Chemotherapy Agents
3-MA, 3-methyladenine; 5-FU, 5-fluorouracil; ADR, adriamycin; Ara-C, arabinosylcytosine; BCL2, B-cell lymphoma 2; CCND1, cyclin D1; EGFR, epidermal growth factor receptor; FASL, Fas ligand; HIF-1α, hypoxia-inducible factor-1α; hMSH2, human DNA MutS homolog 2; MDR, multidrug resistance; ND, not determined; OCT, organic cation transporter; PDCD4, programmed cell death 4; P-gp, P-glycoprotein; PTEN, phosphatase and tensin homolog; SLC22A1, solute carrier family 22 member 1; TRAIL, TNF-related apoptosis-inducing ligand.
Drug efflux
Drug efflux is a key mechanism in the extrusion of cytotoxic agents and other substrates from cells, thereby reducing the intracellular accumulation and associated toxicity of these compounds. Active drug efflux transporters of the ATP-binding cassette (ABC) superfamily are found in all prokaryotic and eukaryotic species, and mediate the transbilayer movement of clinically relevant drugs in intact cells. Overexpression of the ABC transporters mainly including P-glycoprotein (P-gp), multidrug resistance proteins (MRPs) 1–5, and breast cancer resistance protein (BCRP) contributes to multidrug resistance (MDR) in cancer cells [89].
miR-21 can increase drug resistance in cancer cells through upregulation of ABC pumps. It has been revealed that the expression of MDR genes (ABCC2–6) is increased in cancer cells overexpressing miR-21. Therefore, suppressing miR-21 sensitizes tumor cells to chemotherapeutic agents [84]. In a study, Xie et al. explored the mechanism by which overexpression of P-gp is induced in ovarian cancer. Their results indicated that aberrant expression of miR-21 and HIF-1α contributed to upregulation of MDR1/P-gp expression. Therefore, inhibition of miR-21 results in downregulation of HIF-1α, which sensitized ovarian cancer cells to PTX, a common chemotherapy drug used for treatment of several types of cancers [79]. Information about molecular mechanism of P-gp-mediated drug resistance in gastric cancer also revealed that miR-21 can upregulate HIF-1α. In turn, overexpression of this protein results in enhancing pyruvate kinase muscle 1 (PKM1) expression, which may have a critical role in drug resistance acquisition [90]. Previous studies have found that WNT/β-catenin signaling pathway plays an important role in P-gp-mediated drug resistance. As miR-21 is involved in cancer initiation through WNT/β-catenin signaling pathway, it might be associated with upregulation of multidrug-resistant proteins through targeting this pathway.
It has been shown that EMT is a regulator of ABCB1 transporters in tumor cells [91]. During epithelial–mesenchymal transition, some of EMT-related TFs such as twist family BHLH transcription factor 1 (TWIST1), zinc finger E-box-binding homeobox 1/2 (ZEB1/2), SLUG, and SNAIL can also simultaneously regulate expression of ABC transporters and induce drug resistance in cancer cells (Fig. 1) [91]. As most of these TFs are miR-21 target genes, this miR has an important role in EMT [92]. Thus, in addition to having a direct role in overexpression of ABC transporters, miR-21 can indirectly target drug resistance through increasing EMT.
Drug inactivation and suppression of intracellular prodrug activation
Two main mechanisms that reduce the effectiveness of chemotherapeutic agents in tumor cells are drug inactivation and suppression of intracellular prodrug activation [93]. A prodrug is a compound that can acquire clinical efficacy by intracellular metabolic biotransformation, while drug inactivation includes detoxification enzymatic reactions, which chemically alter drugs or destroy them, resulting in resistance of tumor cells to active pharmacological agents [94]. A number of studies have shown high degree of correlation between miRNAs expression and prodrug activation/drug inactivation enzymes [95]. miR-21 can decrease expression of enzymes involved in activation of 5-FU, thymidine phosphorylase (TYMP), and dihydropyrimidine dehydrogenase (DPD), through targeting human nuts homolog 2 leading to resistance of colon cancer cells to chemotherapeutic treatments [43,96].
Cytochrome P450 (P450)-related enzymes are a superfamily of heme proteins, which are responsible for detoxification of drugs through oxidation and some reduction reactions. These heme proteins conjugate with enzymes involved in phase 2 detoxification reactions [eg, glutathione (GSH), sulfate, and glycine transferase] to inactivate and alter the molecular characteristics of activated drugs [93]. Expression of these enzymes increases in cancer cells and results in enhanced chemoresistant relapses. Positive correlation between miR-21 and members of cytochrome P450 (CYP1A1 and CYP2A6) has been reported in hepatic pathophysiological conditions showing the regulatory effects of this small RNA on expression of these enzymes (Fig. 1) [95,97,98].
In addition, it has been revealed that miR-21 suppresses the expression of peroxisome proliferator-activated receptor α (PPARα), which regulates the expression of some detoxification enzymes. Thus, upregulation of miR-21 contributes to resistance of some colon cancer cells to hydroxycamptothecin (HCPT) treatment through reduction of PPARα (Fig. 1) [98].
Apoptosis resistance
Apoptosis resistance is an emerging concept in cancer therapy that contributes to carcinogenesis and tumor progression. Apoptosis can be regulated by two broad pathways: the extrinsic pathway (death receptor pathway) and the intrinsic pathway (the mitochondrial pathway); both ultimately lead to activation of downstream effector caspases [99].
Evidence has shown that miR-21 is involved in regulation of extrinsic apoptosis pathways. Elevated levels of miR-21 and miR-30b/c hinder TRAIL-dependent apoptosis by inhibiting the expression of TAp63 and caspase 3 (Fig. 1) [100]. Furthermore, miR-21 in combination with miR-30c and miR-100 can induce TRAIL resistance-mediated apoptosis in nonsmall-cell lung cancer (NSCLC) through suppression of caspase 8, caspase 3, TRAF7, and FOXO3a [53]. Inhibition of miR-21 suppresses NF-κB activation and sensitizes these cancer cells to TRAIL-induced apoptosis (Table 1). Yang et al. reported that overexpression of miR-21 enhances resistance of nasopharyngeal carcinoma (NPC) cells to cisplatin through inhibition of proapoptotic factors PDCD4 and FASL (Table 1) [60].
Mitochondrion represents a critical organelle in activating intrinsic apoptosis pathway. Any changes in the channels/pores present in outer and inner membranes of this organelle are associated with drug resistance of cancer cells [101]. Voltage-dependent anion channel 1 (VDAC1), an ion channel located in the outer mitochondrial membrane, is mostly upregulated in different cancers [102]. This channel can also interact with hexokinase (HK) and translocate ATP to this enzyme to increase glycolysis in cancerous cells. Papagiannakopoulos et al. reported that repression of miR-21 using LNA-miR-21 can reduce the expression of VDAC1, and lead to apoptosis induction in glioblastoma (Fig. 1) [103]. In addition, upregulation of antiapoptotic protein B-cell lymphoma 2 (BCL2), a direct target of miR-21, enhances resistance of cancer cells to chemotherapeutic drugs. For instance, downregulation of miR-21 increased sensitivity of breast cancer cells to topotecan, a chemotherapeutic agent that inhibits topoisomerase 1, through downregulation of BCL2 (Table 1) [54]. In addition, dysregulation of BCL2 and PTEN induced by miR-21 promotes plutonium resistance in NSCLC (Fig. 1 and Table 1) [73].
Forkhead box protein O1 (FOXO1), a direct target of miR-21, is downregulated in tumor cells. Studies have shown that miR-21 can target FOXO1, which regulates the expression of proapoptotic BIM and suppresses PI3K/AKT/mTOR pathway through inhibition of mTOR in glioblastoma and diffuse large B-cell lymphoma (DLBCL) [104,105]. Therefore, suppression of miR-21 results in upregulation of FOXO1-BIM pathway, which sensitizes cancer cells to chemotherapeutic agents and apoptosis (Fig. 1) [104,105]. It has been revealed that upregulation of miR-21 through JNK-1/c-Jun leads to cisplatin resistance, increased proliferation, and invasion in ovarian cancer. Thus, suppression of miR-21 expression in these cancer cells can overcome cisplatin resistance, inhibit proliferation, and induce apoptosis through upregulation of its tumor suppressor targets like PDCD4 (Table 1) [77]. PDCD4 can negatively modulate antiapoptotic proteins through direct interaction with B-cell lymphoma-extra large (BCL-XL) and XIA, or inhibit their related mRNAs translation through binding to the internal ribosome entry site (IRES) element in their 5′-untranslated regions (UTRs) [88,106].
Repression of miR-21 could also reverse the resistance of NSCLC to gefitinib, which targets epidermal growth factor receptor (EGFR) and leads to increased levels of PTEN after treatment (Table 1) [85]. It has been shown that PTEN can mediate intrinsic apoptosis pathway through suppression of focal adhesion kinase (FAK), which plays an antiapoptotic role through stimulation of P53 proteasomal degradation and/or inhibition of BAX expression [107,108]. Furthermore, PTEN prevents MADD phosphorylation and activation through PI3/AKT, and makes cancer cells more susceptible to extrinsic apoptosis pathway [109]. Other studies have shown that miR-21 increases sensitivity of NSCLC to EGFR-tyrosine kinase inhibitor (TKI) through inhibition of PTEN and PDCD4 and activation of PI3K/AKT signaling. Likewise, suppression of miR-21 reverses resistance of NSCLC to cisplatin through upregulation of PTEN (Table 1) [72,110]. Curcumin, the main bioactive substance in turmeric with various anticancer effects, can repress the miR-21 expression by binding to its promoter. Ali et al. have reported that pancreatic cancer cells were more sensitive to gemcitabine, an anticancer chemotherapeutic drug used in various carcinomas, after inhibition of miR-21 by curcumin (Table 1) [83]. Furthermore, difluorinated curcumin (CDF), a fluorinated curcumin analog, inhibits the growth of 5-FU and oxaliplatin-resistant colon cancer cells through modulation of miR-21, which stimulates the well-known drug resistance PI3/AKT pathway [111,112]. This evidence shows that upregulation of miR-21 cripples cancer treatments through apoptosis and cell survival pathways.
DNA damage repair
DNA damage repair and/or tolerance are the well-known mechanisms contributing to cancer cells resistance to chemotherapy. Some anticancer agents like cisplatin induce apoptosis in cancer cells by either directly or indirectly damaging the DNA [113]. However, DNA repair pathways can reverse the anticancer effects of chemotherapeutic drugs. There are five different DNA repair pathways from which nucleotide DNA mismatch repair (MMR), nucleotide excision repair, and homologous recombination are mostly linked to drug resistance of cancer cells [114]. Therefore, applying DNA damaging anticancer agents in combination with inhibition of repair systems can improve the efficacy of therapeutic agents and increase sensitivity of cancer cells to these drugs.
Different studies have revealed that miRNAs influence DNA repair processes and cellular response to DNA damaging agents. In this regard, Valeri et al. have shown that miR-21 increases resistance of colon cancer cells to 5-FU through downregulating a MMR gene, human DNA MutS homolog 2 (hMSH2), which increases the sensitivity of cancer cells to genotoxic agents (Table 1) [58]. Anti-miR-21 treatment of cancer cells dysregulates the PI3K/AKT/mTOR pathway, which has implications in DNA repair response after therapeutic treatments [115,116]. Poly (ADP-ribose) polymerase (PARP), a nuclear enzyme, is involved in intracellular DNA damage response and plays an important role in cell survival. It has been demonstrated that inhibition of MSH2 by miR-21 confers TGF-β-induced resistance to PARP inhibitor (PARPi), which selectively targets tumor cells with a DNA repair deficiency, in breast cancer cells [117]. Furthermore, miR-21 affects cell-cycle progression after cancer treatment.
Manoel-Caetano et al. predicted that expression of genes involved in recognition of DNA damage [ataxia telangiectasia mutated (ATM), ATM- and Rad3-related (ATR), and apurinic/apyrimidinic endodeoxyribonuclease 1 (APEX1)] is suppressed by miR-21-5p in gastric cancer (Fig. 1) [118]. Therefore, it seems that miR-21 can contribute to survival of gastric cancer cells through downregulating genes involved in DNA repair system. During DNA damage, ATM and ATR kinases activate Checkpoint kinase CHK1/CHK2, which leads to cell-cycle arrest by activating CDC25A [119]. CDC25A, a protein phosphatase, regulates cyclin E- and A-associated cdk2 [120], and is an important regulator of cell cycle during DNA damage treatment. CDC25A is subjected to modulation by miR-21 in human colon and glioblastoma cancer cells; therefore repression of miR-21 slows down the G2/M progression through important G2-check point, and eventually promotes apoptosis in cancer cells [115]. Thus, downregulating miR-21 in combination with other DNA damaging therapeutic agents can effectively induce apoptosis in cancer cells [121,122]. Furthermore, another study has reported that upregulation of cyclin D1 (CCND1) and BCL2 by miR-21 caused G1/S transition and cell-cycle progression in cancer cells (Fig. 1). It has been demonstrated that targeting miR-21 arrests cell cycle and increases apoptosis in these cells [63].
Epigenetic remodeling and genetic reprogramming
Genetic instability and epigenetic remodeling are mostly responsible for changes in expression profile of miRNAs and drug resistance in cancer cells. Epigenetic processes are reversible, so they are important in cancer treatment. Epigenetic remodeling mechanisms mostly include DNA methylation and histone modifications through acetylation or methylation [123]. DNA methylation occurs by the covalent transfer of a methyl group to cytosines at CG-dinucleotides in CpG motifs. Conversely, histone modification is a post-translational modification including acetylation, methylation, ubiquitination, citrullination, and phosphorylation, which changes physical properties of chromatin [124].
Expression of miR-21 is controlled at both genetic and epigenetic levels. Therefore, these two post-translational modifications are also associated with miR-21 dysregulation in some cancers. The CpG island sequences are located upstream of miR-21 and are subjected to hypomethylation in ovarian cancer, which is responsible for upregulation of this noncoding RNA [125]. In addition, Ferraro et al. performed a ChIP analysis, and revealed that H3K9–14ac, H3K3me3, and H3K27ac modifications that result in activating transcription were higher in miR-21 promoter regions, whereas H3K27me3 and H3K9me2 modifications, associated with inactive transcription state, were absent in these regions in EMT colon cancer cells (Fig. 1) [126]. It seems that epigenetic upregulation of miR-21 plays important roles in colon cancer carcinogenesis.
In another study, it has been shown that in 3,6-dihydroxyflavone (3,6-DHF)-treated breast cancer cells, H3K9–14 modifications, associated with inactive transcription, were significantly reduced compared with untreated control cells. In other words, treatment with 3,6-DHF alters the histone modification in breast cancer cells and effectively suppresses the growth of these cells through downregulation of miR-21 [127]. All of these studies suggest that therapeutic agents can target the epigenetic remodeling of miR-21, and by down regulating the expression of this miR, drug-resistant cancer cells can be sensitized and the therapeutic efficacy of chemotherapy can be improved. Furthermore, according to the study conducted by Chen et al., miR-21 potentially promotes reprogramming of cancer cells and inhibits genes involved in proteasome pathway, which results in drug resistance. They have reported that overexpression of miR-21 in colon cancer cells mimics genetic reprogramming related to topoisomerase inhibitors, and confers resistance to SN-38, doxorubicin, and etoposide/VP-16 treatments (Table 1) [128].
miR-21 Involvement in Other Combination Treatments for Cancer
Targeting miR-21 has also improved other treatments for cancer with higher efficacy. Radiation therapy serves as a type of cancer treatment that provides a significant survival advantage for patients. It has been revealed that miR-21 confers radiation resistance to glioma and breast cancer cells through G2/M cell-cycle arrest and PI3/AKT activation. Therefore, targeting miR-21 increases the response to radiotherapy in cancerous cells [115,129]. Simultaneous targeting of miR-21 and other miRNAs also enhances therapeutic outcomes in various cancer types (Fig. 2) [29,130–134].

Simultaneous targeting of miR-21 and different miRNAs in various cancer types. Gray and white colors represent upregulated tumor suppressor proteins and downregulated oncoproteins after cotargeting miR-21 and different miRNAs, respectively. Cas, caspase; CDK4/6, cyclin-dependent kinase 4/6; EGFR, epidermal growth factor receptor; MAPK10, mitogen-activated protein kinase 10; miRNA, microRNA; MMP, matrix metalloproteinase; ND, not determined; p-AKT, phospho-AKT; RAB2B, Ras-related protein Rab-2B; RASSF6, Ras association domain family member 6; RHOC, Ras homolog family member C; SMAD7, SMAD family member 7; SOCS6, suppressor of cytokine signaling 6; STK33, serine/threonine kinase 33; TET1, Tet methyl cytosine dioxygenase 1; TNFRSF11B, TNF receptor superfamily member 11b; TP53INP1/2, tumor protein 53-induced nuclear protein 1; TP63, tumor protein P63; TPM1, tropomyosin 1.
Using a systems biology approach and understanding the larger picture of molecular mechanisms, we can reveal which common genes and pathways are affected by both miR-21 and the other miRNAs during tumorigenesis. Besides, considering the unwanted off-target effects associated with miRNAs, it seems that the effective method to eliminate undesired consequences induced by high-dose single-miRNA treatments would be use of miRNA combinations at lower levels [135,136]. Therefore, knocking down miR-21 simultaneously targets different oncogenic pathways and enhances therapeutic efficacy of other miRNAs while preventing the toxic effects of an individual high-dose miRNA treatment. Furthermore, repression of miR-21 using anti-miRs might target gene regulatory networks that are critical to intensify and accelerate outcomes of tumor suppressor miRNAs on their target genes and pathways. For instance, we showed synergistic effects on decreasing tumor burden through simultaneous suppression of miR-21 and upregulation of miR-7 in different tumor cells. Modulation of miR-21 and miR-7 induced apoptosis through inhibition of PTEN/p-AKT (phospho-AKT) pathway [29]. In addition, suppression of miR-21 in combination with other oncomiRs induces stronger gene repression and restored the expression of their common lost tumor suppressor targets. Li et al. revealed that knockdown of miR-21 and miR-183 could significantly reduce growth and invasion in hepatocellular carcinoma through targeting SOCS6, a suppressor of cytokine signaling 6 (Fig. 2) [134]. Similarly, other studies have demonstrated the significant therapeutic potentials of miR-21 (Fig. 2), which makes this miRNA a good candidate for combination cancer treatment.
Conclusion and Future Perspectives
Cancer therapeutic resistance is an avertable phenomenon that reduces the effectiveness of various treatments. A growing number of studies have found that miRNAs are involved in drug resistance mechanisms, which are big hurdles for successful treatment of malignant cancers. Therefore, modulating the expression of these small molecules can sensitize drug-resistant tumor cells to anticancer therapies [2]. In this regard, it has been shown that downregulation of miR-21 can significantly alleviate cancer chemoresistance that has limited clinical applications of chemotherapeutic agents. In addition, chemotherapy treatment can harm the patients with different short-term and long-term side effects; therefore, combining anti-miR-21 with other agents, which results in decreasing the dosage, would hold great appeals in enhancing the anticancer efficacy while controlling for dose-dependent host toxicity. Furthermore, conventional chemotherapy mainly affects the majority of non-CSCs; however, inhibition of miR-21 combined with anticancer drugs would successfully eradicate CSCs and revoke relapse of cancer.
Although many studies have shown that modulation of miR-21 can be an opportunity to overcome drug resistance, there are still some issues related to anti-miR-21 combination therapy, which need to be addressed before its translation into clinical applications. One of the most serious obstacles to achieving optimal effectiveness is delivery of adequate and sustainable miR-21 suppressors to tumor sites. Furthermore, miR-21 suppressors and chemotherapeutic agents can cause toxicity when delivered through bloodstream. Thus, there is a need for smart carriers as promising tools to overcome their delivery problems. To this purpose, carriers could be utilized, which are resistant to nuclease degradation, selectively localized and releasing the drugs specifically at tumor sites. Various studies revealed that repression of miR-21 not only sensitizes cancer cells to chemotherapeutic agents but also enhances the efficacy of other therapeutic approaches in advanced cancer treatment. It seems that simultaneous targeting of miR-21 and other miRNAs may offer a promising strategy to successfully eradicate heterogeneous cancer cells [29,130–134]. Taken together, modulating miR-21 might be considered as a promising neoadjuvant therapy for cancer. However, further studies are needed to evaluate and optimize the performance of this therapeutic approach before translating it into the clinic.
Footnotes
Acknowledgments
All authors of the article confirm that their research is supported by an institution that is primarily involved in education and research.
Author Disclosure Statement
The authors declare that they have no potential conflicts of interest.
Funding Information
The authors acknowledge the financial support (grant no. 40297) from Ferdowsi University of Mashhad.