
Abstract
A human peripheral blood mononuclear cell (PBMC)-based assay was developed to identify antisense oligonucleotide (ASO) with the potential to activate a cellular innate immune response outside of an acceptable level. The development of this assay was initiated when ISIS 353512 targeting the messenger ribonucleic acid for human C-reactive protein (CRP) was tested in a phase I clinical trial, in which healthy human volunteers unexpectedly experienced increases in interleukin-6 (IL-6) and CRP. This level of immune stimulation was not anticipated following rodent and nonhuman primate safety studies in which no evidence of exaggerated proinflammatory effects were observed. The IL-6 increase induced by ISIS 353512 was caused by activation of B cells. The IL-6 induction was inhibited by chloroquine pretreatment of PBMCs and the nature of ASOs suggested that the response is mediated by a Toll-like receptor (TLR), in all likelihood TLR9. While assessing the inter PBMC donor variability, two classes of human PBMC responders to ISIS 353512 were identified (discriminator and nondiscriminators). The discriminator donor PBMCs were shown to produce low level of IL-6 after 24 h in culture, in the absence of ASO treatment. The PBMC assay using discriminator donors was shown to be reproducible, allowing to assess reliably the immune potential of ASOs by comparison to known benchmark ASO controls that were previously shown to be either safe or inflammatory in clinical trials. Clinical Trial registration numbers: NCT00048321 NCT00330330 NCT00519727.
Introduction
Antisense oligonucleotides (ASOs) are short, synthetic single strands of nucleic acids with phosphorothioate linkage that hybridize to messenger ribonucleic acid (mRNA) through Watson-Crick base pairing [1]. They are designed to alter the expression of a protein by selectively binding to the RNA that encodes the targeted protein [2]. In recent years, ASOs have been used as therapeutic agents to inhibit the expression of a specific target mRNA transcript involved in a growing range of diseases [3], several of them, Waylivra (volanesorsen), Kynamro (mipomersen), and Tegsedi (inotersen), have received regulatory acceptance for commercialization [4–6].
However, despite ASOs being generally well tolerated by human volunteers and patients [7], some phosphorothioate containing ASOs have been shown to elicit nonspecific and proinflammatory effects of varying intensity in a sequence-specific manner [8]. A number of modifications of the ASO designs have been introduced such as incorporation of 2′-O-methoxyethyl (MOE) and systematic methylation of cytosine to mitigate these proinflammatory effects. Despite those improvements, a small fraction of sequences examined in rodents has exhibited a higher level of proinflammatory effects [9].
Those proinflammatory effects have been linked to a growing network of receptors and coreceptor involved in the recognition of both DNA and RNA oligonucleotides (ODNs) [9–13], including one family of the cell surface and endosome-associated receptor, Toll-like receptors (TLRs), which has been shown to recognize synthetic nucleic acids [14–16]. One of the members of this family, TLR9, can recognize unmethylated cytosine-guanosine (CpG) motifs, which might be present in ODNs, and mediates proinflammatory effects [17–19]. These unmethylated CpG motifs, common in bacteria and DNA viruses, are mostly suppressed and methylated in vertebrate genomes. While some therapeutic ODNs incorporate CpG motifs to elicit therapeutically beneficial proinflammatory effects for a range of indications [20,21], most therapeutic ASOs have been optimized to minimize risk of proinflammatory response by methylating every cytosines in the ODN sequence, as well as avoiding the presence of CpG motifs.
Historically, the bulk of the screening experience with ASOs has been gained in rodents. Overall, rodents have been shown to be the most sensitive species, exhibiting a range of inflammatory features ranging from elevated chemokines, increased organ weight attributed to lymphohistiocytic infiltration and various inflammatory cell proliferation [22,23]. As a result of this rigorous screening process, the incidence of proinflammatory effects observed in the patients such as injection site reactions and constitutional symptoms has decreased, resulting in better tolerated ODN in a clinical setting.
In this study, we report the case of a 2′MOE ASO, ISIS 353512, which was developed originally to inhibit human C-reactive protein (hCRP) mRNA and, as a result, reduce the level of human CRP in the serum (hsCRP) to decrease the severity of cardiovascular diseases [24,25]. In preclinical studies, ISIS 353512 did not reveal any evidence of exaggerated proinflammatory effects in either rodent or nonhuman primate safety studies.
To assess the safety in humans, ISIS 353512 was administered subcutaneously initially to healthy human volunteers in a single and multiple ascending dose double-blind randomized trial (NCT00734240). Since ISIS 353512 was intended to reduce levels of hCRP mRNA, serum levels of hsCRP were closely monitored during the clinical trial. Unexpectedly, treatment with ISIS 353512 caused a higher degree of injection site inflammation, transient constitutional symptoms such as fever, chills, asthenia, feeling hot, and feeling cold, shortly after the first dose and increases in several proinflammatory endpoints such as interleukin (IL)-6 than expected. In particular, hsCRP was markedly elevated in the majority of volunteers treated with a single 100 and 200 mg dose of ISIS 353512. As a result, clinical investigations of ISIS 353512 were discontinued.
In this article, we present an in vitro method using human peripheral blood mononuclear cells (hPBMCs) to identify ODNs, such as ISIS 353512, capable of eliciting a human-specific proinflammatory response, and exclude them as a clinical development candidate, thereby minimizing the risk of unforeseen inflammatory-related adverse events in volunteers or patients. As with any recent ASOs tested in patients, ISIS 353512 did not contain any problematic motif such as proinflammatory CpG motifs and G quartets. Using hPBMCs, we demonstrated that, despite the lack of CpG motifs, ISIS 353512 is likely eliciting a human TLR9-mediated proinflammatory response. In hindsight, this outcome is not completely surprising as known differences between rodents and humans in term of the cellular distribution of TLRs and their propensity to recognize slightly different canonical CpG motifs in a species-dependent manner have been reported previously [26].
Materials and Methods
ODN design, synthesis, and preparation
All ODNs were designed and synthesized at Ionis Pharmaceuticals (Supplementary Table S1). To identify mouse ASO inhibitors, rapid throughput screens were performed in vitro as previously described [27]. The first 3 to 5 bases (5′ end) and last 3 to 5 bases (3′ end) of chimeric ASO have a 2′-O-(2-methoxy)-ethyl modification, and the ASOs also have a phosphorothioate backbone. ASOs were diluted in phosphate-buffered saline (PBS) for both in vivo and in vitro usage.
Meso Scale Discovery platform
Multiplex plates precoated with capture antibodies for specific cytokines, along with diluents, were brought to room temperature before use. Diluted standards (three replicates) and undiluted samples were added to the wells and incubated for 2 h in the Meso Scale Discovery® Custom array. All incubations were performed at room temperature with vigorous shaking (300–1,000 rpm). After the 2-h incubation, the plate was decanted and washed three thrice before adding a cocktail of Sulfo-Tag® detection antibodies to each well.
After incubating with detection antibodies for 2 h, the plate was again decanted and washed thrice. Read buffer was added to the wells using reverse pipetting techniques to ensure bubbles are not created to interfere with imaging. The plate was immediately imaged using the Meso Scale Discovery (MSD) Sector 2400 imaging system, and data were analyzed using MSD Discovery Workbench® software. Concentrations of all unknown samples were back-calculated using results interpolated from the corresponding standard curve regression using a weighted, four-parameter fit. Final sample concentrations (pg/mL) were calculated by factoring the dilution factor used for each sample.
Primate source and care
Male and female cynomolgus monkeys were obtained from Guangxi Grandforest scientific primate company, Ltd. (Guangxi, China). All animal procedures were conducted at the Korean Institute of Toxicology (Daejon, Korea) utilizing protocols and methods approved by the Institutional Animal Care and Use Committee (IACUC) and were in compliance with Animal Welfare Act and Guide for the Care and Use of Laboratory Animals (by ILAR publication).
Good laboratory practice toxicology and toxicokinetic study of ISIS 353512 in cynomolgus monkeys
Monkeys received either subcutaneous injection of saline (vehicle control) or ISIS 353512 at dose levels of 3, 8, 20, and 40 mg/kg/dose on days 1, 8, and 11 (loading doses), and then once weekly for a total duration of 13 weeks, with a 13-week recovery period (saline control, 20 and 40 mg/kg groups only). Serum samples were collected at predose, 2, 6, 24, and 48 h following the first dose (day 1), and last dose (day 85). Serum samples were analyzed for CRP concentration by an enzyme-linked immunosorbent assay (ELISA) method using IMUCLONE CRP (hs) ELISA kit (American Diagnostica, Stamford, CT). Serum samples were analyzed for cytokine and chemokine levels (IL-6, IL-10, MCP-, and MIP-1α) using Meso Scale Discovery platform.
Good laboratory practice toxicology and toxicokinetic study of ISIS 353512 in Sprague-Dawley rats
Sprague-Dawley rats received either a weekly intravenous injection of saline (vehicle control) or ISIS 353512 at dose levels of 1, 3, 10, and 40 mg/kg/dose on days 1, 3, and 7 (loading doses), and then once weekly for a total duration of 13 weeks. Serum samples were collected 2 days after the last dose of ISIS 353512 on day 91 and were analyzed for C-Reactive Protein (CRP) concentration by an ELISA method using IMUCLONE CRP (hs) ELISA kit (American Diagnostica) according to the manufacturer's instructions.
In vivo treatment in mice with ODN
C57BL/6 mice received an subcutaneous (SC) injection of PBS, ISIS 104838, or ISIS 353512 at 150 mg/kg. Blood samples were collected by tail bleed before dosing, and 6 and 24 h after subcutaneous dosing. Immediately before sacrifice, mice were terminated in compliance with Animal Welfare Act and Guide for the Care and Use of Laboratory Animals (by ILAR publication). Plasma samples were isolated from whole blood and analyzed for cytokine and chemokine levels (IL-6, IL-10, MCP-1, and MIP-1β) using Meso Scale Discovery platform.
293 Cell lines expressing the mouse or human TLR9 gene
The cell lines, 293-mTLR9 expressing mouse TLR9, 293XL-huTLR9, expressing human TLR9, and the control cell 293XL-null, were obtained from Invivogen (San Diego, CA). The cells were cultured according to the manufacturer's instructions. Cell culture supernatants were analyzed for cytokine and chemokine levels (IL-8, IL-10, IP-10, and MCP-1) using Meso Scale Discovery platform.
Human and cynomolgus PBMC assay
Whole blood was collected from human volunteer donors with informed consent in 8–10 BD Vacutainer CPT 8 mL tubes (BD Biosciences, San Jose, CA). The blood samples were mixed immediately before centrifugation by gently inverting tubes 8–10 times. The CPT tubes were centrifuged at room temperature in a horizontal (swing-out) rotor for 30 min at 1,500–1,800 relative centrifugal force with brake off. The PBMCs were retrieved at the interface between Ficoll and polymer gel layers and transferred to a sterile 50 mL conical tube, pooling up to 5 CPT tubes/50 mL conical tube/donor. The cells were washed twice with PBS (Ca++ and Mg++ free; Gibco) before another round of centrifugation. The supernatant was discarded without disturbing the cell pellets which were then resuspended in RPMI + 10% fetal bovine serum (FBS) + penicillin/streptomycin.
The cell density was estimated, and the density adjusted as to plate cells at 5 × 106/mL in 50 μL/well of 96-well, sterile, round-bottom, polypropylene plates for each ODN concentration tested for each donor. The ODNs 1:5 serial dilutions [2 × ] were prepared in medium (RPMI + 10% FBS + pen/strep) in a separate sterile, V-bottom, polypropylene plate starting with the highest concentration (400 μM) in the top row. To each well 50 μL/well of 2 × concentrated ODN diluted in RPMI + 10% FBS + penicillin/streptomycin was added. The plated cells (5 × 105/well) were incubated for 24 h at 37°C and 5% CO2.
The PBMC samples were treated with ODN shortly after isolation, typically within 3 h of isolation. Resting PBMCs overnight or freezing the samples typically resulted in a complete abrogation of the response to non-CpG ODNs. Plates were centrifuged at 330 g for 5 min before removing supernatants to a separate sterile polypropylene 96-well plate. Cell supernatants were stored at −70°C until processing for cytokine assay profiling. Cell culture supernatants were analyzed for cytokine and chemokine levels (eotaxin, eotaxin-3, GM-CSF, IL-1β, IL-2, IL-6, IL-8, IL-10, IL-12p70, IP-10, MCP-1, MCP-4, MDC, MIP-1β, TARC, and TNF-α) using Meso Scale Discovery platform.
Magnetic cell sorting of B cells
Human B cells were isolated from PBMCs collected from human volunteer donors by magnetic-activated cell sorting using the B cell Isolation Kit II for human cells from Miltenyi Biotec, Inc. (Auburn, CA) according to the manufacturer's instruction. Using this approach, B cells are left untouched and ready for subsequent analysis. Human B cells from PBMCs were labeled with B cell lineage-specific CD19 antibodies and removed by magnetic-activated cell sorting by using the Lineage Cell Depletion Kit for human cells from Miltenyi Biotec, Inc. according to the manufacturer's instruction. Using this approach, remaining cells (CD19−) are left untouched and ready for subsequent analysis. The success or the enrichment or depletion was confirmed by flow cytometry. Supernatants were analyzed for cytokine and chemokine levels (eotaxin, eotaxin-3, GM-CSF, IL-1β, IL-2, IL-6, IL-8, IL-10, IL-12p70, IP-10, MCP-1, MCP-4, MDC, MIP-1β, TARC, and TNF-α) using Meso Scale Discovery platform.
Statistical analysis
The data of in vivo studies (at least four animals per group) are expressed as the mean – standard deviation and were analyzed by ANOVA. When significance was obtained, multiple comparison analysis was conducted using Bonferroni as a post hoc test. In some instances, unpaired Student's t-test was utilized. All graphs and statistical analysis were obtained using JMP 15 (SAS Institute, Inc., Cary, NC).
Results
Evaluation of the subchronic tolerability of ISIS 353512 in nonhuman primates and rodents
To ensure that the risk of adverse events in human healthy volunteers or patients is minimized upon the initiation of the phase I clinical trial, ISIS 353512 was tested for tolerability in cynomolgus monkeys as part of a 3-month repeat dosing good laboratory practice (GLP) toxicology study. ISIS 353512 was administered weekly at 8 and 20 mg/kg/dose (intravenous bolus injection) and 40 mg/kg/dose (subcutaneous injection) for 85 days, which revealed no evidence of marked proinflammatory effects. There was neither acute nor sustained induction of serum CRP levels immediately after either a single or multiple dose (Fig. 1A). There were neither acute nor sustained significant alterations in inflammatory cytokines (IL-1β, IL-6, IL-10, IFN-γ, and/or TNF-α) or chemokines (MIP-1α and/or MIP-1β) measured, induced by ISIS 353512, except highly variable MCP-1 values at doses ≥20 mg/kg/week 2 to 6 h after injection (Fig. 1B).
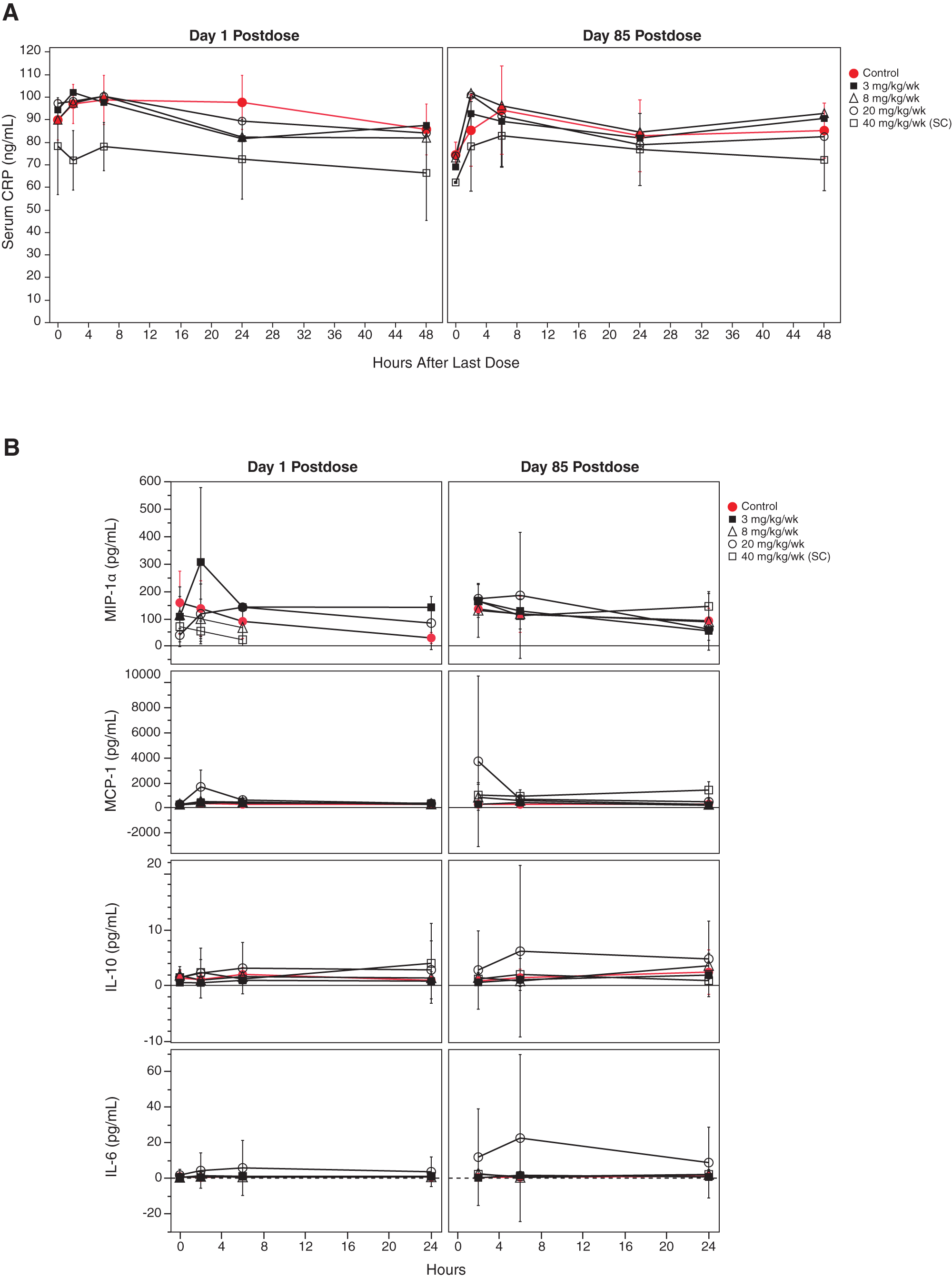
Cynomolgus monkeys were administered ISIS 353512 by IV dosing for control, and 3, 8, and 20 mg/kg/week groups and SC dosing for 40 mg/kg/week group. Serum samples were collected at 2, 6, 24, and 48 h after a single dose and on days 1 and 85 after the last dose.
As with cynomolgus monkeys, repeat administration of ISIS 353512 to Sprague-Dawley rats (GLP, 13 weeks, at up to 40 mg/kg weekly intravenous dosing) showed significantly increased spleen weight (P < 0.05, females: 1.6-fold, males: 1.7-fold at 3 mg/kg, Supplementary Fig. S1a). CRP level was also measured 7 days after the last dose and did not indicate a sustained induction of serum CRP levels (Supplementary Fig. S1b). The increased spleen the weight in absence of sustained CRP elevation noted at 3 mg/kg was interpreted then as nonadverse and provided a safety margin of ∼4-fold relative to the starting dose for the initial phase I clinical trial.
A follow-up study conducted in C57BL/6 compared cytokine and chemokine levels (IL-6, IL-10, MCP-1, and MIP-1β) induced at 6 and 24 h after a subcutaneous administration of either ISIS 353512 or ISIS 104838. ISIS 104838 is another MOE ASO that had been through toxicology and showed fewer constitutional symptoms and ISRs [28]. Overall, the level of induction by ISIS 353512 of various cytokine/chemokine levels was either lower or comparable to ISIS 104838 at 6 and 24 h, further suggesting that the proinflammatory potential in primates and rodents of ISIS 353512 was comparable to those of other ASOs such as ISIS 104838, which had previously advanced into clinical studies (Supplementary Fig. S2).
Therefore, combined findings from nonhuman primates and rodent studies indicated that administration of ISIS 353512 was tolerated well enough to proceed into human testing and would be unlikely to induce adverse proinflammatory responses in human volunteers.
Development of hPBMC assay to identify proinflammatory ODNs
The discrepancy between the mild proinflammatory signal in both nonclinical species and the constellation of proinflammatory signals such as elevated CRP or prevalent constitutional symptoms observed in ISIS 353512 phase I trial emphasized the need for an in vitro assay able to assess human-specific proinflammatory potential of novel ASOs to complement the preclinical toxicology studies.
Despite the lack of canonical CpG motif present on ISIS 353512, we hypothesized that the proinflammatory response observed in human volunteers might be, at least in part, mediated through TLR9 [15,17–19,29,30]. If indeed ISIS 353512 was able to activate TLR9 signaling, the expression of TLR9, which is mostly limited to B cells and plasmacytoid dendritic cells in humans [15,31,32], suggested that hPBMCs might provide a suitable model to assess the proinflammatory potential of ISIS 353512. Several authors [31,33,34] have previously shown hPBMCs to be an appropriate model to study the proinflammatory effect of CpG and non-CpG ODN.
PBMCs (5 × 105 cells/well) from five human volunteers were isolated and treated with either ISIS 104838 or ISIS 353512 within 2 h of collection with increasing concentrations of ASOs from 0.4 to 100 μM. The cells were incubated for 24 h, upon which time, the cell supernatant was collected and the levels for 17 cytokines and chemokines were measured using the MSD platform. Two distinct patterns emerged: IL-6 and IL-10, and to a lesser extent, TNF-α levels were clearly elevated in cells treated with ISIS 353512 relative to the other ASO tested (Fig. 2). While the magnitudes of the increase varied from donor to donor, the pattern remains consistent. IL-10 elevation was highest at the lowest dose tested (0.4 μM). IL-6 also showed a bell curve response to ISIS 353512, although less pronounced.

IL-6, IL-10, TNF-α, MIP-1β, MDC, and IP-10 levels following treatment of hPBMCs from five human volunteers treated with ISIS 104838 (open circles) and ISIS 353512 (closed circles) for 24 h. Cytokine and chemokine protein levels were measured using MSD. hPBMC, human peripheral blood mononuclear cell; MSD, Meso Scale Discovery.
In contrast, TNF-α displayed a more dose-responsive increase, but with less separation between cells treated with ISIS 353512 and ISIS 104838. The other pattern of response to ASO treatment to emerge is sequence independent but ASO treatment dependent elevations in chemokines MDC or MIP-1β. Other cytokines and chemokines analyzed showed no clear change (Supplementary Fig. S3). In follow-up studies, the focus of the analysis centered around IL-6, and to a lesser extentd IL-10 and TNF-α.
Next, we sought to investigate the range of responses of hPBMCs exposed to either ISIS 353512 or ISIS 104838 of a broader set of healthy human blood donors (n = 47). PBMCs from healthy volunteers were isolated and cells plated at 5 × 106 cells/mL and treated with increasing concentrations of ASOs for 24 h before harvesting the supernatant. Considering previous experiments conducted with a starting dose of 0.4 μM, which indicated that IL-6 and IL-10 productions were already at their maximum at this dose level, lower concentration range of MOE ASO from 0.019 to 80 μM was explored. The levels of IL-6, IL-10, and TNF-α were measured in the cell supernatant using MSD (Supplementary Fig. S4).
Overall, the hPBMCs from various donors exhibited a very broad range of IL-6 production in response to ISIS 104838 and ISIS 353512, and fell into two broad categories: discriminator and nondiscriminator donors. Depending on the donor, the IL-6 response of hPBMCs to ISIS 353512 peaks between 0.078 and 0.31 μM. To classify donors either as discriminators or nondiscriminators, the ratio between the IL-6 level produced in response to 0.31 μM of ISIS 353512 and the IL-6 produced in response to 0.31 μM of ISIS 104838 was calculated (Fig. 3A). Donors with an arbitrary ratio of 4 or greater were deemed to be discriminators (Fig. 3A). Based on this arbitrary threshold, hPBMCs isolated from 21 discriminator donors could very easily separate ISIS 353512 from ISIS 104838, with ISIS 353512 consistently producing the highest increase in IL-6 (Supplementary Fig. S4a and Fig. 3B).

In contrast, hPMBCs from other 26 donors (nondiscriminator) failed to show much separation between the two MOE ASOs. When IL-6 levels were normalized for each donor to the untreated level, donors classified as discriminators can produce a much greater level of IL-6 in response to ISIS 353512 than ISIS 104838 relative to untreated IL-6 levels (Fig. 3C). In contrast, hPBMCs from nondiscriminator donors tended to have a higher baseline production of IL-6 (Supplementary Fig. S5 and Fig. 3B), but much lower dynamic range of IL-6 production between ISIS 104838 and ISIS 353512 (Fig. 3C).
To further validate hPBMC ability to identify inflammatory ASOs, we selected three additional MOE ASOs, which had previously undergone phase I or phase II clinical trials {ISIS 104838 (NCT00048321) [28], ISIS 113715 (NCT00330330), ISIS 304801 (Volanesorsen) [35], and ISIS 325568 (NCT00519727) [36]} found to have comparatively milder constitutional symptoms than those induced by ISIS 353512 (data not shown). Although hsCRP or IL-6 was not among the endpoints collected during the clinical trials with those ASOs, a retrospective review of constitutional symptoms and other safety-related endpoints suggested that those MOE ASOs were better tolerated with respect to constitutional symptoms and ISR compared to ISIS 353512 (data not shown).
In addition to ISIS 104838 and ISIS 353512, those ASOs were used to treat hPBMCs from donors previously classified as discriminator and nondiscriminator. While discriminator donor hPBMCs treated with ISIS 353512 cells produced a significant increase in IL-6 (Fig. 3D) relative to ISIS 104838 at several of the ASO concentrations tested, the other ASOs (ISIS 113725, ISIS 304801, and ISIS 325568) demonstrated marginal increase in IL-6 levels relative to ISIS 104838, none of which was significant. In contrast, ISIS 353512 (or ISIS 113715, ISIS 304801, and ISIS 325568) treatment of nondiscriminator donor hPBMCs resulted in a similar degree of increase in IL-6 relative to ISIS 104838 at any of the ASO concentrations tested.
In contrast to IL-6, for which only treatment of hPBMCs from discriminator donors enable to identify ISIS 353512 as distinctly proinflammatory relative to ISIS 104838, IL-10 levels were also significantly elevated at lower doses of ISIS 353512 in hPBMCs from nondiscriminator donors (Supplementary Fig. S6). To a lesser extent, ISIS 113715 and ISIS 325568 also caused some significant increase in IL-10 levels relative to ISIS 104838, but at higher doses compared with ISIS 353512.
Reproducibility of donor-specific response
Some hPBMC (discriminator) donors have been tested many times on different days with ISIS 353512 and ISIS 104838. The PBMCs were collected and treated within 3 h of collection. In Fig. 4, we compare the IL-6 production in response to ISIS 104838 or ISIS 353512 stimulation of hPBMCs from six different donors in five independent runs of the assay. While the pattern of response remains fairly consistent over time for some donors (eg, #52), most donors display a broad range of response to ISIS 35512. However, in all runs and all donors, stimulation with ISIS 353512 always leads to a greater production of IL-6 than stimulation with ISIS 104838.
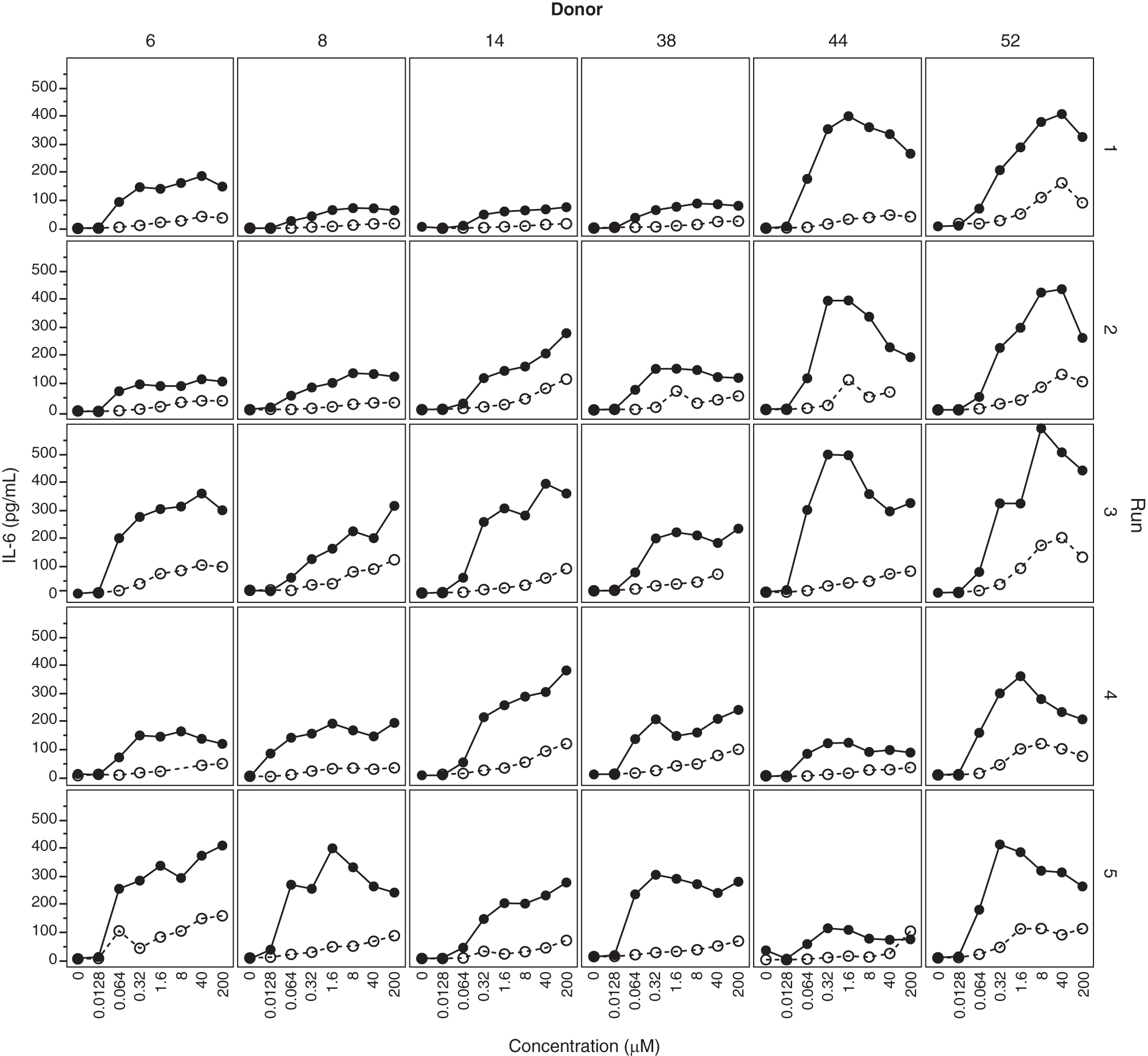
Reproducibility of IL-6 production in response to treatment to ISIS 104838 and ISIS 353512. PBMCs from six discriminator donors were treated with either ISIS 104838 (open circles) or ISIS 353512 (closed circles) on five different occasions.
PBMCs isolated from cynomolgus monkeys are unsensitive to ISIS 353512 stimulation
As stated previously, preclinical safety studies conducted in mice or non human primate did not identify the proinflammatory potential of ISIS 353512. Therefore, we sought to compare the effect of ISIS 353512 treatment in PBMCs isolated from either human volunteers or naive cynomolgus monkeys. NHP PBMCs were isolated and exposed to ISIS 353512 or ISIS 104838 under conditions similar to those employed for the hPBMC assessment. After 24 h of exposure, neither of the two MOE ASOs assayed induced an increase in any of the cytokine or chemokine measured at ASO concentrations overlapping those capable of eliciting an inflammatory response in hPBMCs (using a panel shown to be cross reactive to NHP) (Fig. 5A).
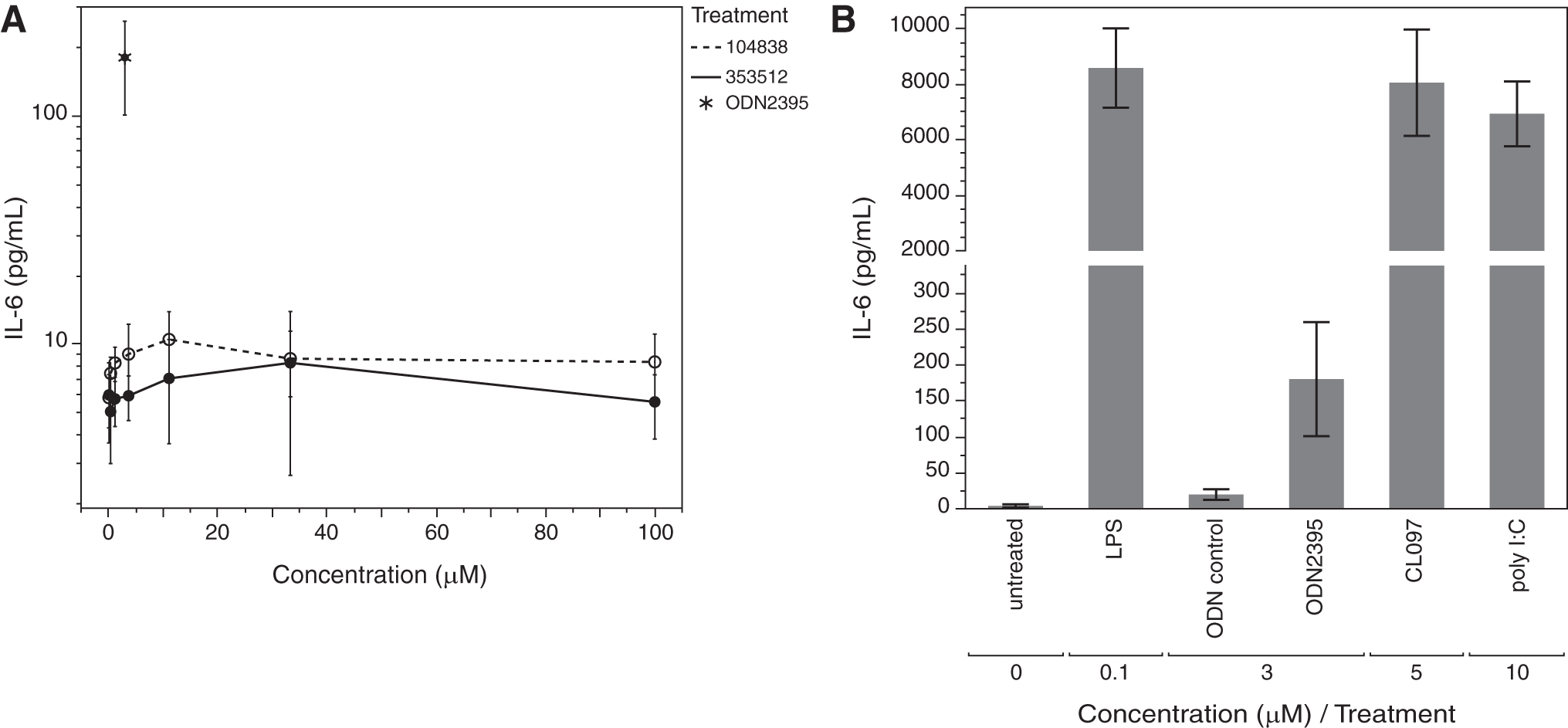
We sought to confirm that under the experimental conditions selected for the treatment, NHP PBMCs retained their sensitivity to several well-characterized proinflammatory stimuli. Responsiveness of monkey PBMCs to lipopolysaccharide, poly I:C, and a CpG ODN (ODN2395) [37] was demonstrated, thereby confirming that, while functional in their ability to respond to the range of TLR agonists, including TLR agonist (ODN2395), NHP PBMCs appeared to lack the ability to respond to more moderately proinflammatory MOE ASOs such as ISIS 353512, in contrast to hPBMCs (Fig. 5B). In addition, the magnitude of IL-6 production as measured using human anti-IL-6 detection confirms the cross-reactivity of the assay.
B cells are producing IL-6 in response to ISIS 353512
Next, we sought to identify which cell type(s) among the hPBMCs would be principally responsible for mediating the response to ISIS 353512. We had previously hypothesized that the most likely receptor mediating the inflammatory response would be TLR9 based on its recognition of CpG ODN [9].
In humans, two cell populations are mostly expressing TLR9: B cells and plasmacytoid dendritic cells. B cells are mostly responsive to CpG type B ODN, which has a structure most similar to 2′MOE ASOs, in that, they lack a PO central CpG-containing palindromic motif and a phosphorothioate-modified 3′ poly-G string present in CpG type A ODNs, to which plasmacytoid dendritic cells are more responsive [30,33]. To ascertain the involvement of B cells, we used a mixture of magnetically labeled antibodies directed against all the main cell types present in hPBMCs, except for B cells to negatively isolated B cells from the rest of the hPBMCs (Fig. 6 and Supplementary Fig. S7). We also conducted the reverse experiment in which we also used magnetically labeled antibodies against B cells to generate hPBMCs depleted of B cells isolated from two different donors.
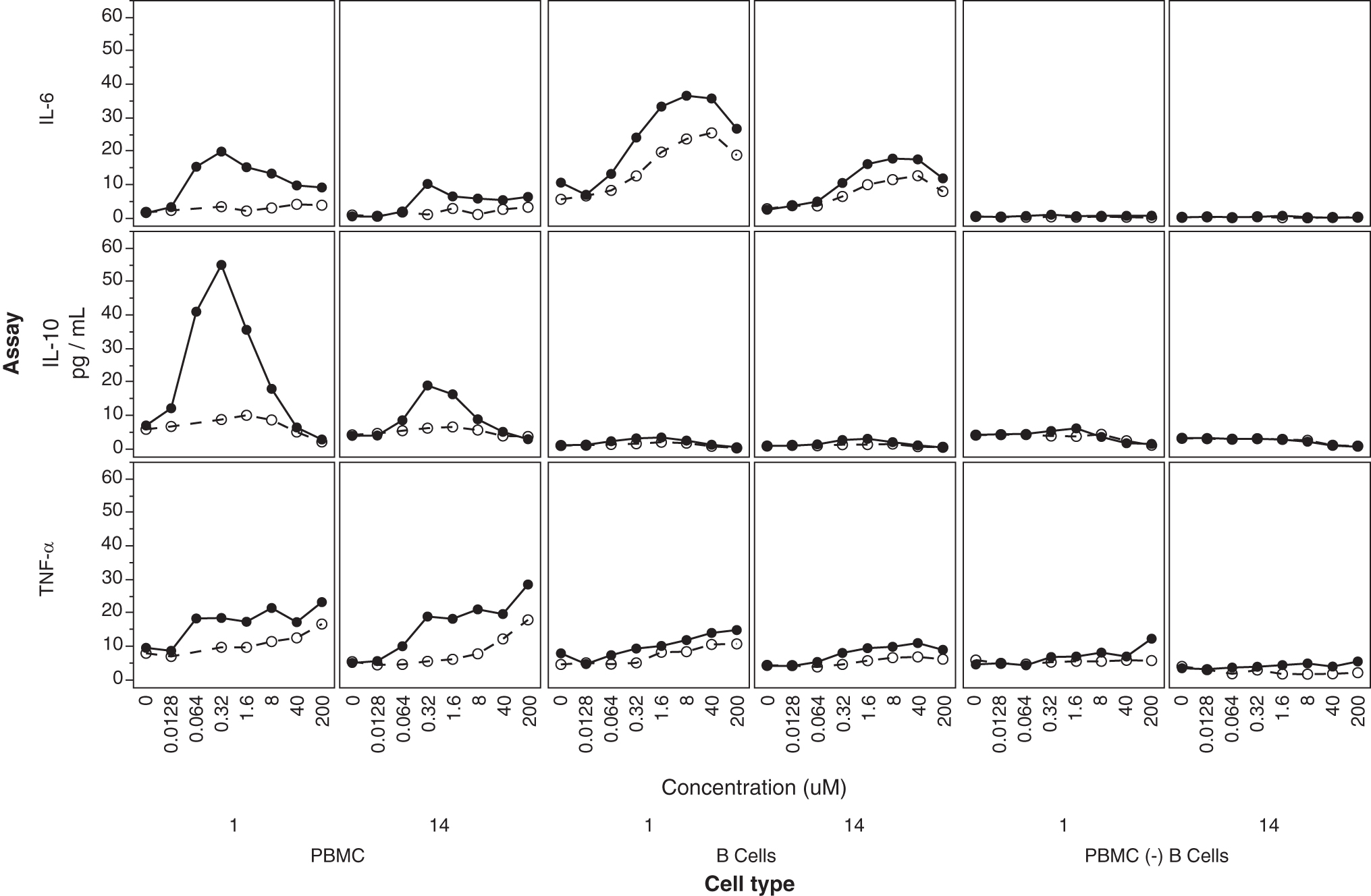
(Center) B cell (isolated from PBMCs—negative selection from donors 1 and 14) response to ISIS 353512 (closed circles) and ISIS 104838 (open circles) was compared to (left) hPBMCs or (right) hPBMCs depleted of B cells (positive selection of B cells). IL-6, IL-10s, and TNF-α were measured by MSD in the supernatant of the three cell populations following 24-h exposure to ISIS 3535312 for 24 h.
We then proceeded to treat the three different populations (isolated and unlabeled B cells, hPBMCs depleted of B cells, or unmodified hPBMCs) with either ISIS 353512 or ISIS 104838. Isolated B cells retained their ability to respond to ISIS 353512 through the production of IL-6 (Fig. 6). IL-6 production of isolated B cells in response to ISIS 104838 was somewhat increased relative to that of hPBMCs treated with ISIS 104838, but remained significantly lower than that to ISIS 353512. In contrast, B cells treated with either ISIS 353512 or ISIS 104838 did not display much increase in IL-10 production relative to untreated cells in contrast to hPBMCs treated with ISIS 353512, suggesting that the IL-10 production is a secondary event caused by the stimulation of other accessory cells [38].
Further emphasizing the contribution of B cells to the overall hPBMC response to inflammatory ASOs, hPBMCs depleted of B cells largely failed to show any increase in any of the cytokines assayed (Fig. 6). Other endpoints measured either did not show evidence of induction with either ASOs or did not discriminate between ISIS 353512 and ISIS 104838 response regardless of the cell type assessed (Supplementary Fig. S7).
B cell response to ISIS 353512 and ODN2006 is inhibited by chloroquine pretreatment
TLR9 signaling by proinflammatory CpG-containing ODNs requires internalization of TLR9/ODN complex [39–41]. Once internalized, TLR9 cleavage and activation by cathepsin, which requires endosomal acidification, take place. Suppression of endosomal TLR9 activation can be induced by inhibition of endosomal acidification using chloroquine, a weak base that accumulates in acidic compartments [42]. This requirement for internalization and endosomal acidification is shared by TLR3, TLR7, and TLR8, which also share with TLR9 an ability to respond to nucleic acid-based pathogen-associated molecular patterns (PAMP), in contrast to other TLRs (TLR1, TLR2, TLR4, TLR5, and TLR6), which do not recognize non-nucleic acid-based PAMP [such as LPS (TLR4) and flagellin (TLR5)].
To determine whether B cells were responding to ISIS 353512 stimulation through activation and signaling of one of the TLRs (TRL3, TLR7, TLR8, and TLR9) requiring endosomal acidification to enable TLR cleavage and signaling, hPBMCs were pretreated with 10 μM chloroquine, which resulted in the complete loss of the IL-6 production by ISIS 353512, ISIS 518477 [9], and ISIS 120704 (CpG, ODN 2006) (Fig. 7).
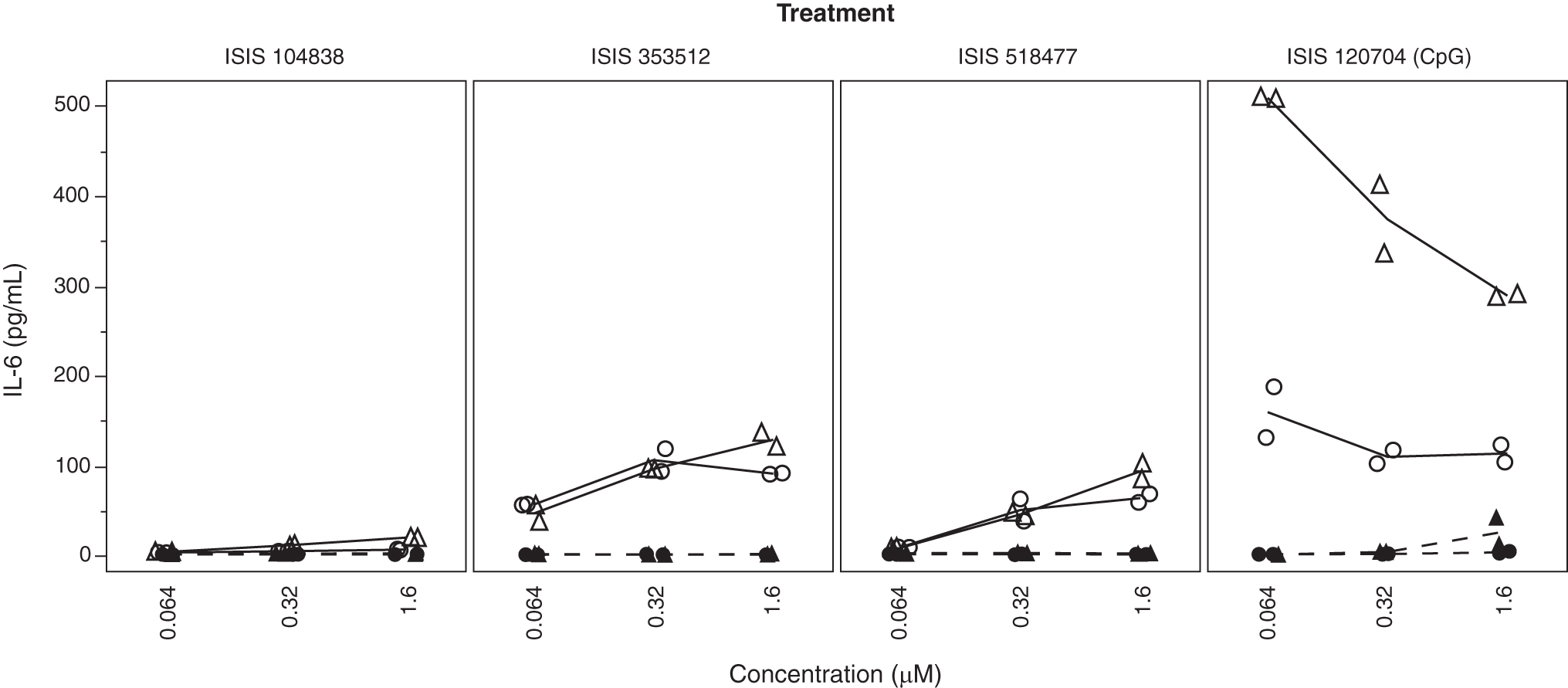
Responses of hPBMCs from two different human volunteers [44 (circle) and 52 (triangle)] to proinflammatory ODN stimulation were tested with absence (solid line and open symbols) or presence (dotted line and close symbols) of 10 mM chloroquine. TLR, toll-like receptor.
ISIS 120704 is a CpG-containing phosphorothioate ODN, while ISIS 518477 is a 2′MOE gapmer ASO, which does not contain any canonical CpG motif. Both have been previously shown to activate TLR9 signaling in rodents [9]. hPBMC response to LPS (TLR4), flagellin (TLR5), and other synthetic activators of non-nucleic acid PAMP-responsive TLRs, which does not depend on endosomal acidification as a prerequisite for activation, is essentially unaffected. In contrast, TLR3, TLR7, or TLR8 activation by their respective synthetic agonists (poly I:C, Imiquimod and ssRNA40) was abrogated in presence of chloroquine (Supplementary Fig. S8).
ISIS 353512 immunostimulation depends on the presence of unmodified bases
ISIS 353512 uses a “gapmer” design in which the sugars for 3 bases on the 3′ end and 3 bases on the 5′ end of the ODNs contain 2′MOE modifications. The remaining bases are natural DNA sugars. We hypothesized that, in part, the increase in proinflammatory signal was mediated by the size of the unmodified “gap.” Historically, a major improvement in the pharmacological and tolerability profile of ODNs was the addition of modified bases on the 5′ and 3′ end of the ODN. This resulted in increased stability and increased activity, as well as decreased proinflammatory properties relative to fully unmodified phosphorothioate ODNs [8]. Indeed, the increased number of modified 2′MOE bases and therefore the reduction in gap size from 14 bases (ISIS 353512) to 10 bases (ISIS 330012) resulted in a profound reduction in IL-6 and IL-10 production by hPBMCs from two donors (Supplementary Fig. S9).
293XL-huTLR9 cells overexpressing human TLR9 are insensitive to ISIS 353512 stimulation
Screening ODN for putative proinflammatory properties, hPBMCs come with a number of challenges: donor to donor variability and reproducibility, as well as limited or lack of insight into the mechanism of action of a given ODN. On the other hand, cell lines and in particular genetically modified cells, which overexpress a receptor such as TLR9 suspected of mediating an inflammatory response, promise to overcome many of those challenges. To that end, we tested ISIS 353512 and a number of reference ODNs in 293 cells overexpressing either mouse or human TLR9 (Supplementary Fig. S10).
The 293-mTLR9 cells overexpressing mouse TLR9 were the most sensitive to CpG type B (ODN2006) and CpG type C (ODN2395) ODN treatments, as well as a non-CpG ODN, ISIS 519488, also known to signal through mouse TLR9 [9]. The 293XL-huTLR9 cells overexpressing human TLR9 were mostly sensitive to CpG type B (ODN2006) and to a lesser extend CpG type C (ODN2395) ODN treatment. While both TLR9-overexpressing cell lines were responsive to CpG ODNs, treatment of either cell lines with ISIS 353512 failed to produce an induction of any of the cytokine/chemokine tested.
Discussion
During the pharmaceutical development process, establishing the safety or tolerability of a new pharmacological entity in patients is paramount. This relies on establishing a therapeutic index or margin: chiefly the ratio between the dose that would result in an unacceptable incidence of adverse events (whatever they maybe) and the dose that achieves the desired pharmacological effect designed to decrease the disease burden for the patient. One challenge in the nonclinical safety assessment is the translational relationship, or predictiveness of animals to humans. In most cases with ASOs transitioning into clinical trials, this approach has been a reliable mean of conducting the safety assessment.
ISIS 353512 was an exception as it had been shown to be well tolerated in both rodents and NHP 3-month tolerability studies at clinically relevant dose levels (Fig. 1), yet during the phase I/II human trials (NCT00734240), it became clear that the incidence of mild to moderate constitutional symptoms, along with increases in IL-6 and CRP at dose level required to demonstrate therapeutic efficacy, was unacceptably high. While the clinical findings were transient and did not result in severe injuries to the human volunteers, the discrepancy between the outcome of animal studies and those clinical findings demonstrated the necessity to include a human-based measure of inflammation to the nonclinical safety assessment process of ASOs. This is now a routine part of the compound selection and safety testing at Ionis Pharmaceuticals.
Past experience has suggested mice to be a sensitive species suitable to screen out nonspecific (TLR9 independent) proinflammatory effects of non-CpG ASO before the initiation of human clinical trials [8,11]. Screening for TLR9-mediated effects was commonly conducted in genetically modified cell lines overexpressing human TLR9 (Supplementary Fig. S10). Such cell lines are very sensitive to stimulation by TLR9 agonists such as canonical CpG ODNs, but were unable to identify ODNs such as ISIS 353512 as proinflammatory. Until the clinical trials with ISIS 353512, this approach resulted in multiple ASOs translating successfully into the clinic with acceptable proinflammatory profiles.
However, ISIS 353512 phase I findings revealed exaggerated proinflammatory phenotypes (volunteers treated with ISIS 353512 showed a higher incidence in mild to moderate constitutional symptoms such as fever, chills, asthenia, feeling hot, and feeling cold,… as well as injection site reactions, including injection site pain, erythema, itching, and swelling, and elevated IL-6 and CRP observed shortly after the first dose of ISIS 353512 relative to placebo-treated volunteers). Those findings were not predicted by either mice or monkey studies in which evidence of a proinflammatory response was minimal and similar to previous ASOs, which successfully entered clinical trials.
The nature of the clinical findings suggested a B cell-mediated response might be at play, while the nature of the stimuli, which is synthetic DNA like, suggested an innate immune receptor, likely TLR9, might be responsible for the proinflammatory response observed in the study. We and other authors had previously demonstrated that such a receptor, TLR9, mediated the inflammatory response to some ASOs in mice, despite the absence of any kind of CpG motif in those ASOs [9,33,43].
ODNs containing CpG motifs are known to elicit robust proinflammatory responses from rodent to humans [15,16,29]. Furthermore, TLR9-mediated DNA recognition and cell distribution have been shown to vary between species. In humans, TLR9 is predominantly expressed in B cells and plasmacytoid dendritic cells, while in rodents, TLR9 is expressed more widely, including myeloid immune cells. The optimal CpG motifs capable of activating TLR9 have also been shown to vary slightly from one species to the next [44,45]. However, ODNs lacking canonical CpG motifs, but with pyrimidine-rich immunostimulatory motifs such as TTTC motif present in the gap portion of ISIS 353512, have also been shown to be able to stimulate a potent immune response in PBMCs and primary B cells [33,43,46].
Therefore, we hypothesized that, despite the absence of canonical CpG motif, ISIS 353512 might be inducing a TLR9-mediated response involving primarily B cells and that the lack of ISIS 353512 proinflammatory stimulation in rodent may stem from species-specific recognition of CpG and non-CpG ODNs [47].
As with CpG ODNs exemplified by ODN2006 (ISIS 120704), treatment of PBMCs collected from five human volunteers with ISIS 353512 resulted in IL-6 increase 24 h after the initiation of the treatment in the majority of the PBMC samples tested (Figs. 2 and 7).
In contrast, this elevation in IL-6 was either minimal or not observed following exposure with ISIS 104838. The clinical experience with ISIS 104838 suggested that it was overall better tolerated in human volunteers than ISIS 353512 with a lower incidence or severity of flu-like symptoms [28]. As ISIS 353512, ISIS 104838 was found to be well tolerated in rodent and nonhuman primate studies. As such, ISIS 104838 was selected as being representative of ODNs with an acceptable tolerability profile in human volunteers. In parallel to testing those ODNs, in hPBMCs, we also tested them using whole blood collected from the same volunteers.
While treating whole blood samples with those ODNs also resulted in a differential increase in IL-6, which clearly discriminated ISIS 104838 from ISIS 353512 (data not shown), we settled on optimizing a tolerability screening assay using hPBMCs due to the ability to standardize the starting cell number to achieve greater consistency of response. Besides, IL-6, IL-10, TNF-α, or IL-8 were also differentially induced by ISIS 353512 relative to ISIS 104838. Other cytokines/chemokines such as MDC or eotaxin (Fig. 2 and Supplementary Fig. S3), similarly induced by either ODN, suggested another sequence-independent mechanism of inflammation might be at play as suggested by other authors [8,11,22]. In follow-up investigations, we sought to investigate the consistency of the response to ISIS 353512 across a larger number of hPBMC samples.
Surprisingly, the response of hPBMCs to ISIS 353512 could be assigned to two categories (Fig. 3 and Supplementary Fig. S4–S6): 21 donor samples demonstrated a good separation between ISIS 104838 and ISIS 353512 at a dose of 0.31 μM with an arbitrary ratio of IL-6 levels of 4 or more between the 2 ODNs (these donors were referred to as discriminators). In contrast, 26 donors were unable to discriminate those ODNs (with an ISIS 353512/ISIS 104838 IL-6 ratio below 4) (these donors were referred to as nondiscriminators). While the threshold is arbitrary, it is supported by the level of IL-6 production in the absence of treatment. Nineteen out of 21 discriminator donors had IL-6 level between 1 and 10 pg/mL after 24 h of incubation without ODNs (Supplementary Fig. S5). In contrast, under the same conditions, nondiscriminator donors exhibited IL-6 > 20 pg/mL in 23 out 26 cases.
The basis for the differential sensitivity of PBMCs from human volunteers to ISIS 353512 remains unclear and does not seem to translate to the clinical experience (although limited in scope) in which all volunteers treated with ISIS 353512 exhibited some increase in CRP. For ethical reasons, we were not able to ascertain whether the discrimination could be translated in vivo in a clinical setting. We were unable to test retrospectively the responsiveness of hPBMCs from volunteers enrolled in the clinical trials to further explore the connection between in vitro and in vivo response.
Regardless, the presence of two types of in vitro responses to ISIS 353512 highlighted the need for prescreening donors to identify samples with the optimal response profile to ISIS 353512 before screening novel ODNs. For donors classified as discriminators, while ISIS 353512 consistently led to a greater increase in IL-6 relative to ISIS 104838, the absolute level of IL-6 productions varied markedly from one run to the next (Fig. 4), which illustrated the need to include some reference compounds such as ISIS 104838 and ISIS 353512 to be able to determine whether the PBMCs from a given donor are behaving as required.
Typically, screening using hPBMCs to determine the likelihood of novel compounds to produce an adverse proinflammatory response is performed using at least two different (discriminator) donors. Interpreting the screening data to determine whether a novel compound should move forward largely relies on the relative ranking of compounds rather than an absolute pass or fail threshold. Any ODN that elicits an IL-6 response, which nears or exceeds that of ISIS 353512 in at least one donor, is typically eliminated. Similarly, any ODN that produces an increase in IL-6, which nears or is below that of ISIS 104838 in both donors, is deemed to have a very low risk of producing an adverse inflammatory response in patients. Typically, any ODN that exhibits a relative increase in IL-6 halfway between ISIS 104838 and ISIS 353512 is treated with caution.
As such, a follow-on MOE ASO to ISIS 353512, ISIS 329993, also designed to reduce levels of CRP mRNA (and protein), was administered to human volunteers in both single and multiple ascending dose double-blind randomized phase I clinical trial at a starting dose of 50 mg [48]. ISIS 329993 is targeting a different region of the CRP mRNA transcript and therefore has a distinct sequence compared to ISIS 353512, and has a 5-10-5 MOE gapmer configuration (Supplementary Table S1).
In contrast to ISIS 353512, treatment with ISIS 329993 did not cause an increase in hsCRP in human volunteers at doses up to 400 mg/week SC relative to placebo-treated volunteers. In addition, clinical observations of the volunteers treated with ISIS 329993 showed a marked reduction in incidence and severity of constitution symptoms such as fever, chills, aches, relative to ISIS 353512. ISIS 329993 was well tolerated at doses exceeding those tested with ISIS 353512 and progressed to phase II (NCT01710852 and NCT1414101). ISIS 329993 caused little to no increase in CRP level even at a dose of 600 mg. The minimal immune stimulation seen in the clinical trial was paralleled by the minimal induction in IL-6 production by hPBMCs treated with ISIS 329993 (Supplementary Fig. S11).
Besides ISIS 353512 and ISIS 329993 having different sequences, ISIS 353512 has 14 unmodified bases with phosphorothioate linkages against 10 for ISIS 329993 or ISIS 104838. When the number of unmodified bases was reduced from 14 (ISIS 353512) to 10 (ISIS 330012), IL-6 production by hPBMCs treated with ISIS 330012 was very similar to that of hPBMCs treated with ISIS 104838 (Supplementary Fig. S9), indicating that the number of unmodified bases with phosphorothioate linkages is an important contributor to the immune stimulation by ASOs. In recent years, most ASOs that have transitioned to a preclinical tolerability study in NHP and eventually to a clinical study have a “gap” of 10 unmodified bases or less with 3′ and 5′ “wings,” with modified bases that can vary in nature (mostly 2′MOE or cEt) [49].
As a result, one hundred thirty 5-10-5 MOE gapmer ASOs scheduled to be tested in NHP preclinical studies have been tested in the hPBMC assay before the start of those studies. IL-6 profiling from hPBMCs treated with those ASOs indicated that even with a constant number of unmodified bases, ASOs with difference sequence composition, but identical chemistry design can display a wide range of IL-6 induction, in some cases exceeding that of ISIS 353512 by more than one order of magnitude, despite the absence of canonical CpG motif in the sequence indicating that both the number of unmodified bases and the nature of the sequences are two fundamental modulators of the potential to trigger IL-6 stimulation (Supplementary Fig. S12).
We sought to gain a better understanding of the mechanism underpinning the production of IL-6 following treatment with ISIS 353512. As mentioned previously, we hypothesized that the mechanism might be, in part, overlapping with that of canonical CpG ODNs. Both human B cells and plasmacytoid dendritic cells express TLR9. Depending on the nature of the CpG motif, ODNs, the response will be either mostly driven by B cells or plasmacytoid dendritic cells [30,33]. Three types of stimulatory CpG ODNs have been characterized, type A, B, and C, which differ in their immune-stimulatory activities [17,30]. ODN2006 (ISIS 120704) is a 24-mer ODN with a full phosphorothioate backbone and belongs to the type B CpG ODNs, which contain one or more unmethylated CpG dinucleotides in particular sequence contexts (CpG motifs), and are recognized by human TLR9.
Type B CpG ODNs strongly activate B cells, but stimulate weak IFN-α secretion. Type A CpG ODNs are characterized by a phosphodiester central CpG-containing palindromic motif and a phosphorothioate 3′ poly-G string. They induce high IFN-α production from plasmacytoid dendritic cells. Type C CpG ODNs combine features of both types A and B. In this case, the structure of ISIS 353512 loosely resembles CpG type B such as ISIS 120704 [29,50], in that ISIS 353512 does not include a 3′ poly-G string characteristic of type A and C CpG ODNs. The ability of ISIS 353512 to induce an inflammatory response was compared to ISIS 120704 in nine different hPBMC donors (Supplementary Fig. S13a, b).
ISIS 120704 induced a maximum 189-fold increase in IL-6 production at 0.064 μM relative to untreated hPBMCs. In contrast, treatment with ISIS 353512 resulted in a maximum 37-fold increase in IL-6 production at 1.6 μM relative to untreated hPBMCs. Finally, treatment with ISIS 104838 induced a maximum of 11-fold increase in IL-6 production at 200 μM relative to untreated hPBMCs. Despite the inadequate safety profile of ISIS 353512, the potency of ISIS 353512 to induce an inflammatory response is much lower than a CpG ODN such as ISIS 120704.
Next, we sought to examine whether the cellular response to ISIS 353512 might be driven by B cells. The ability of hPBMCs and B cells to mount an IL-6 response to CpG ODN treatment has previously been demonstrated [51]. Indeed, depleting PBMC of B cells resulted in the complete abrogation of IL-6, IL-10, and TNF-α response in hPBMCs from two separate donors (Fig. 6). When B cells were isolated and treated with ISIS 353512, the ability to produce an IL-6 response to the treatment was retained. Interestingly, the IL-10 production and to a lesser extent TNF-α remained absent, suggesting that the IL-10 response observed in hPBMCs is a secondary event caused by the stimulation of other accessory cells, indicating an interaction between distinct cell populations in hPBMCs [38].
While treating hPBMCs with ISIS 353512 consistently produces more IL-6 than ISIS 104838 in all the donors tested, this is achieved under specific collection conditions. Typically, PBMCs are isolated from the blood of healthy volunteers and treated within a 3-h window. Attempts to streamline the assay by either resting PBMCs at 4°C overnight or freezing PBMCs obtained from buffy coats resulted in a systematic loss of IL-6 following stimulation with ISIS 353512 (data not shown).
This result was in contrast with the stimulation by a canonical CpG ODN such as ISIS 120704, for which resting or freezing PBMCs before the assay resulted in minimal loss of IL-6 induction (data not shown). The reason for this anergy could not be established. Despite the rapid anergy limiting our ability to fully dissect the mechanism, we gain some limited insight into the IL-6 production by ISIS 3512 in pretreating hPBMCs with chloroquine.
Chloroquine prevents endosomal acidification and the activating cleavage of TLR3, TLR7, TLR8, and LR9 [39,42,52]. Chloroquine treatment suppressed TLR3-, TLR7-, TLR8-, and TLR9-mediated inflammatory response by their respective canonical agonist (poly I:C, imiquimod, ssRNA-40, and ISIS 120704, respectively). The same was also true for ISIS 353512 and ISIS 518477, where chloroquine treatment of hPBMCs also substantially reduced the IL-6 production (Fig. 7). While we were unable to directly demonstrate that ISIS 353512 treatment leads to an inflammatory response, mediated through an interaction with TLR9, the circumstantial evidence (B cell responsiveness and mitigation of the inflammatory response by chloroquine) presented above strongly implicates a subset of the TLRs, and TLR9 in particular as the key mediator.
In conclusion, we describe a simple in vitro screening paradigm to supplement rodent and nonhuman primate toxicology studies, which enables the investigators to reliably identify ODNs susceptible of eliciting an adverse inflammatory response, specifically in humans before the initiation of clinical trials, thereby minimizing the risk to patients. This screening paradigm has a limited throughput due to its requirement for freshly collected hPBMCs. In addition, the donor-to-donor variability reduces the robustness of the assay. Nonetheless, the nature of the cell responsive to ASO stimulation (B cells) and the likely receptor (TLR9) open a path forward to identify a cell line-based alternative, which would replicate most features of the hPBMC assay, while increasing the throughput and consistency of the assay, thereby enabling users to implement the cell line assay at a much higher throughput.
The approach detailed herein also shows limitations in conducting effective structure activity relationship studies or mechanistic studies due to the variable nature of the PBMC response. A cell line meeting those criteria, Bjab, has been identified and is described in a follow-up article by Pollak et al. [53].
Footnotes
Acknowledgments
The authors thank the donors who participated in this study; Erin Morgan for her help in critical review of an earlier version of the article; Wanda Sullivan for assistance with formatting the figures; and Angela Colabucci and Tamika Holmes in proofreading and preparing the article for publication.
Author Disclosure Statement
The authors are either current or former employees of Ionis Pharmaceuticals.
Funding Information
No external funding was received for this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.