
Abstract
Double-stranded RNAs (dsRNAs) longer than 30 bp have not been considered desirable RNA interference (RNAi) triggering structures in mammalian cells as they nonspecifically activate innate immune response. However, in earlier studies, not only dsRNA length but also 5′-triphosphate moiety produced by in vitro transcription might have affected the stimulation of innate immune system. Herein, using chemically synthesized long dsRNAs without 5′-triphosphate, we elucidated direct relationship between length of dsRNAs and innate immune stimulation. First, we found that blunt-ended, chemically synthesized 38/40–60 bp-long dsRNAs induced retinoic acid-inducible gene I (RIG-I)-mediated innate immune response, which was suppressed by the introduction of the 2-nt 3′ overhang structure. Surprisingly, we discovered that RIG-I activation by these long dsRNAs is also sequence dependent, and the sequence composition at dsRNA termini is important for RIG-I activation. In addition, we identified that long dsRNAs over 38 bp could elicit single- or dual-target gene silencing in a Dicer-independent manner. Taken together, our findings may serve as guidelines to develop an immunostimulatory RNAi trigger to exploit host's innate immune system, as well as a specific dual-gene targeting RNAi therapeutics platform without nonspecific innate immune stimulation by RIG-I.
Introduction
RNA interference (RNAi) is a natural phenomenon that drives inhibition of translation or degradation of mRNA in a sequence-specific manner [1,2]. RNAi-mediated gene silencing can be initiated by 19–23 bp-long double-stranded RNA (dsRNA) with 2-nucleotide (nt) 3′ overhangs, termed small interfering RNA or siRNA, generated by Dicer cleavage or chemical synthesis [3–6]. Although RNAi mechanism exists in plants, insects, and fungi [7–9], the silencing of disease-causing genes by RNAi in mammalian cells led to the development of RNAi therapeutics [10,11]. Until now, four siRNA-based drugs, Patisiran, Givosiran, Vutrisiran, and Inclisiran, were approved by the US Food and Drug Administration (FDA) or European Medicines Agency (EMA), and many more RNAi therapies are expected to enter the market in coming years [12–15].
Since the introduction of the original siRNA structure, which harbors 19 bp-long dsRNA region with 2-nt 3′ overhangs on both ends, several RNAi triggers that have unique structural features and yet show comparable efficiency compared with original siRNA structure have been reported [3].
However, when devising complex RNAi structures with multifunctional features such as multiple-target gene silencing [16–18], the possibility of innate immune stimulation by complex RNA structures such as long dsRNA needs to be carefully addressed [19–23]. Especially, long dsRNAs are known to be recognized by several pattern recognition receptors (PRRs) as pathogen-associated molecular patterns (PAMPs) and elicit an innate immune response because mammalian cells recognize dsRNAs as intermediate of viral replication [24,25].
Protein kinase R (PKR) recognizes dsRNAs longer than 30 bp, inhibiting protein synthesis by phosphorylation of eIF2α [26,27]. Toll-like receptor 3 (TLR3) also recognizes 40–50 bp-long dsRNA or polyinosine-polycytidylic acid (poly I:C), initiating the production of interferon (IFN) or proinflammatory cytokines [28–30].
Melanoma differentiation-associated protein 5 (MDA5) and retinoic acid-inducible gene I (RIG-I) also detect long dsRNAs in the cytosol and elicit IFN response [31]. Despite structural similarities, they recognize different structural features of dsRNAs. MDA5 mainly recognizes dsRNAs longer than 2,000 bp; however, RIG-I senses dsRNAs shorter than 300 bp [32]. In addition, blunt-ended 23–27 bp-long dsRNAs, and single- or dsRNAs with 5′ triphosphate produced by viral replication or in vitro transcription, are well known as a specific PAMP against RIG-I due to the difference of C terminal domain between RIG-I and MDA5 [33–36].
Earlier studies reported that long dsRNAs over 30 bp activate IFN system and induce nonspecific gene silencing [10,30]; however, in these studies, not only dsRNA length but also 5′-triphosphate produced by in vitro transcription might have contributed the innate immune response observed. This motivated us to elucidate the direct relationship between dsRNA length and innate immune stimulation using chemically synthesized 38/40–60 bp-long dsRNAs without 5′-triphosphate. Although we have previously reported that a chemically synthesized 38 bp-long blunt-ended dsRNA can trigger dual-gene silencing along with innate immune stimulation [37], the structure-activity relationship of long dsRNAs with various lengths was not thoroughly analyzed.
Herein, we systematically studied the RNAi potency and immunogenicity of chemically synthesized long dsRNAs with various lengths and the end structures. First, we found that 38/40–60 bp-long dsRNAs showed gene silencing in Dicer-independent manner, and the gene silencing efficacy was enhanced by the addition of two 2-nt 3′ overhangs. Second, we found that blunt-ended 38/40–60 bp-long dsRNAs induced RIG-I-mediated innate immune response, and it could be suppressed by the introduction of 2-nt 3′ overhangs. Unexpectedly, we discovered the RIG-I-activating sequence motif at long dsRNA termini, which shows that RIG-I activation by long dsRNAs is sequence dependent.
Materials and Methods
RNAs
Chemically synthesized RNA oligonucleotides to make up 19 bp-long siRNAs targeting Luciferase or green fluorescent protein (GFP) and dsRNAs longer than or equal to 38 bp were purchased from Dharmacon, Inc. (UK). The other RNA oligonucleotides shorter than or equal to 27-nt were purchased from Bioneer Co. Ltd (Republic of Korea) (Supplementary Tables S1–S3, and Supplementary Fig. S1). All oligonucleotides used in this study do not have any chemical modification and each single-stranded oligonucleotide longer than or equal to 27-nt was purified by HPLC.
Cells lines
HeLa (CCL-2) and T98G (CRL-1690) cell line were purchased from American Type Culture Collection (ATCC, USA) and cultured in Dulbecco's modified Eagle's medium (DMEM, 11965092; Corning) supplemented with 10% fetal bovine serum (FBS, 16000044; Gibco), 100 U/mL penicillin, and 100 mg/mL streptomycin (15140122; Gibco). HAP1 wild-type (WT) (C631) and HAP1 Dicer knockout (KO) (HZGHC000909c012) cell line were purchased from Horizon Discovery Ltd. (UK) and cultured in Iscove's modified Dulbecco's medium (12440053; Gibco) supplemented with 10% FBS (16000044; Gibco), 100 U/mL penicillin, and 100 mg/mL streptomycin (15410112; Gibco).
RAW-Lucia interferon-stimulated gene (ISG) (rawl-isg) and RAW-Lucia ISG-KO-RIG-I (rawl-korigi) cell lines are mainly used in this study. They were purchased from InvivoGen (USA) and cultured in DMEM (11965092; Corning) supplemented with 10% FBS (16000044; Gibco), 100 μg/mL Normocin (ant-nr-1; InvivoGen), and 200 μg/mL Zeocin (ant-zn-05; InvivoGen).
Gene silencing analysis
HeLa cells were plated in 24-well or 48-well plates 24 h before transfection at 30% confluency in a complete medium without antibiotics. Cells in 24-well plates were treated with 19, 40, 50, or 60 bp-long dsRNAs targeting Survivin or Lamin A/C-glyceraldehyde 3-phosphate dehydrogenase (GAPDH), and cells in 48-well plates were treated with 19, 38, 50, or 60 bp-long dsRNAs targeting Luciferase-GFP with concentrations ranging from 0.001 to 10 nM using Lipofectamine RNAiMAX (13778150; Invitrogen).
For Survivin, Lamin A/C, and GAPDH mRNA analysis, total RNA was extracted using Tri-RNA Reagent (FATRR 001; Favorgen) after 24 h post-transfection, and 0.5 μg of total RNA was used for complementary DNA (cDNA) synthesis using the high-capacity cDNA reverse transcription kit (4368813; Applied Biosystems) according to the manufacturer's protocol. cDNA was analyzed by quantitative reverse transcription-polymerase chain reaction (qRT-PCR) using the CFX96 Touch Real-Time PCR Detection System (Bio-Rad).
Primers for the target gene were mixed with SYBR Green PCR Master Mix (RR420A; TaKaRa). The target genes and internal control mRNA levels were determined by using the relative standard curve quantitation method. The primer sequences for each gene are described in Supplementary Table S4. For luciferase and GFP expression analysis, luciferase activity was measured through a plate reader (VICTORX2; PerkinElmer) at a 1-s reading time using luciferase assay kit (E1500; Promega), and GFP expression was also measured using the same instrument at a 1-s reading time. GraphPad Prism 8 was used to fit a dose–response curve to determine IC50 values.
HAP1 WT or HAP1 Dicer KO cells were plated in 24-well plates 24 h before transfection at 30% confluency in a complete medium without antibiotics. Those cells were transfected with 0.3 nM of 19, 40, 50, or 60 bp-long dsRNAs targeting Survivin or 1 nM of 19, 40, 50, or 60 bp-long dsRNAs targeting Lamin A/C-GAPDH using Lipofectamine RNAiMAX (13778150; Invitrogen). After 24 h post-transfection, target gene and internal control mRNA level were analyzed by qRT-PCR following the protocol described above. The primer sequences for each gene are described in Supplementary Table S4.
Cytokine expression analysis
T98G cells were plated in 24-well plates 24 h before transfection at 30% confluency in a complete medium without antibiotics. Cells were treated with 10 nM of 19, 40, 50, or 60 bp-long dsRNAs targeting Survivin using Lipofectamine RNAiMAX (13778150; Invitrogen) and harvested 24 h after transfection, or cells were treated with 10 nM of 19, 38/40, 50, or 60 bp-long dsRNAs targeting Luciferase-GFP or Lamin A/C-GAPDH using Lipofectamine RNAiMAX (13778150; Invitrogen) and harvested 12 h after transfection. mRNA level of IFIT1, CXCL10, ISG15, tumor necrosis factor (TNF)-α, and IL-1β was analyzed using qRT-PCR following the protocol described above. The primer sequences for each gene are described in Supplementary Table S4.
Reporter cell assay
RAW-Lucia ISG and RAW-Lucia ISG-KO-RIG-I cells stably express an IFN regulatory factor–inducible Lucia luciferase reporter construct. Both cells were seeded at 1 × 105 cells per well in a 96-well plate and treated with 19, 38/40, 50, or 60 bp-long dsRNAs using Lipofectamine RNAiMAX (13778150; Invitrogen) at the same time. After 24 h, culture supernatant was collected to measure luciferase using QUANTI-Luc (rep-qlc2; InvivoGen) to detect the level of interferon regulatory factor (IRF) activity with a plate reader (VICTORX2; PerkinElmer) at a 1-s reading time.
Results
Chemically synthesized long dsRNAs trigger efficient gene silencing in Dicer-independent manner
We designed three groups of chemically synthesized long dsRNAs targeting one (Survivin) or two genes (Luciferase-GFP or Lamin A/C-GAPDH) without chemical modifications. Forty, 50, or 60 bp-long dsRNAs targeting Survivin were made by extending a 19 bp-long siRNA targeting Survivin, while 38/40, 50, or 60 bp-long dsRNAs targeting Luciferase-GFP or Lamin A/C-GAPDH were made by combining two 19 bp-long or extended dsRNAs targeting each gene (Supplementary Table S1, Supplementary Fig. S1). All dsRNAs showed IC50 values lower than 0.5 nM for the target genes regardless of length or end structure (Table 1, Supplementary Fig. S2).
IC50 Assessment of Chemically Synthesized Long Double-Stranded RNAs in HeLa Cell
Comparison of single- or dual-gene silencing potency by chemically synthesized long dsRNAs targeting Survivin, Luciferase-GFP, or Lamin A/C-GAPDH with blunt ends or two 2-nt 3′ overhangs. mRNA levels of target genes were analyzed 24 h after transfection. The mean of 2–3 independent experiments was used to calculate IC50 values.
dsRNA, double-stranded RNA; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; GFP, green fluorescent protein.
Survivin-targeting 40 bp-long dsRNA with blunt ends showed IC50 value similar to 19 bp-long blunt-ended siRNA, but 50 and 60 bp-long dsRNA with blunt ends showed two to three times reduced gene silencing potency (Table 1). Among long blunt-ended dsRNAs targeting Luciferase-GFP, 50 bp-long dsRNA showed the best gene silencing efficacy; however, it showed threefold to fivefold reduction compared with the mixture of 19 bp-long blunt-ended siRNA (Table 1). In the case of long dsRNAs targeting Lamin A/C-GAPDH, 60 and 40 bp-long dsRNAs showed the most potent gene silencing for Lamin A/C and GAPDH, respectively, despite a reduced efficacy compared to 19 bp-long blunt-ended siRNA (Table 1).
Also, as previously observed [37], the introduction of two 2-nt 3′ overhangs enhanced RNAi-mediated gene silencing induced by all dsRNAs targeting Survivin or Lamin A/C-GAPDH (Table 1, Supplementary Fig. S2). In contrast, no improvement in gene silencing efficiency was observed with the introduction of two 2-nt 3′ overhangs in dsRNAs targeting Luciferase-GFP (Table 1). Taken together, chemically synthesized 38/40–60 bp-long dsRNAs are able to induce single- or dual-target gene silencing and the optimal length of dsRNAs varies depending on the sequence. In addition, the introduction of two 2-nt 3′ overhangs can enhance RNAi; however, it also depends on target gene or dsRNA sequence.
Surprisingly, long dsRNA-induced gene silencing for Survivin or Lamin A/C-GAPDH was not abrogated under Dicer-deficient condition, regardless of dsRNA length or 3′ end structure (Fig. 1A–C), suggesting that chemically synthesized dsRNAs longer than 38 bp could trigger target gene silencing in Dicer-independent manner.
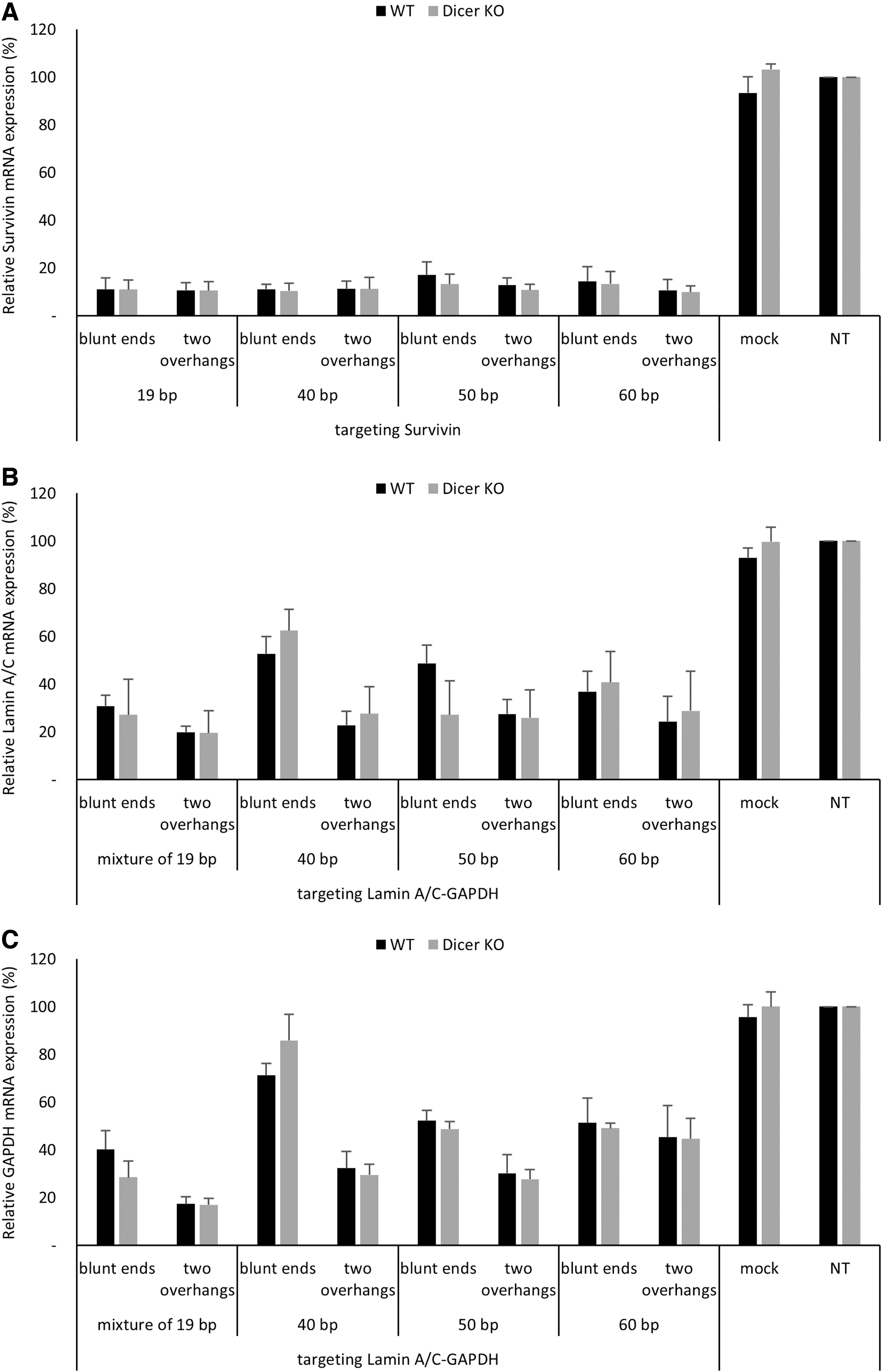
Dicer-independent target gene silencing triggered by chemically synthesized long dsRNAs. Gene silencing of chemically synthesized long dsRNAs with blunt ends or two 2-nt 3′ overhangs was analyzed by qRT-PCR in HAP1 WT and HAP1 Dicer KO cell line 24 h after transfection.
Long dsRNAs with blunt ends elicit innate immune responses
To evaluate immunogenicity of long dsRNAs, all dsRNAs were transfected into T98G cells and cytokine expression associated with innate immune response was analyzed. Poly I:C was transfected as a positive control and 19 bp-long siRNAs with blunt ends or 2-nt 3′ overhangs were used as negative controls of innate immune stimulation. We analyzed the induction of two ISGs: IFIT1 and ISG15, one chemokine: CXCL10, and two proinflammatory cytokines: TNF-α and IL-1β. Thirty eight or 40 bp-long dsRNAs targeting Survivin or Luciferase-GFP showed a strong induction of IFIT1, CXCL10, and ISG15 expression, which was comparable or greater than seen with poly I:C treatment (Fig. 2A, B).
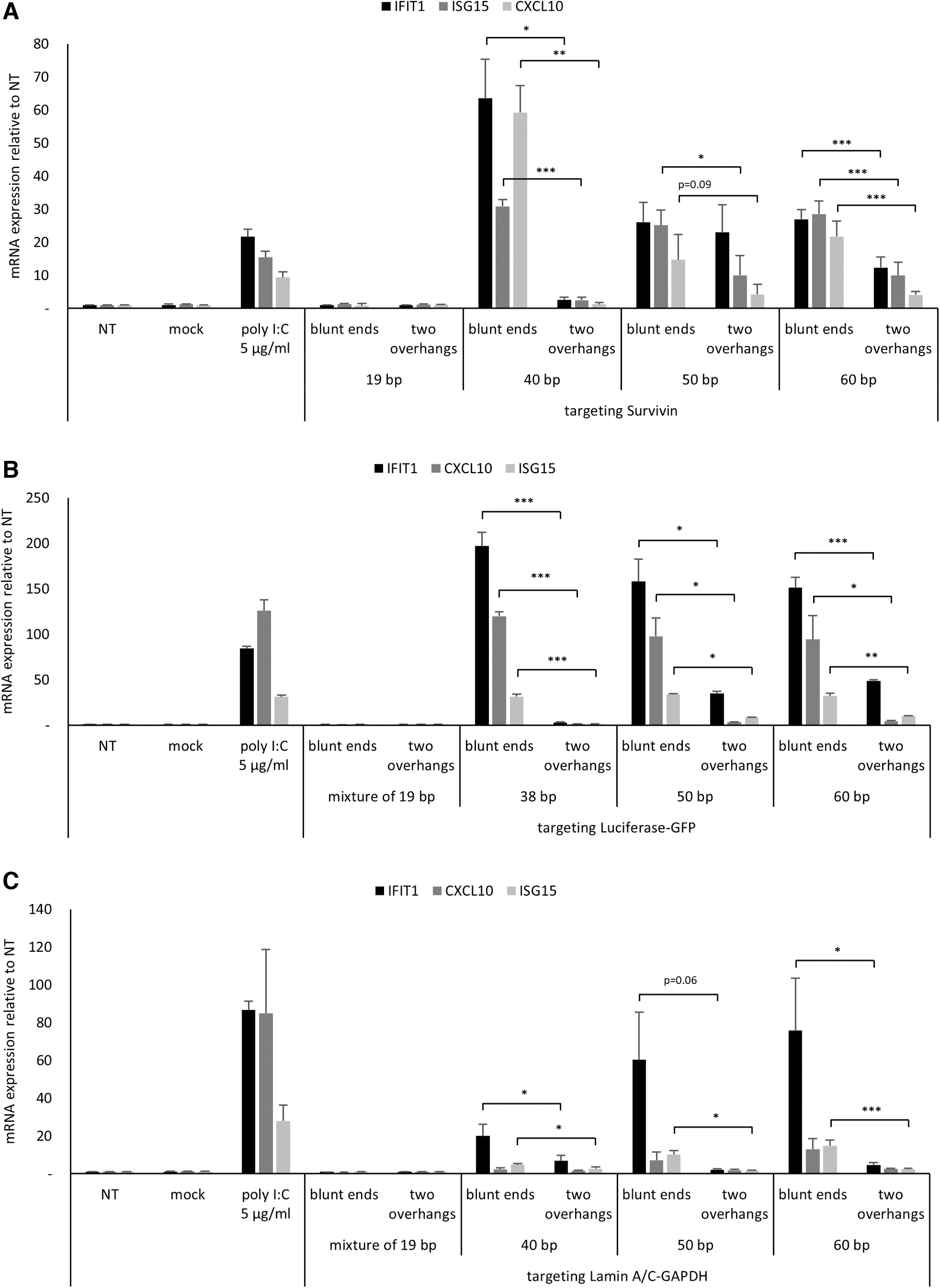
Innate immune response triggered by chemically synthesized long dsRNAs in T98G cell line. Innate immune activation triggered by three kinds of chemically synthesized long dsRNAs in T98G cell line.
Forty bp-long dsRNAs targeting Lamin A/C-GAPDH also showed immune activation, although it was weaker compared with other 38/40 bp-long dsRNAs (Fig. 2C). This result demonstrates that chemically synthesized long dsRNAs with blunt ends can induce innate immune response and indicates the presence of a particular PRR, which recognizes these molecular patterns. Also, despite the similarity in length and end structure, three dsRNAs targeting different genes showed varying extent of innate immune responses, suggesting that immunogenicity of long dsRNAs may also depend on the sequence.
Interestingly and against our expectation, 50 and 60 bp-long dsRNAs targeting Survivin or Luciferase-GFP showed IFIT1 and CXCL10 induction level lower compared with 40/38 bp-long dsRNAs (Fig. 2A, B). In contrast, 50 and 60 bp-long dsRNAs targeting Lamin A/C-GAPDH showed dsRNA length-dependent increase of induction level of all three cytokines (Fig. 2C).
Unlike the cytokines described above, proinflammatory cytokines such as TNF-α and IL-1β were only marginally induced by Survivin or Lamin A/C-GAPDH-targeting 40–60 bp-long blunt-ended dsRNAs (Supplementary Fig. S3A, C). Only blunt-ended long dsRNAs targeting Luciferase-GFP induced marginal TNF-α and IL-1β expression (Supplementary Fig. S3B). Taken together, chemically synthesized 38/40–60 bp-long dsRNAs with blunt ends trigger innate immune response, and the level and profile of immune-related gene activation pattern are sequence dependent.
Interestingly, the presence of 2-nt 3′ overhangs on both ends of long dsRNAs showed significantly reduced expression of ISGs and CXCL10 involved in innate immune stimulation in T98G cell line (Fig. 2A–C). Especially, the addition of 2-nt 3′ overhangs on both ends of 38/40 bp-long dsRNAs completely suppressed innate immune stimulation regardless of dsRNA sequence (Fig. 2A–C). Marginal expression of TNF-α and IL-1β, which was elicited by blunt-ended long dsRNAs targeting Luciferase-GFP, was also reduced by the addition of 2-nt overhangs (Supplementary Fig. S3B).
These results clearly suggest that the addition of 2-nt 3′ overhangs on both ends can control the innate immune response triggered by blunt-ended long dsRNAs. Considering the previous reports that the addition of 2-nt 3′ overhangs blocked RIG-I activation initiated by blunt-ended dsRNAs with 23–27 bp [35], we hypothesized that RIG-I could be the PRR involved in sensing these long dsRNAs with blunt ends to trigger downstream induction of innate immune responses.
Long dsRNAs with blunt ends trigger RIG-I-mediated innate immune response
It is known that macrophage cell lines are rich in PRRs [38]. Therefore, RAW 264.7 macrophage-derived RAW-Lucia ISG cell line, which expresses IRF-inducible Lucia luciferase gene, and RAW-Lucia ISG-KO-RIG-I, which is the corresponding RIG-I KO cell line, were used to verify if RIG-I is a major PRR, which recognizes chemically synthesized, blunt-ended long dsRNAs as PAMPs. Similar to the previous results in T98G cells, 38/40–60 bp-long, blunt-ended dsRNAs targeting Survivin or Luciferase-GFP triggered strong IRF-mediated immune responses in RAW-Lucia ISG cells (Fig. 3A, B). However, none of these dsRNAs elicited any IRF-dependent immune response in RAW-Lucia ISG-KO-RIG-I cells, suggesting that RIG-I is a major PRR to recognize blunt-ended 38/40–60 bp-long dsRNAs in this cell type (Fig. 3A, B).
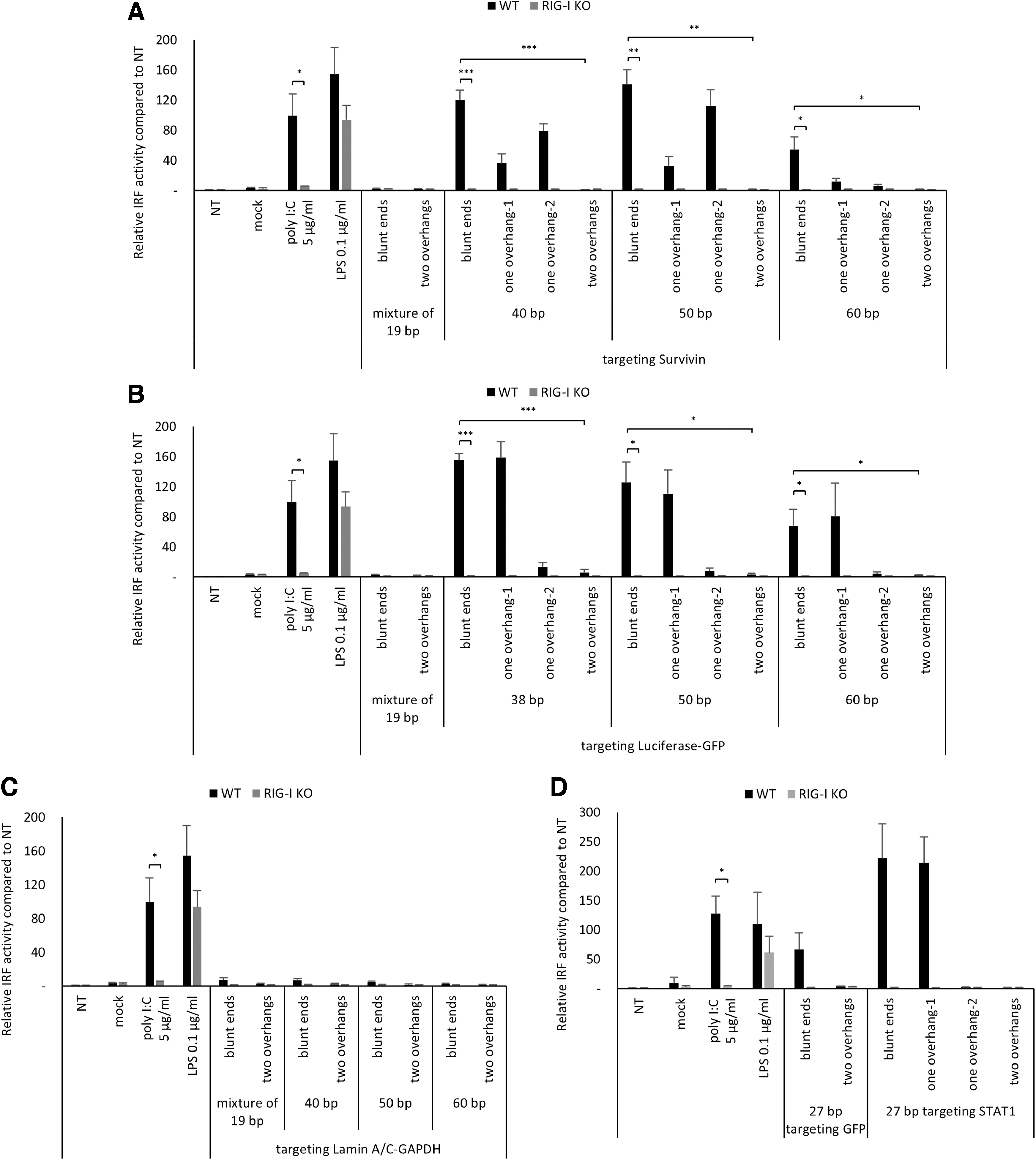
RIG-I-mediated innate immune response triggered by chemically synthesized long dsRNAs in mouse macrophage cell line. Innate immune response induced by chemically synthesized 38/40, 50, and 60 bp-long dsRNAs with blunt ends or 2-nt 3′ overhangs in RAW-Lucia ISG and RAW-Lucia ISG-KO-RIG-I which expresses IRF-inducible Lucia luciferase gene.
Addition of 2-nt 3′ overhangs on both ends of Survivin or Luciferase-GFP targeting dsRNAs completely abrogated the IRF-mediated innate immune responses (Fig. 3A, B), which further corroborate the RIG-I-mediated innate immune stimulation by these dsRNAs [22,34,35]. Taken together, we conclude that RIG-I is a major PRR that initiates IRF-dependent innate immune response by chemically synthesized, blunt-ended 38/40–60 bp-long dsRNAs.
We noted that, unlike the results from T98G cells, 40, 50, and 60 bp-long blunt-ended dsRNAs targeting Lamin A/C-GAPDH did not induce IRF-dependent immune response in RAW-Lucia ISG cells (Fig. 3C). Considering that RIG-I displays sequence variation depending on species or even individuals in the same species [39], cell line-dependent immune stimulation by dsRNAs targeting Lamin A/C-GAPDH may originate from the difference between human and mouse RIG-I.
Long dsRNAs with blunt ends activate RIG-I in a structure- and sequence-dependent manner
To further understand whether 2-nt 3′ overhangs on both ends are necessary to regulate RIG-I-mediated immune response, IRF-mediated immune stimulation by long dsRNAs with single 2-nt 3′ overhang was also measured in RAW-Lucia ISG and RAW-Lucia ISG-KO-RIG-I cells (Supplementary Table S1).
The addition of a single 2-nt overhang on either end of a 60 bp-long dsRNA targeting Survivin (one overhang-1 and overhang-2) significantly suppressed IRF stimulation observed with the blunt end structure, although not complete (Fig. 3A), reflecting the already reduced immune stimulation by a 60 bp-long blunt-ended dsRNA compared with that of 40 and 50 bp-long dsRNAs. Forty and 50 bp-long dsRNAs targeting Survivin with one overhang (one overhang-1 and overhang-2) showed reduced IRF activation compared with that of blunt-ended one, although the level of reduction is different (Fig. 3A).
In addition, the position of 3′ overhang resulted in completely different effects on innate immune stimulation triggered by 38–60 bp-long dsRNAs targeting Luciferase-GFP in RAW-Lucia ISG cells, irrespective of dsRNA length (Fig. 3B). When a single 2-nt 3′ overhang was added on the duplex end of GFP-targeting sequence, the IRF-mediated innate immune response was considerably reduced (one overhang-2, Fig. 3B). In contrast, there was no reduction of IRF activation when single 2-nt 3′ overhang was added on the other end of dsRNA, which is the end of Luciferase-targeting sequence (one overhang-1, Fig. 3B). As one overhang-1 and overhang-2 are structurally identical, these results indicate that the immunostimulation seen with Luiferase-GFP-targeting dsRNAs may originate from sequence features, specifically, the dsRNA sequence targeting GFP.
As expected, and further confirming the involvement of RIG-I pathway, dsRNAs targeting Survivin or Luciferase-GFP with one 2-nt 3′ overhang (one overhang-1 and overhang-2) failed to elicit any innate immune response in RAW-Lucia ISG-KO-RIG-I cells, the RIG-I knockout cell line (Fig. 3A, B).
We tested another blunt-ended 27 bp-long dsRNA targeting GFP or STAT1 reported by Marques et al. as RIG-I agonists [35]. Both dsRNAs showed induction of innate immune response in RAW-Lucia ISG; however, dsRNAs targeting GFP induced an innate immune response about twofold lesser than STAT1-targeting dsRNAs (Fig. 3D). Furthermore, full suppression of the immunogenicity was observed when 2-nt 3′ overhang was added on one end (one overhang-2), but not the other end (one overhang-1) (Fig. 3D), similar to what we observed with dsRNAs targeting Luciferase-GFP (Fig. 3B). Overall, these results demonstrate that sequence composition of chemically synthesized 27–60 bp-long blunt-ended dsRNAs plays an important role in RIG-I activation and downstream innate immune response.
To further clarify the sequence-dependent RIG-I activation by long dsRNAs, a 20 bp-long dsRNA targeting Lamin A/C or GAPDH was combined with the 20 bp-long dsRNA targeting GFP or Luciferase to generate four kinds of 40 bp-long chimeric dsRNAs, because immune stimulation profile of dsRNAs targeting Luciferase-GFP indicated the presence of immunostimulatory motif in GFP-targeting sequence (Supplementary Table S2 and Fig. 3B). We found that 40 bp-long chimeric dsRNAs with 20 bp-long GFP-targeting sequence induced innate immune response similar to 40 bp-long blunt-ended dsRNAs targeting Luciferase-GFP in T98G cells and RAW-Lucia ISG cells; however, the other chimeric dsRNAs without GFP-targeting sequence failed to induce innate immune response in both cells (Fig. 4A, B).
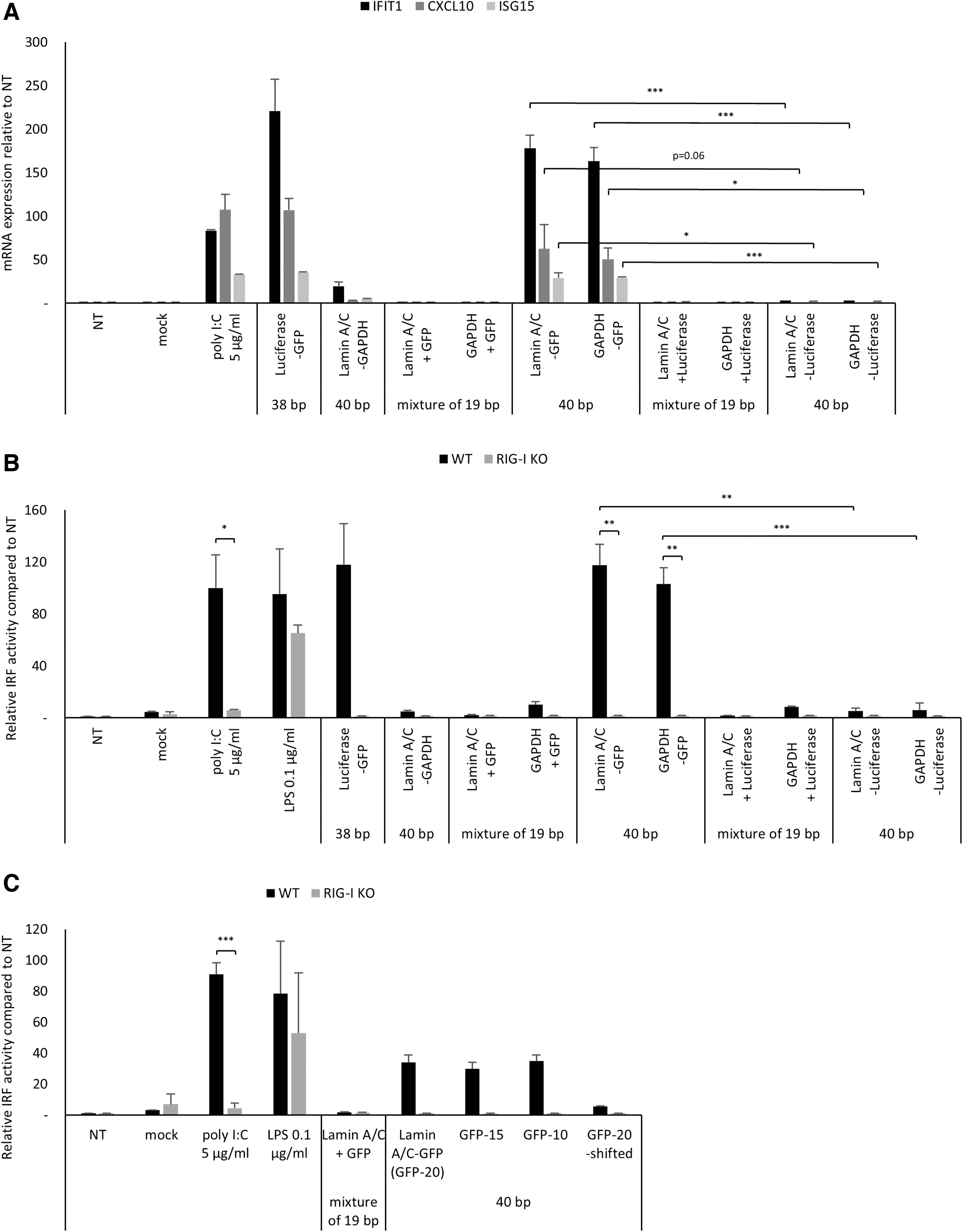
RIG-I activation in structure- and sequence-dependent manner. Innate immune stimulation by chemically synthesized 40 bp-long chimeric dsRNAs with or without GFP-targeting 20 bp-long dsRNA motif.
As expected, these chimeric dsRNAs failed to elicit IRF activation in RAW-Lucia ISG KO RIG-I cells (Fig. 4B). Taken together, we show that RIG-I activation by long dsRNAs can be controlled in a sequence-dependent manner, by introducing an immunogenic dsRNA sequence motif. We would like to note that, in addition to the immunostimulatory sequence motif, the structural requirements of dsRNAs such as duplex length and the end structure are also important factors to elicit RIG-I-mediated innate immune response, as the mixture of 19 bp-long siRNAs targeting Luciferase or GFP failed to induce an innate immune response in both T98G and RAW-Lucia ISG cells (Fig. 4A, B).
Sequence composition at blunt-end termini of dsRNAs is important for sequence-dependent RIG-I activation
Considering that Watson-Crick base pair at the duplex termini is important to initiate RIG-I-mediated immune response [40], we hypothesized that sequence composition at dsRNA termini determines sequence-dependent RIG-I activation by long dsRNAs. To examine this hypothesis, blunt-ended 40 bp-long chimeric dsRNAs harboring terminal 15 or 10 bp-long sequence from the GFP-targeting dsRNAs were designed (GFP-15 and GFP-10, Supplementary Table S3).
In addition, the 20 bp-long dsRNA sequence targeting GFP was placed at the center of 40 bp-long dsRNAs targeting Lamin A/C-GAPDH with blunt ends, maintaining the sequence of dsRNA termini similar to 40 bp-long dsRNAs targeting Lamin A/C-GAPDH (GFP-20 shifted, Supplementary Table S3). GFP-10 and GFP-15 induced innate immune response similar to that of original 40 bp-long dsRNA targeting Lamin A/C-GFP with complete 20 bp-long dsRNA targeting GFP (Fig. 4C). In contrast, when the 20 bp-long GFP-targeting sequence was removed from the termini (GFP-20-shifted), RIG-I-mediated innate immune stimulation was abrogated in RAW-Lucia ISG cells (Fig. 4C). Taken together, we conclude that the sequence composition at dsRNA termini determines the activation of RIG-I by chemically synthesized long dsRNAs.
Discussion
Our group previously reported that chemically synthesized 38 bp-long dsRNAs with blunt ends can induce dual-target gene silencing as well as innate immune response in mammalian cells [37]. However, immunogenicity and gene silencing potency of chemically synthesized 38 bp or longer dsRNA without 5′-triphosphate have not been comprehensively analyzed.
In this study, we performed a detailed analysis of the innate immune response triggered by chemically synthesized 38/40–60 bp-long dsRNAs in T98G and RAW-Lucia ISG cells. In agreement with the earlier studies on innate immune stimulation by 27 bp-long blunt-ended dsRNAs [35], we found that 38/40–60 bp-long blunt-ended dsRNA also evoked innate immune response, despite the absence of 5′-triphosphate (Figs. 2 and 3). Especially, using RIG-I KO cells, we unveiled that RIG-I is a major PRR that recognizes 38/40–60 bp-long dsRNA with blunt ends (Fig. 3).
Of note, the introduction of 2-nt 3′ overhangs completely blocked the innate immune stimulation by blunt-ended long dsRNAs targeting Survivin or Luciferase-GFP regardless of their length in RAW-Lucia ISG cells (Fig. 3A, B). However, in T98G cells, the immune response induced by 50 and 60 bp-long blunt-ended dsRNAs targeting Survivin or Luciferase-GFP was only partially blocked by the introduction of 2-nt 3′ overhangs (Fig. 2A, B).
These data suggest that other PRRs than RIG-I such as PKR or TLR3 could also be involved in innate immune stimulation by long dsRNAs, longer than or equal to 50 bp depending upon cell type or cellular conditions [26,41,42]. In addition, 40–60 bp-long blunt-ended dsRNAs targeting Lamin A/C-GAPDH showed innate immune stimulation in T98G, not in RAW-Lucia ISG cells (Figs. 2C and 3C), probably due to the difference between human and mouse RIG-I [39]. In the future, cytokine assessments in protein level are warranted to obtain more detailed immunogenicity profile of chemically synthesized long dsRNAs.
Although many single- or dsRNA structures have been reported to activate RIG-I [43], there are only a few reports regarding sequence-dependent RIG-I activation and they focus on the sequence of single-stranded RNA (ssRNA). Saito et al. reported that poly-U/UC ssRNA motif with 5′-triphosphate derived from hepatitis C virus could elicit RIG-I-mediated innate immune response in a sequence-dependent manner [44]. Similarly, U/A-rich region in the 3′ UTR of influenza cRNA can elicit RIG-I-mediated IFN-β induction without 5′-triphosphate [45].
Herein, we found that RIG-I activation requires not only specific molecular patterns such as the blunt-ended long dsRNA structure but also particular dsRNA sequence features. Especially, studies with 40 bp-long blunt-ended chimeric dsRNAs containing 20 bp-long dsRNA targeting GFP showed that the sequence composition at duplex termini is important to elicit sequence-dependent RIG-I activation by long dsRNAs.
It has been reported that RIG-I activation by dsRNA shorter than 200 bp requires the presence of stimulatory motif such as 5′-triphosphate, which increases the affinity of dsRNA for RIG-I [46]. It is possible that like 5′-triphosphate, certain sequence motifs at duplex termini may help in the binding of dsRNA to RIG-I, leading to a potent RIG-I activation. Further studies are warranted to clearly understand the effect of dsRNA sequence composition on RIG-I binding or activation.
Considering that several groups showed type I IFN-related cytokine release such as CXCL10 and IFN-β confirmed in in vitro assay was also observed in in vivo animal model, resulting in anticancer and antiviral effect [47,48], we believe that our in vitro finding also can be utilized to predict in vivo pharmacology in regard to immunogenicity.
However, the prediction of systemic adverse effect such as cytokine storm through in vitro assay is challenging due to species, cell type, donor, or assay system-dependent immune response [49–51]. In 2006, TGN1412, a superagonist anti-CD28 monoclonal antibody, caused serious cytokine storm in six healthy volunteers in phase I clinical trial, although no adverse effect was observed in preclinical safety test using in vitro cytokine release assays (CRAs), using human peripheral blood mononuclear cell (hPBMC) and administration of TGN1412 to cynomolgus macaques [50].
Nonetheless, hPBMC is widely used for CRAs and whole blood (WB) is another option for in vitro CRAs [52]. WB CRA is considered as a system that better reflects in vivo environment, including the presence of several factors in blood than hPBMC [50]. However, this system showed low level of cytokines and low frequency of response depending on donors, requiring larger group size than hPBMC CRAs [50]. In addition, four cytokines, IL-2, TNF-α, IL-6, and IFN-γ, were reported as a panel of cytokines that can be analyzed through Luminex or Meso Scale Discovery platforms to predict in vivo cytokine storm or cytokine release syndrome [52]. In the future, in vitro CRA might provide us the insight to predict in vivo side effects triggered by chemically synthesized long dsRNAs.
In addition to the immunogenicity, we also assessed the RNAi efficiency of these chemically synthesized long dsRNAs. Through IC50 calculations, we confirmed that the long dsRNAs showed similar or slightly reduced gene silencing compared with 19 bp-long siRNAs. Introduction of two 2-nt 3′ overhangs enhanced gene silencing potency of long dsRNAs targeting Survivin or Lamin A/C-GAPDH regardless of dsRNA length (Table 1). We believe that like 19 + 2 conventional siRNA, the presence of 2-nt 3′ overhangs on dsRNA may allow preferential loading of guide strand into Ago2, enhancing its silencing efficiency [53,54]. In addition, using Dicer KO cells, we showed that Dicer is not required for RNAi by chemically synthesized 38/40–60 bp-long dsRNAs, expanding the diversity of Dicer-independent RNAi triggers [16,55].
RIG-I activation along with target gene silencing is a promising therapeutic approach against cancer and viral diseases to exploit host antitumor immunity through the release of IFN or by inducing apoptosis of cancer cells [47,48,56]. Therefore, we believe that our chemically synthesized blunt-ended dsRNAs, which simultaneously target disease-associated genes and activate RIG-I-mediated innate immune response, could be a novel therapeutic platform for the development of anticancer or antiviral drugs. We also showed that chemically synthesized 38/40 bp-long dsRNAs with 3′ overhangs could be an RNAi therapeutic platform that simultaneously targets two genes without the activation of the innate immune system [57,58].
In conclusion, the structure–function relationships of long dsRNAs derived from this study should provide important information for researchers to develop novel RNAi-based drugs in the future.
Footnotes
Author Disclosure Statement
D.K.L., Y.G.K., J.Y.C., and P.D. are employees and shareholders of OliX Pharmaceuticals, Inc. The other authors have no competing financial interests to disclose.
Funding Information
This work was supported by a Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2017R1D1A1B03036001).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.