
Abstract
2′-O-Methoxyethyl antisense oligonucleotide (2′-MOE ASO)-induced severe thrombocytopenia (TCP) [platelet (PLT) count <50 K/μL] was observed in the Asian-sourced cynomolgus monkeys with low incidence (2%–4% at doses >5 mg/kg/week). The potential mechanisms for TCP were studied using the Mauritian-sourced cynomolgus monkeys, which were shown to be more susceptible to ASO-induced TCP, along with the Asian-sourced animals. ISIS 405879, a 2′-MOE ASO, induced severe TCP (PLT <50 K/μL) in seven of nine Mauritian-sourced monkeys but not in the Asian-sourced monkeys after 16 weeks of treatment at 40 mg/kg/week. Marked increases in PLT-bound C3d/C4d were detected in all thrombocytopenic Mauritian-sourced monkeys but not in the unaffected Mauritian- or Asian-sourced monkeys, suggesting increased PLT clearance due to complement deposition on the PLTs. However, this effect was independent of the ASO-mediated fluid-phase alternative complement activation. A correlation was also observed between serum antiglycoprotein (GP) IIb/IIIa immunoglobulin G (IgG) and PLT reduction. In addition, increases in total serum IgM, anti-PLT IgM, and anti-PLT factor 4 IgM levels were observed in monkeys from both sources but were more evident in the Mauritian-sourced monkeys. These data suggest an enhanced innate immune cell activation to ISIS 405879, leading to increased PLT destruction through complement fixation on the PLTs or PLT crossreacting polyclonal antibody production.
Introduction
Nonhuman primates are considered the most relevant animal species for the human safety assessment of the 2′-O-methoxyethyl antisense oligonucleotide (2′-MOE ASO) due to their comparable tolerability and pharmacokinetic properties to humans [1]. The nonclinical toxicology profiles of this class of ASOs have been well characterized previously [2]. Decreases in platelet (PLT) counts have been observed in Asian-sourced cynomolgus monkeys (eg, Cambodian, Vietnamese, and Chinese origins), given selected 2′-MOE ASOs [3]. However, the incidence and severity of PLT reductions vary with ASO sequences, studies, and animals. There are two phenotypes of PLT changes in monkeys; both are dose dependent and reversible [3].
Phenotype 1 is a consistent, reproducible, and self-limiting PLT decline occurring over ∼4–6 weeks of treatment, with ∼30% to 50% reduction from baseline, but mostly remains within the normal range with continued treatment (ie, PLT count ≥150 K/μL). This phenotype is not considered an adverse effect and occurs in ∼30% of the 2′-MOE ASOs evaluated in monkeys, based on a query from the Ionis Monkey Toxicology Database [3]. Phenotype 2 is sporadic and associated with more severe decreases in PLTs (ie, PLT count <50 K/μL) that can affect the overall hemostasis of the monkeys.
The incidence of this type of severe PLT reduction is ∼2% to 4% in Asian-sourced monkeys receiving 2′-MOE ASOs at doses >5 mg/kg/week [3]. Despite the low incidence of phenotype 2 in nonhuman primates, it did translate to a clinical issue in the Tegsedi (TTR) and Waylivra (ApoCIII) development programs at Ionis [3–5]. Nonetheless, contributing factors from the underlying disease conditions cannot be ruled out completely as a cause in the clinical issue [4,6].
The mechanism of phenotype 1 PLT reduction was investigated previously using 111Indium (In)-labeled PLTs from Asian-sourced cynomolgus monkeys treated with ISIS 104838 [7]. ISIS 104838 is a prototypical 2′-MOE ASO that reproducibly causes the consistent, moderate, phenotype 1 PLT reductions in monkeys. An increase in 111In-labeled PLT sequestration in the liver and spleen was observed after 40 mg/kg/week ISIS 104838 treatment in monkeys for 12 weeks.
In addition, evidence of monocyte activation with increases in innate immunoglobulin M (IgM) responses [eg, total serum IgM, anti-PLT IgM, and/or anti-PLT factor 4 (PF4) IgM] was noted, likely leading to the increased PLT sequestration in peripheral tissues and the PLT count reduction in circulation [7].
In contrast, studying the mechanism for the sporadic, severe, phenotype 2 PLT reduction has been challenging due to the low incidence in the Asian-sourced cynomolgus monkeys and the inconsistent nature of such effect. Therefore, a more sensitive source of cynomolgus monkeys is needed to help understand the mechanism(s) behind the phenotype 2 PLT reduction.
In this study, Asian-sourced (Cambodian origin) or Mauritian-sourced cynomolgus monkeys were treated with ISIS 405879 at 40 mg/kg/week for 16 weeks. The primary parameters evaluated included clinical pathology [hematology, coagulation, and serum chemistry, including complement component 3 (C3), and immunoglobulin (IgG and IgM)]. Additional assessments included PLT-bound IgG and IgM, PLT-bound C3d and C4d, serum biomarkers for anti-PLT IgG and IgM, epitope-specific anti-PLT IgG [eg, glycoprotein (GP) IIb/IIIa, GPIa/IIa, GPIb/IX, GPIV, and Class I HLA-specific monoclonal antibodies], anti-PF4 IgG and IgM, cytokines/chemokines, fluid-phase complement split products in the plasma (Bb, C3a, and C4a), in vitro and in vivo PLT activation assay, PLT microparticles, and leukocyte-PLT aggregation (whole blood).
Materials and Methods
This study and related analyses were not conducted in compliance with the Food and Drug Administration Good Laboratory Practice (GLP) regulations for Nonclinical Laboratory Studies; however, applicable standard operating procedures and sound scientific practices were followed.
Chemical and reagents
ISIS 405879 is a 20-nucleotide PS ASO with the sequence
Animals
The study was conducted at Altasciences Preclinical Seattle LLC (Everett, WA, USA), an Association for Assessment and Accreditation of Laboratory Animal Care accredited facility that has an animal welfare assurance issued by the Office of Laboratory Animal Welfare, is registered with the U.S. Department of Agriculture, and has an Institutional Animal Care and Use Committee (IACUC) that is responsible for compliance with applicable laws and regulations regarding the use of laboratory animals.
Twenty-four Asian-sourced (Cambodian origin supplied by Orient BioResource Center) or Mauritian-sourced (Noveprim Ltd., Mauritius) cynomolgus monkeys were individually housed in a temperature- and humidity-controlled environment, providing a 12-h light/dark cycle. Animals were fed twice daily with ad libitum access to water. The study protocol was approved by IACUC at the Testing Facility before dose administration.
Study design
A total of 12 male and 12 female cynomolgus monkeys [6/sex Mauritian- and 6/sex Asian-sourced (Cambodian origin)] were used in the study (Table 1). Doses were administered subcutaneously (SC) for a total of 16 weeks. Monkeys were dosed on Days 1 (1st day of dosing), 4, 7, and then weekly thereafter on Days 14, 21, 28, 35, 42, 49, 56, 63, 70, 77, 84, 91, 98, 105, and 112 (total of 18 doses). Monkeys were evaluated daily for abnormal clinical signs. Body weights and qualitative food consumption were recorded weekly.
Study Design for the 16-Week Investigative Study with ISIS 405879 Using Asian (Cambodian Origin)- and Mauritian-Sourced Monkeys
Monkeys were dosed on Days 1 (1st day of dosing), 4, 7, and then weekly thereafter on Days 14, 21, 28, 35, 42, 49, 56, 63, 70, 77, 84, 91, 98, 105, and 112 (total of 18 doses). The total dose volume (mL) was calculated based on the most recent body weight. Terminal sacrifice was performed on Day 114 (∼48 h after the final dose on Day 112).
SC, subcutaneously.
The parameters evaluated at the Testing Facility included clinical observations, injection site observations, body weights, clinical pathology (hematology, coagulation, and serum chemistry, including C3, IgG, and IgM), PLT-bound IgG and IgM, C3d, and C4d, in vitro and in vivo PLT activation assays by flow cytometry. Additional assessments, including cytokines/chemokines, fluid-phase complement split products in the plasma (Bb, C3a, and C4a), anti-PLT and anti-PF4 antibodies (serum biomarker), were conducted at various contract laboratories (indicated in the sections below).
At the scheduled (Day 114) and unscheduled necropsies, gross observations were performed, and specific tissues/organs were collected (eg, liver, kidney, heart, spleen, lymph node, and bone marrow), fixed in 10% neutral buffered formalin, embedded in paraffin, sectioned, and stained with hematoxylin and eosin for examination by light microscopy.
In addition to any necessary standard veterinary medication, a corticosteroid (methylprednisolone acetate, Depo-Medrol®) treatment and/or dose suspension were used in monkeys when critically low PLT count (<2.5 K/μL) was confirmed (Table 2).
Veterinary Treatments and the Fate of the Animals Developed Thrombocytopenia in the 16-Week Investigative Monkey Study with ISIS 405879
In addition to any necessary standard veterinary medication, Depo-Medrol treatment and/or dose suspension were given to selected monkeys with severely low PLT counts (<2.5 K/μL). One course of Depo-Medrol treatment includes a 1st dose of 80 mg by intramuscular injection followed by a 2nd dose of 40 mg one week later.
PLT, platelet.
PLT-bound IgG, IgM, C3d, and C4d assay
The PLT-bound IgG, IgM, C3d, and C4d assays were conducted by Altasciences Preclinical Seattle LLC. Antibodies against CD41, C3d, C4d, IgG, and IgM were used to stain PLT-rich plasma (PRP) from either ISIS 405879-treated or control animals collected on Days -2 (baseline), 16, 44, 58, 72, 86, 100, and 114. Assessment of the relative percentage of positive cells was measured using flow cytometry. All reagents were prepared and used at concentrations recommended by the supplier or manufacturer and as established at Altasciences.
Positive control for IgM staining was performed on frozen cynomolgus peripheral blood mononuclear cells with antibodies against CD20 and IgM. Positive control for IgG staining was performed on Quantum Simply Cellular antimouse IgG beads from Bangs Laboratories (Fishers, IN, USA; Catalog No. 815) with an antibody against IgG. Positive control for C3d and C4d was performed on PRP incubated with heat-activated gamma globulin from Quidel (San Diego, CA, USA; Catalog No. A114) and stained with antibodies against C3d and C4d.
In vitro and in vivo PLT activation assays
The in vitro PLT activation assay was performed at Altasciences Preclinical Seattle LLC. Whole blood samples from three naïve Mauritian- and Asian-sourced (Cambodian origin) monkeys were centrifuged at 170 g for 15 min without braking. PRP was collected from the top layer and used for PLT activation experiments. PLTs were resuspended in modified HEPES-Tyrode buffer (10 mM HEPES, 137 mM sodium chloride [NaCl], 2.8 mM potassium chloride, 1 mM magnesium chloride, 12 mM sodium bicarbonate, 0.4 mM disodium phosphate, 0.35% [w/v] BSA, 5.5 mM glucose, pH 7.4) in a volume equal to original PRP volume.
The control and test articles included various concentrations of ISIS 405879 (1.6, 8, 40, and 200 μM), ISIS 120704 (200 μM), and adenosine diphosphate (ADP) (20 μM, as positive control). ISIS 120704 is a CpG containing non-MOE ASO that has been shown to cause PLT activation. The control or test articles were added to 20 μL whole blood in a final volume of 25 μL and incubated for 20 min at room temperature (RT).
After incubation, anti-CD41a-APC (for identification of PLTs) and anti-CD62P-PE (for assessment of activation), diluted in modified HEPES-Tyrode buffer, were added, each at 10 mg/mL final concentration, and incubated for 20 min at RT. Lyse/fixative solution (400 μL; 2% formalin, 10 mM HEPES, 0.15 mM NaCl, pH 7.4) was added to each sample, and samples were stored at 4°C until analyzed by flow cytometry (FACSCalibur™; Beckton-Dickinson). PLTs were identified by gating on CD41a+ and excluding events with low forward scatter to remove PLT-derived extracellular vesicles. Activated PLTs were identified by an increase in median fluorescence intensity (MFI) of CD62P compared with resting PLTs. Data were analyzed using FlowJo software (v.7.3; Treestar, Ashland, OR, USA).
The in vivo PLT activation assay was conducted as part of the PLT-bound IgG, IgM, C3d, and C4d assay by measuring the percentage of CD62P-positive PLTs and using PRPs collected from either saline or ISIS 405879-treated monkeys at various time points throughout the study.
Flow cytometry assay for detection of PLT drug-dependent antibodies
The drug-dependent antibodies assay was conducted at Versiti–Wisconsin, Inc. (formerly Blood Research Institute Blood Center Wisconsin, Milwaukee, WI, USA). In brief, serum samples were tested for drug-dependent and nondrug-dependent PLT antibodies by flow cytometry based on the methods published elsewhere [8]. Fresh human blood group O PLTs (1 × 107/well) from a single random blood donor were incubated with 40 μL of monkey sera and 25 μL of either ISIS 405879 (0.5 mg/mL) or PBS. PLTs incubated with serum and ISIS 405879 were then washed with buffer containing drug (0.5 mg/mL ISIS 405879).
PLTs incubated with serum and PBS were then washed in PBS alone. PLT-bound Igs were then detected with fluorescent-labeled antihuman IgG and antihuman IgM. PLT-bound IgG or IgM fluorescence intensity was determined by flow cytometry. Fluorescence ratio values were calculated for each sample by dividing MFI in the presence of the drug by MFI without the drug. This assay utilized human PLTs for testing cynomolgus monkey serum samples because of logistical issues involved in obtaining fresh monkey PLTs.
Only nondrug-dependent PLT antibody data were included in this article as there was no evidence of drug-dependent PLT antibody present in monkeys with 2′-MOE ASO treatment. This assay was not validated for use in testing nonhuman primate serum samples.
PLT antibody bead array assay
The PLT antibody bead array assay was conducted at Versiti–Wisconsin, Inc. Luminex polystyrene microspheres with different distinct emission spectra were coated with GPIIb/IIIa, GPIa/IIa, GPIb/IX, GPIV, and Class I HLA-specific monoclonal antibodies. Fresh human group O PLTs incubated with monkey serum were lysed, and the lysates were incubated with a mixture of the antibody-coated beads to specifically capture antigen–antibody complexes. The beads were then washed and incubated with phycoerythrin-labeled antihuman IgG. Fluorescence analysis of beads was performed on a Luminex instrument [9]. This assay was not validated for use in testing nonhuman primate serum samples.
Heparin-PF4 ELISA for detection of heparin-dependent antibodies
The PF4 ELISA was conducted at Versiti–Wisconsin, Inc., based on a previously described proprietary kit that detects antibodies of three Ig classes (IgG/A/M) against PF4/polyvinyl sulfonate (PVS) complexes [10]. A confirmatory high-concentration heparin step was performed concurrently, in which 100 U/mL (high dose) of unfractionated porcine heparin was added to the patient's sample. Monkey serum samples were diluted to 1:50 in PBS (pH 7.4) before testing.
In brief, monkey sera were incubated in duplicate with complexes of PVS and PF4 (PVS-PF4) in two separate wells of a microtiter plate. It is well established that PVS is equivalent to unfractionated heparin (UFH) when complexed with PF4 to detect heparin-dependent anti-PF4 antibodies in human sera [11]. UFH of 100 U/mL was added to the second well as a confirmatory step. Each well was washed and incubated with alkaline phosphatase-labeled antihuman IgG, IgA, or IgM.
Each well was washed again, and the bound antibody was detected by addition of phosphatase substrate. The optical density (OD) was measured by spectrophotometry. Sera from patients with heparin-induced thrombocytopenia (HIT) with IgG OD values >1.0 were used as positive controls. Only anti-PF4-IgG and anti-PF4-IgM were measured in this study. This assay was not validated for use in testing nonhuman primate serum samples.
Measurement of serum biomarkers
Multiplex cytokine analysis was performed on serum samples using a panel with known crossreactivity with cynomolgus monkeys, Milliplex panel kits (EMD Millipore, Billerica, MA, USA), and a Luminex FlexMAP 3D instrument with the XPonent version 4.0 software.
Thawed serum samples were assessed for the following cytokines/chemokines: granulocyte macrophage-colony stimulating factor (GM-CSF), interferon gamma (IFN-γ), interleukin 1β (IL-1β), IL-4, IL-5, IL-6, IL-8, IL-10, IL-12p40, IL-13, IL-17A, IL-21, IL-22, macrophage inflammatory protein 1-alpha, monocyte chemoattractant protein-1 (MCP-1), tumor necrosis factor-α (TNF-α).
von Willebrand factor (vWF) antigen was detected using an ELISA developed at Versiti–Wisconsin, Inc., using captured antibodies developed in their hybridoma laboratory and polyclonal antibodies from the appropriate vendor.
Complement Bb, C3a, and C4a assay
Fluid phase plasma complement Bb, C3a, and C4a concentrations were analyzed using the human ELISA assay kits from Quidel.
Statistics
Multiple comparisons were evaluated with one-way analysis of variance, followed by a post hoc Dunnett's test with JMP version 13 (SAS Institute). A P value <0.05 was considered significant.
Results
Ionis historical observation of 2′-MOE ASO-induced TCP in Mauritian-sourced monkeys
ISIS 405879, a 2′-MOE ASO developed by Ionis to treat hypercholesterolemia, demonstrated good tolerability in the 13-week GLP monkey study using the Asian-sourced (Chinese origin) cynomolgus monkeys. No incidence of TCP (PLT <50 K/μL) was observed at doses up to 40 mg/kg/week. However, in a subsequent chronic study using the Mauritian-sourced monkeys (Noveprim Ltd.), 5 of the 14 high-dose group animals (24 mg/kg/week) developed TCP with PLT nadir <50 K/μL after 3–6 months of treatment (Table 3). The study was terminated at 6 months due to the PLT finding.
Incidence of Thrombocytopenia from the 13-Week and 6-Month GLP Repeat-Dose Toxicity Studies with ISIS 405879 Using Asian (Chinese Origin)- or Mauritian-Sourced Monkeys
All five monkeys who developed TCP in the 6-month study were from the 24 mg/kg/week group; there was a total of 14 monkeys in the high-dose group.
GLP, Good Laboratory Practice; TCP, thrombocytopenia.
The differential effect on the PLT counts with ISIS 405879 treatment suggested a different sensitivity in the Mauritian-sourced monkeys compared with the Asian-sourced monkeys. However, the longer duration of treatment may also have played a role in the higher incidence of TCP. Based on this observation, the Mauritian-sourced monkey (Noveprim Ltd.) was used as a more susceptible model to study the potential mechanisms of ASO-induced TCP along with the Asian-sourced monkeys.
A higher incidence of 2′-MOE ASO-induced TCP was confirmed in Mauritian-sourced monkeys
In this study, Asian-sourced monkeys (Cambodian origin) and Mauritian-sourced (Noveprim Ltd.) monkeys were treated with a high dose of 40 mg/kg/week ISIS 405879 for 16 weeks. There were no changes in PLT count after the first dose, but severe TCP (PLT <50 K/μL) developed in Mauritian-sourced monkeys but not in the Asian-sourced (Cambodian origin) monkeys after repeat treatment (Fig. 1A, B). Of the nine Mauritian-sourced monkeys, seven developed severe TCP with PLT nadir <50 K/μL as early as after 6 weeks of treatment. Three Mauritian-sourced monkeys were terminated before the scheduled necropsy (Monkey 4501 on Day 56, Monkey 4001 on Day 68, and Monkey 4003 on Day 108) due to low PLT counts.
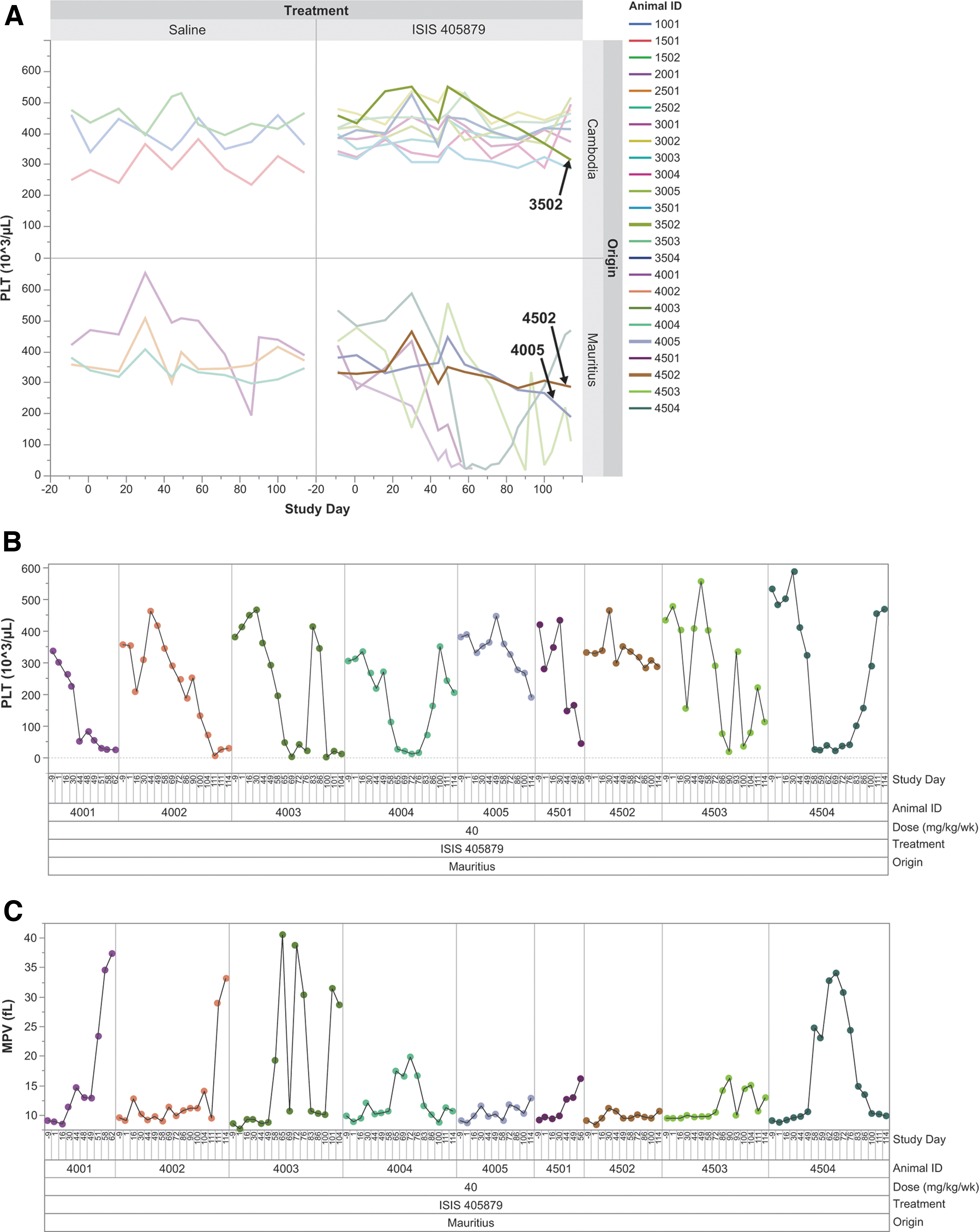
Severe TCP was observed in Mauritian-sourced monkeys but not in the Asian-sourced (Cambodian origin) monkeys after 16 weeks of treatment with ISIS 405879 at 40 mg/kg/week. Time-course changes of PLT counts for all groups at the scheduled and unscheduled time points
Two of the nine Mauritian-sourced monkeys (Monkeys 4005 and 4502) maintained their PLT counts >200 K/μL after 16 weeks of treatment, although a ∼30% reduction in PLT count from baseline was also observed in Monkey 4005 at the end of the treatment period (Day 114). The PLT reduction was reversible, after cessation of treatment alone or in combination with corticosteroid treatment. Monkey 4003 showed rapid recovery on PLT counts one week after the steroid treatment on Days 72 and 79. However, the PLT counts dropped to <50 K/μL again after reinitiating the ASO treatment on Day 91. Dosing was suspended again on Day 105 for Monkey 4003 before the unscheduled necropsy on Day 108.
Slower recovery in PLT counts (up to ∼5 weeks) was noted in monkeys with dose suspension only (Monkeys 4004, 4503, and 4504). Dosing resumed on Day 100 for all three monkeys, and the PLT counts reduced slightly again at the time of necropsy on Day 114 except for Monkey 4504 (Fig. 1B). The PLT counts for the Asian-sourced (Cambodian origin) monkeys remained in the normal ranges, with a 30% reduction from the baseline in one Asian-sourced (Cambodian origin) monkey (Monkey 3502), although the PLT nadir was still >300 K/μL.
All thrombocytopenic Mauritian-sourced monkeys showed increases in mean PLT volume (MPV) with peak value corresponding with PLT count nadirs in those monkeys (Fig. 1C). In addition, immature, large PLTs were noted in the blood smear from those thrombocytopenic monkeys, correlated with the MPV changes (Fig. 2A). Microscopic evaluation showed minimal-to-moderate megakaryocyte hyperplasia in the bone marrow from five of the seven thrombocytopenic Mauritian-sourced monkeys treated with ISIS 405879 (4001, 4002, 4003, 4501, and 4504), but not in the two unaffected Mauritian-sourced monkeys (4005 and 4502) (Fig. 2B and Table 4).
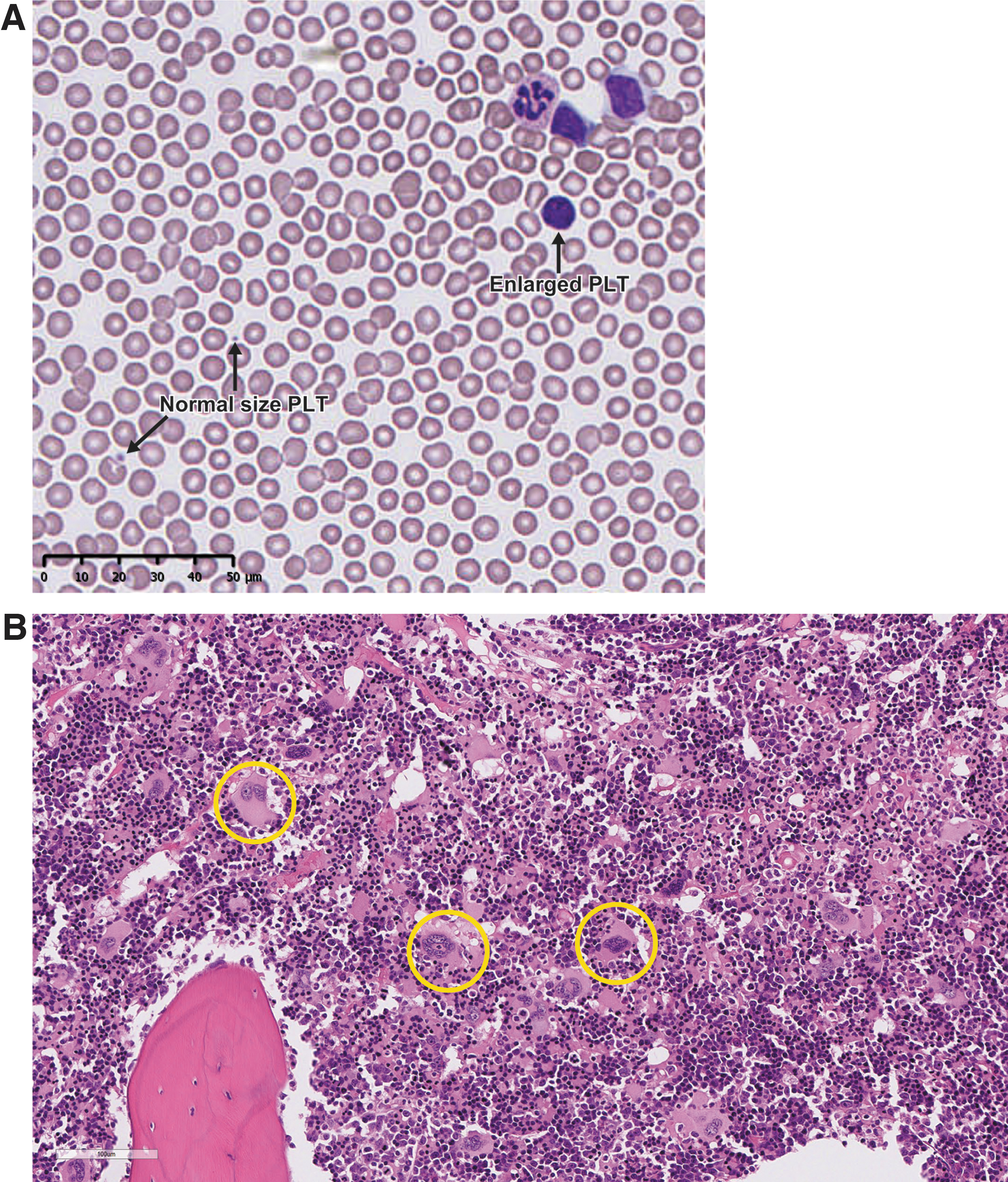
Enlarged immature PLTs were observed in the blood smear from the thrombocytopenic Mauritian-sourced monkeys and correlated with bone marrow megakaryocyte hyperplasia microscopically. Representative blood smear from Mauritian-sourced Monkey 4001 on Day 55 with a PLT count at 40 K/μL
Summary of Microscopic Findings in Selected Tissues from the 16-Week Investigative Monkey Study with ISIS 405879
Severity: minimal, slight, moderate, marked, and severe.
“—”: histologically not remarkable.
PTEC, proximal tubule epithelial cell.
There were also three ISIS 405879-treated Asian-sourced (Cambodian origin) monkeys that had megakaryocyte hyperplasia (slight in severity) at the terminal necropsy (Day 114), including Monkey 3502, who had a 30% reduction from baseline and two other monkeys (Monkeys 3004 and 3005) with no PLT change throughout the 16-week treatment period. Increased MPV and megakaryocyte hyperplasia in those thrombocytopenic Mauritian-sourced monkeys suggested a normal bone marrow response to a profound PLT reduction.
One of the thrombocytopenic Mauritian-sourced monkeys (Monkey 4501) also had increased cellularity of splenic red pulp that might be reflective of increased PLT removal from the circulation. In addition to bone marrow and spleen, microscopic evaluation was conducted in selected organs and tissues, such as liver, kidney, heart, and lymph nodes (Table 4). All other lesions present were either spontaneous background alterations or of the spectrum commonly seen in nonhuman primates treated with ASOs (e.g., findings related to ASO uptake and tissue accumulation).
Increased complement C3d/C4d deposition on PLTs was observed in all thrombocytopenic Mauritian-sourced monkeys, suggesting complement fixation on PLT surface
Deposition of complement split products C3d and C4d was noted on PLTs (CD41+ C3d+% and CD41+ C4d+%) in the ISIS 405879-treated thrombocytopenic Mauritian-sourced monkeys, with the peak values generally correlated with the PLT nadirs for those monkeys (Fig. 3A, B). No apparent increases in C3d and C4d deposition on the PLTs were noted in the unaffected Mauritian- (Monkeys 4005 and 4502) or Asian-sourced (Cambodian origin) monkeys. Increases in the percentage of PLT-bound C3d and C4d generally correlated with a reduction in PLT counts with R2 at 0.571 and 0.452, respectively (Fig. 3C).
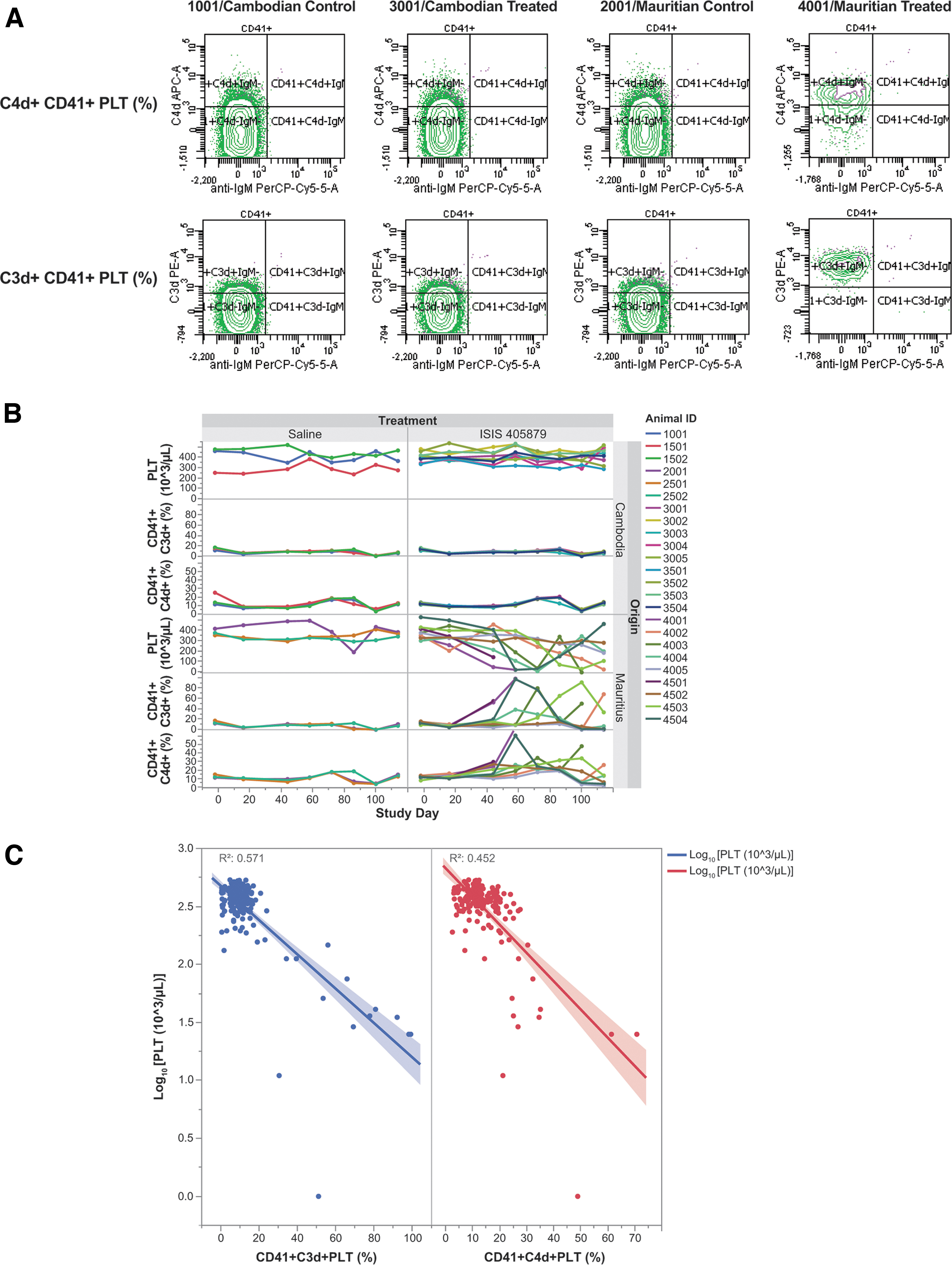
Increased C3d and C4d deposition on PLTs in the thrombocytopenic Mauritian-sourced monkeys. PRPs were collected at the scheduled time points, and measured for PLT-bound C3d and C4d levels using flow cytometry. Representative cytograms of PLT-bound C3d/C4d on Day 58 from control and ISIS 405879-treated Asian (Cambodian origin)- and Mauritian-sourced monkeys
The PLT-bound IgG and IgM levels were also evaluated in the same assay but showed no meaningful signals due to the poor crossreactivity of human antibodies to monkeys (data not shown).
ASO-induced TCP is independent of the fluid-phase alternative complement pathway activation
The ASO-induced fluid-phase complement activation was evaluated using plasma samples collected at various time points. The acute increases in Bb (an alternative pathway complement activation biomarker) and C3a were observed in most monkeys from both sources on Day 1 at 4 h postdose ISIS 405879 treatment, with a higher mean response seen in the Asian-sourced (Cambodian origin) monkeys, driven mainly by two animals (Monkeys 3502 and 3503) (Fig. 4A, B).
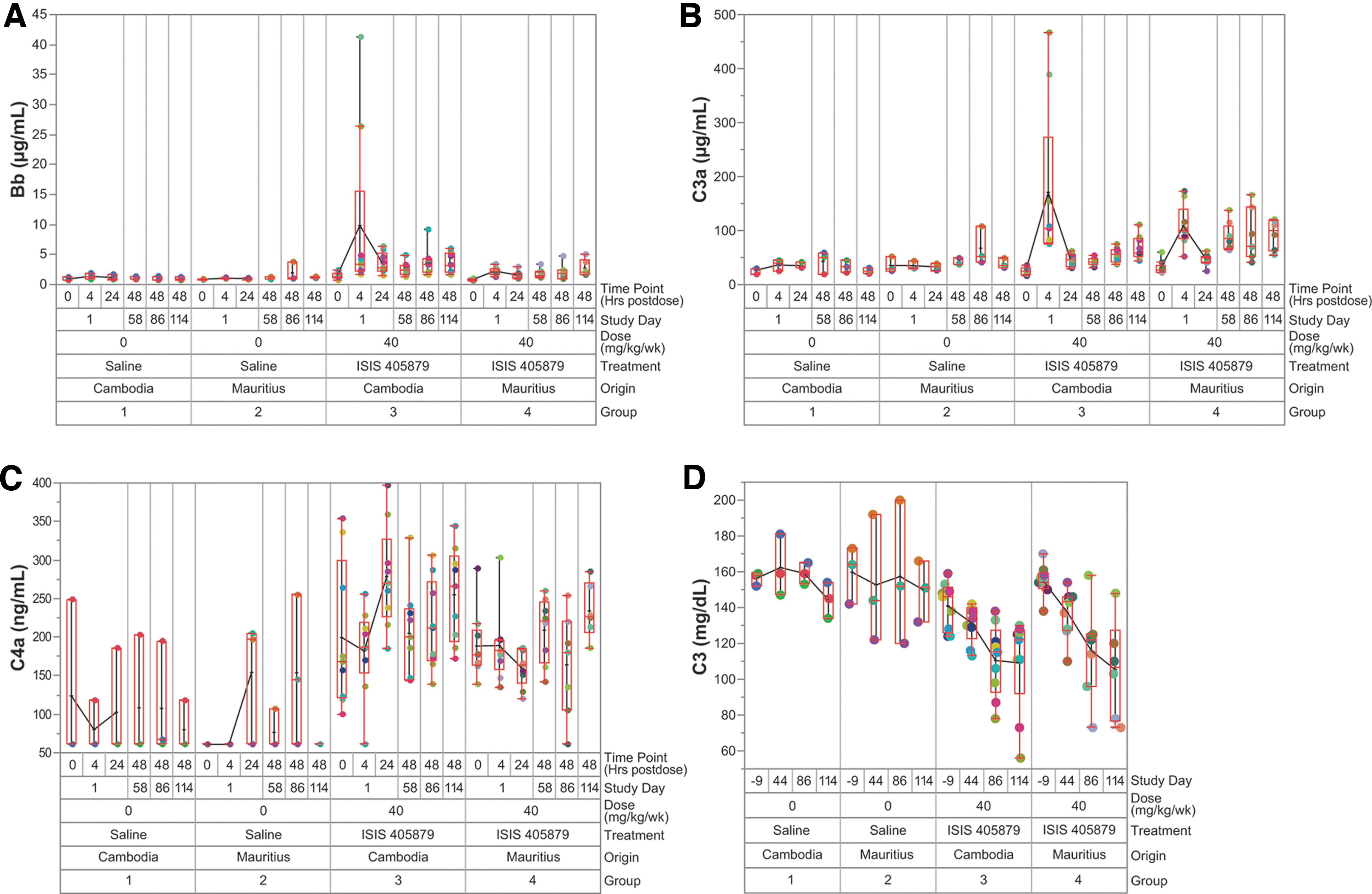
Fluid-phase alternative pathway complement activation was observed in both Asian (Cambodian origin)- and Mauritian-sourced monkeys treated with ISIS 405879 at 40 mg/kg/week. Complement split products Bb
The acute increases in Bb and C3a were mostly resolved by 24 h postdose. However, sustained increases in Bb and C3a levels (two- to threefold from baseline) were seen after repeat ISIS 405879 treatment on Day 114 (48 h postdose) and correlated with ∼30% reduction in C3 in monkeys from both sources (Fig. 4D). There was no apparent increase in C4a (a classical pathway complement activation biomarker) after a single or repeat dose ISIS 405879 treatment (Fig. 4C).
Detecting serum anti-PLT antibodies and epitope-specific anti-GPIIb/IIIa IgG in thrombocytopenic monkeys
Anti-PLT IgG, anti-PLT IgM, epitope-specific anti-PLT IgG (GPIIb/IIIa, GPIa/IIa, GPIb/IX, GPIV, and Class I HLA specific anti-PLT IgG), as well as total IgG and IgM levels were evaluated in the monkey serum collected at various time points. Treatment-related mild to marked increases in anti-PLT IgG from baseline were observed in some of the thrombocytopenic Mauritian-sourced monkeys, but not in the Asian-sourced (Cambodian origin) monkeys (Fig. 5A).
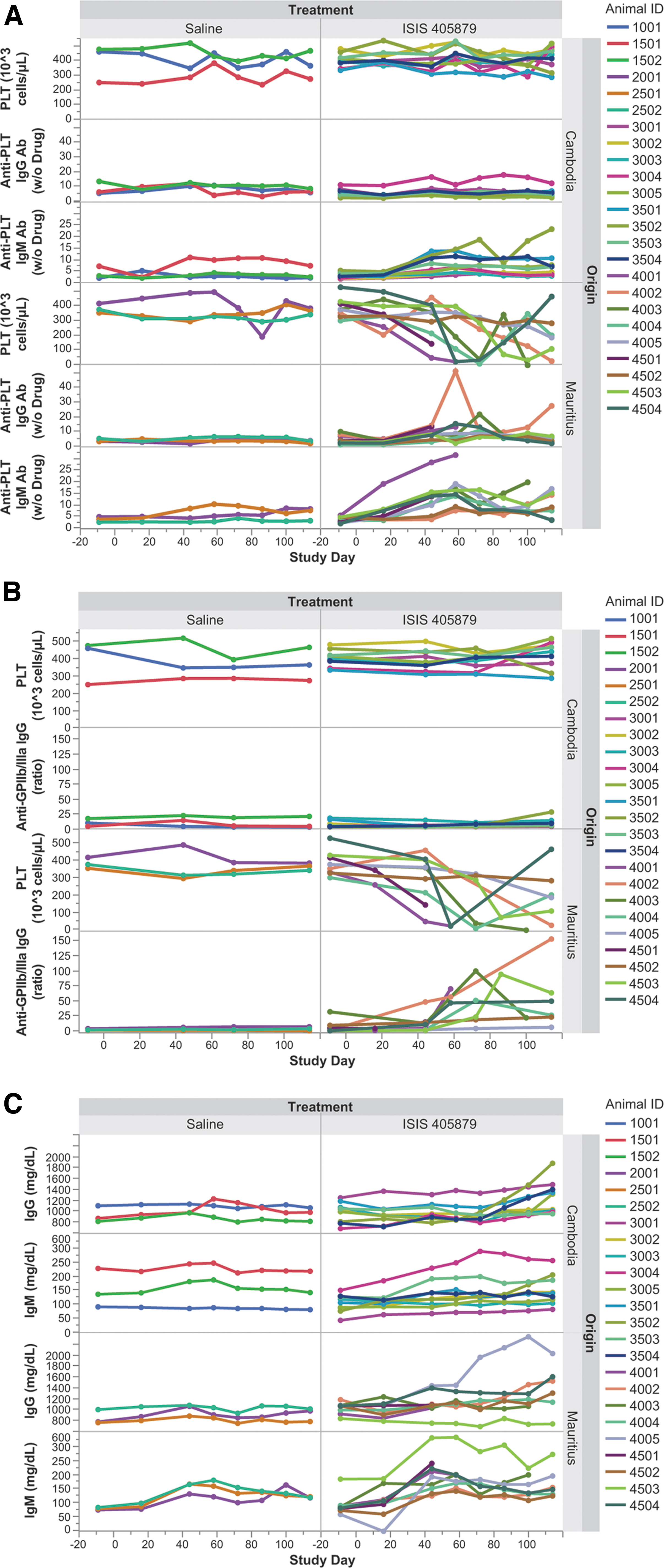
Time-course changes in serum anti-PLT antibodies, anti-GPIIb/IIIa IgG, and total IgG/IgM after 16 weeks of ISIS 405879 treatment. Anti-PLT IgG and IgM antibodies
Further investigation of epitope-specific anti-PLT IgG demonstrated a marked increase in anti-GPIIb/IIIa IgG antibody in all thrombocytopenic Mauritian-sourced monkeys (Fig. 5B). Except for Monkey 4501, who was terminated on Day 56 due to low PLT counts, no sample for anti-GPIIb/IIIa IgG analysis was collected from this animal at the unscheduled necropsy.
Minimal or no increase in anti-GPIIb/IIIa IgG was observed in the two unaffected Mauritian monkeys or Asian-sourced (Cambodian origin) monkeys treated with ISIS 405879. There were no increases in GPIa/IIa, GPIb/IX, GPIV, and Class I HLA-specific anti-PLT IgG in any monkeys with or without PLT reduction (data not shown). Among those thrombocytopenic Mauritian-sourced monkeys, the increases in anti-GPIIb/IIIa IgG generally peaked around the PLT nadirs, and then partially resolved after a long dosing holiday period (Monkeys 4004, 4503, and 4504).
A greater increase in anti-PLT IgM over control was also observed in the Mauritian-sourced monkeys (group mean up to 4.3-fold of baseline) than in the Asian-sourced (Cambodian origin) monkeys (group mean up to 2.7-fold of baseline) with ISIS 405879 treatment (Fig. 5A).
All seven Mauritian-sourced monkeys that developed severe PLT reduction (<50 K/μL) and one of the nonthrombocytopenic monkeys (Monkey 4005) had elevated anti-PLT IgM levels at various stages of treatment. However, an apparent increase in anti-PLT IgM was also observed in some of the Asian-sourced (Cambodian origin) monkeys with normal PLT counts (Monkeys 3501 and 3504) and with one that showed a 30% PLT reduction from baseline (Monkey 3502).
Variable increases in total serum IgG and IgM from baseline were seen in both Asian (Cambodian origin)- and Mauritian-sourced monkeys treated with ISIS 405879, with a comparable increase in total serum IgG (up to 1.3-fold of baseline), but a slightly higher group mean increase in total serum IgM in Mauritian-sourced monkeys [up to 1.5- vs. 2.1-fold of baseline for Asian (Cambodian origin)- and Mauritian-sourced monkeys, respectively] (Fig. 5C).
No evidence of treatment-related increase in anti-PF4 IgG in monkeys with TCP
Anti-PF4 antibodies were measured to assess the potential for HIT. Increases in anti-PF4 IgG level from baseline were observed in selected control (Monkey 2501) and ISIS 405879-treated Mauritian-sourced monkeys with or without severe PLT reduction (Monkeys 4002 and 4005). These changes were considered within the normal variability for this parameter, thus not supporting a HIT-like mechanism for ASO-induced TCP. Higher than control anti-PF4 IgM levels were most evident in one Asian-sourced (Cambodian origin) monkey (Monkey 3502) and several Mauritian-sourced monkeys (eg, Monkey 4001 and 4002 and 4503) (Fig. 6).
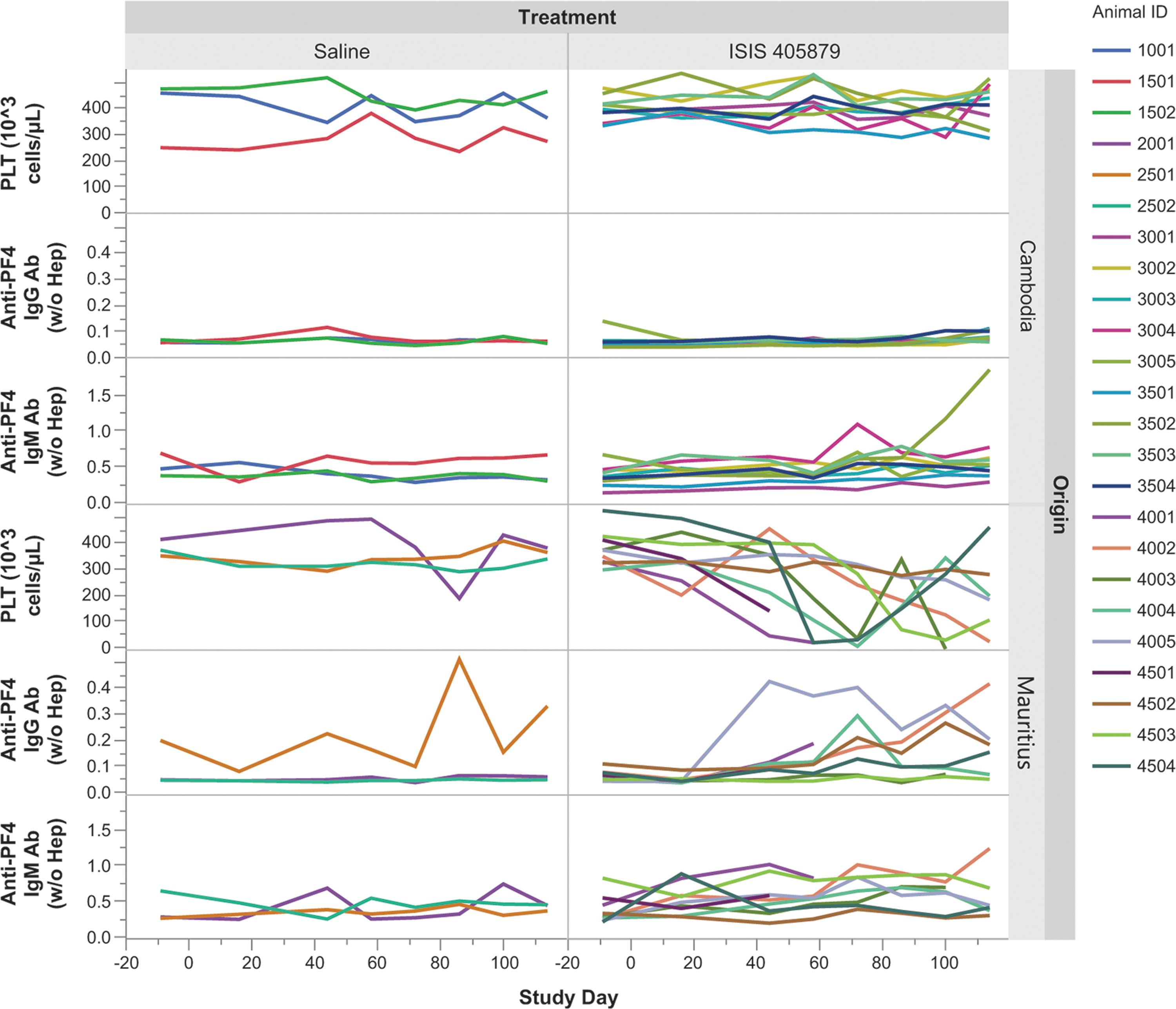
Time-course changes in anti-PF4 IgG and IgM antibodies after 16 weeks of ISIS 405879 treatment. The corresponding PLT levels at matched time points are presented in the top panel. Individual data from each monkey are plotted. PF4, PLT factor 4.
No clear evidence of PLT activation in vivo and in vitro with ISIS 405879 treatment
In vivo and in vitro PLT activation was evaluated by measuring CD62p on PLT. For the in vivo study, PRPs were collected from saline or ISIS 405879-treated monkeys at different time points and measured for CD62 expression on PLTs. A marked increase in CD62p-positive PLT (%) was only observed in one Mauritian-sourced monkey (Monkey 4002) on Day 114 (Fig. 7). Additional analyses for PLT activation, including PLT microparticle and leukocyte-PLT aggregation assay, were also conducted using samples collected at different time points but showed no evidence of PLT activation in monkeys with ISIS 405879 treatment (data not shown).
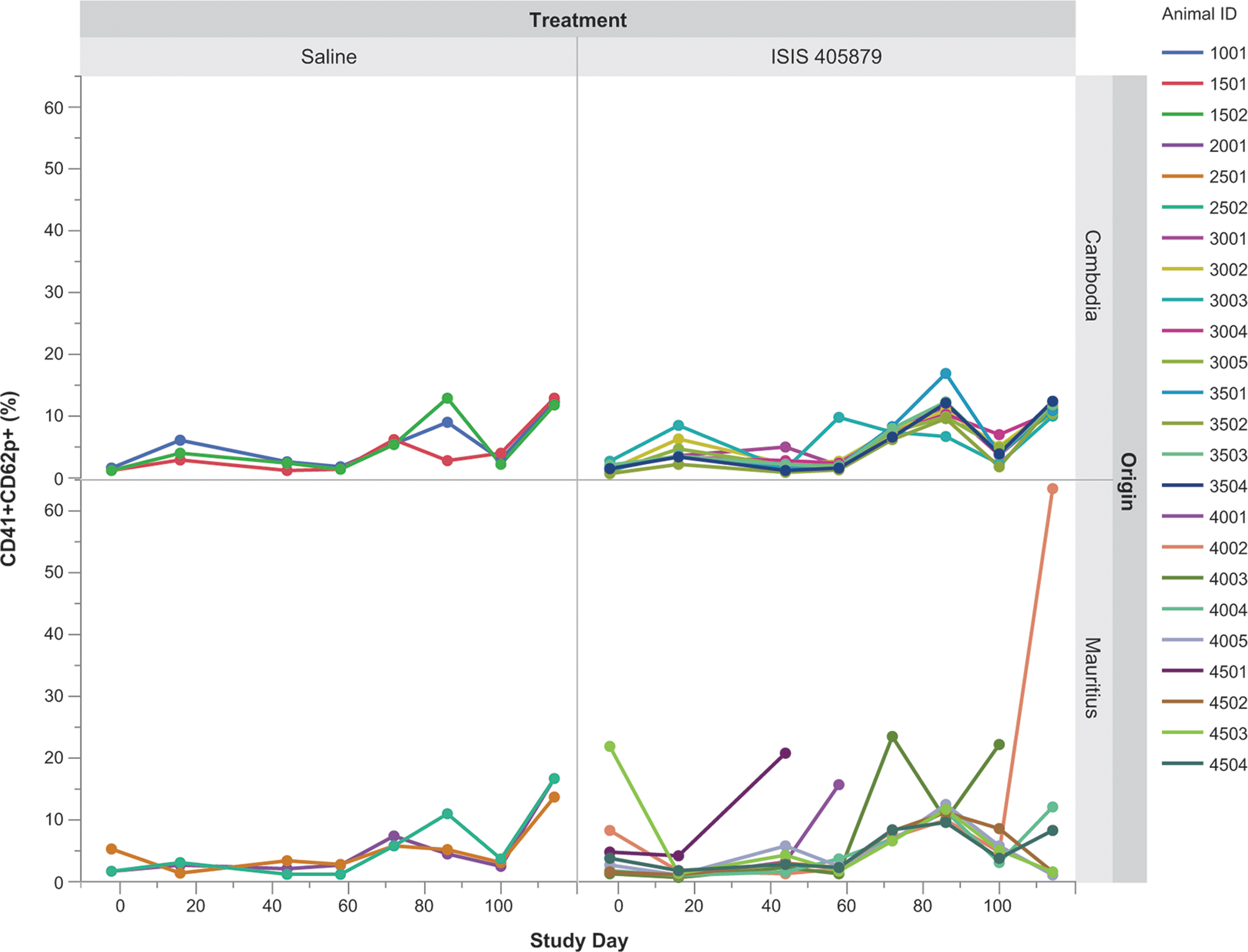
Lack of strong evidence of in vivo PLT activation in monkeys after 16 weeks of ISIS 405879 treatment. In vivo PLT activation was evaluated by flow cytometry using antibodies against CD41 (total PLT) and CD62P (activated PLT). PRPs from control and ISIS 405879-treated monkeys were collected at various time points up to Day 114. The data are expressed as % of CD41+ and CD62p+ PLT over total PLT for individual monkey.
In the in vitro study, PRPs collected from naïve Mauritian- and Asian-sourced (Cambodian origin) monkeys were incubated with increasing concentrations of ISIS 405879 for up to 200 μM. No statistically significant increase of CD62p-positive PLT (%) in ISIS 405879-treated PRPs collected from either Asian (Cambodian origin)- or Mauritian-sourced monkeys was observed (Fig. 8). Statistically significant increases of CD62p-positive PLT (%) were seen in ADP (20 μM) and ISIS 120704 (200 μM), an ASO that contains a CpG motif.
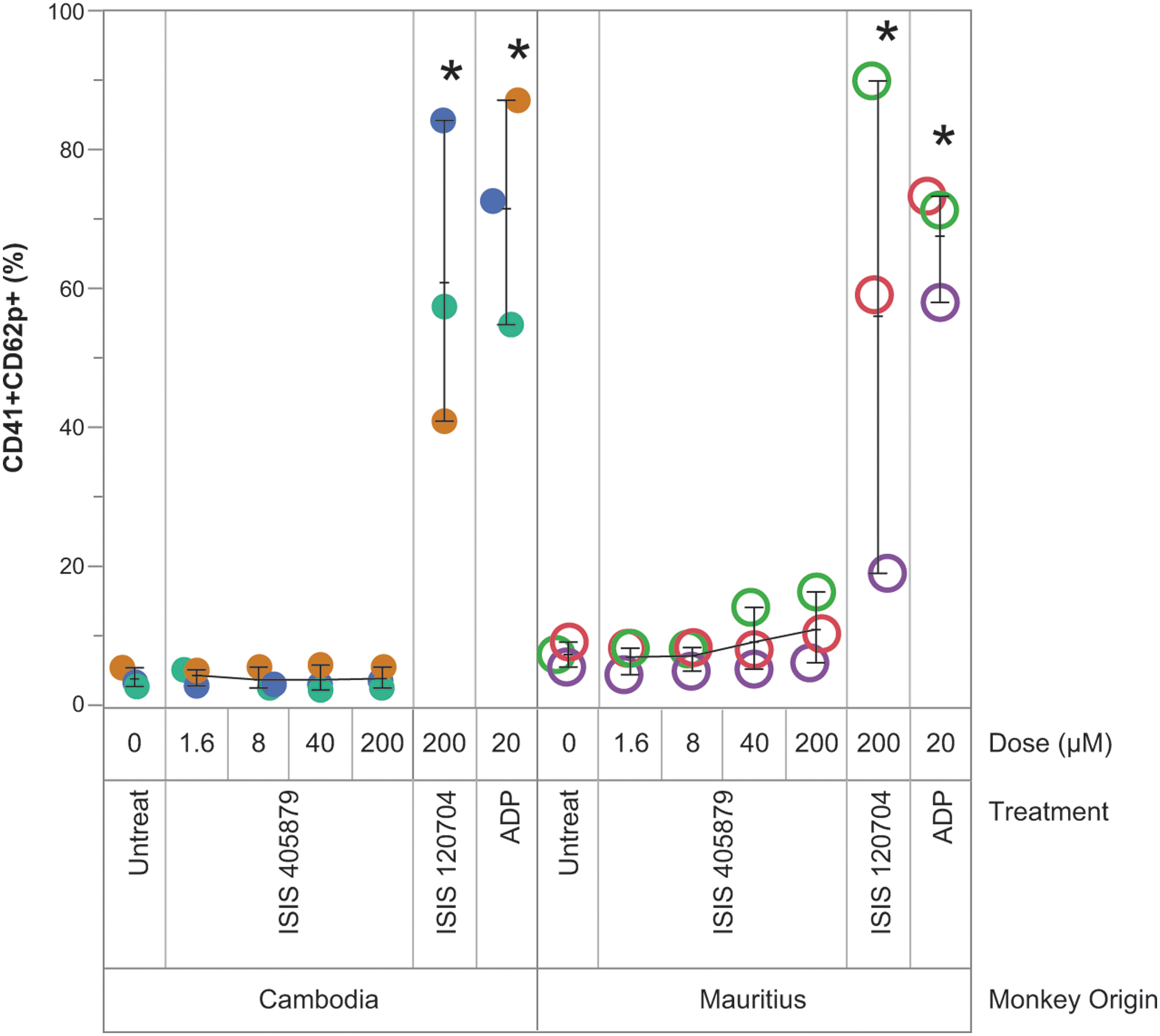
Absence of PLT activation in vitro after incubation of ISIS 405879 with PRPs from naïve monkeys. ISIS 120704 is a PS ASO that possesses CpG dinucleotide motifs and was used as a positive control for CD62P upregulation in monkey PLTs. Additional assay positive control, ADP was also used. Data represent individual values of PLTs collected from three naïve Asian- (Cambodian origin) (●) and Mauritian-sourced (○) monkeys. *P value <0.01 relative to untreated control (one-way analysis of variance with Dunnett's). ADP, adenosine diphosphate; ASO, antisense oligonucleotide.
Increase in vWF with ISIS 405879 treatment
An increase in vWF level was seen previously in monkeys treated with ISIS 104838, another 2′-MOE ASO, concurrent with the consistent gradual PLT reduction in monkeys [7]. A similar increase in vWF antigen level from baseline was observed in eight of nine Asian-sourced (Cambodian origin) monkeys and all Mauritian-sourced monkeys (Fig. 9). This increase in vWF antigen level started as early as Day 16. However, no correlation between vWF antigen increase and PLT reduction was observed, suggesting that the increase in vWF is likely a nonspecific ASO effect in monkeys at the high dose used in this study.
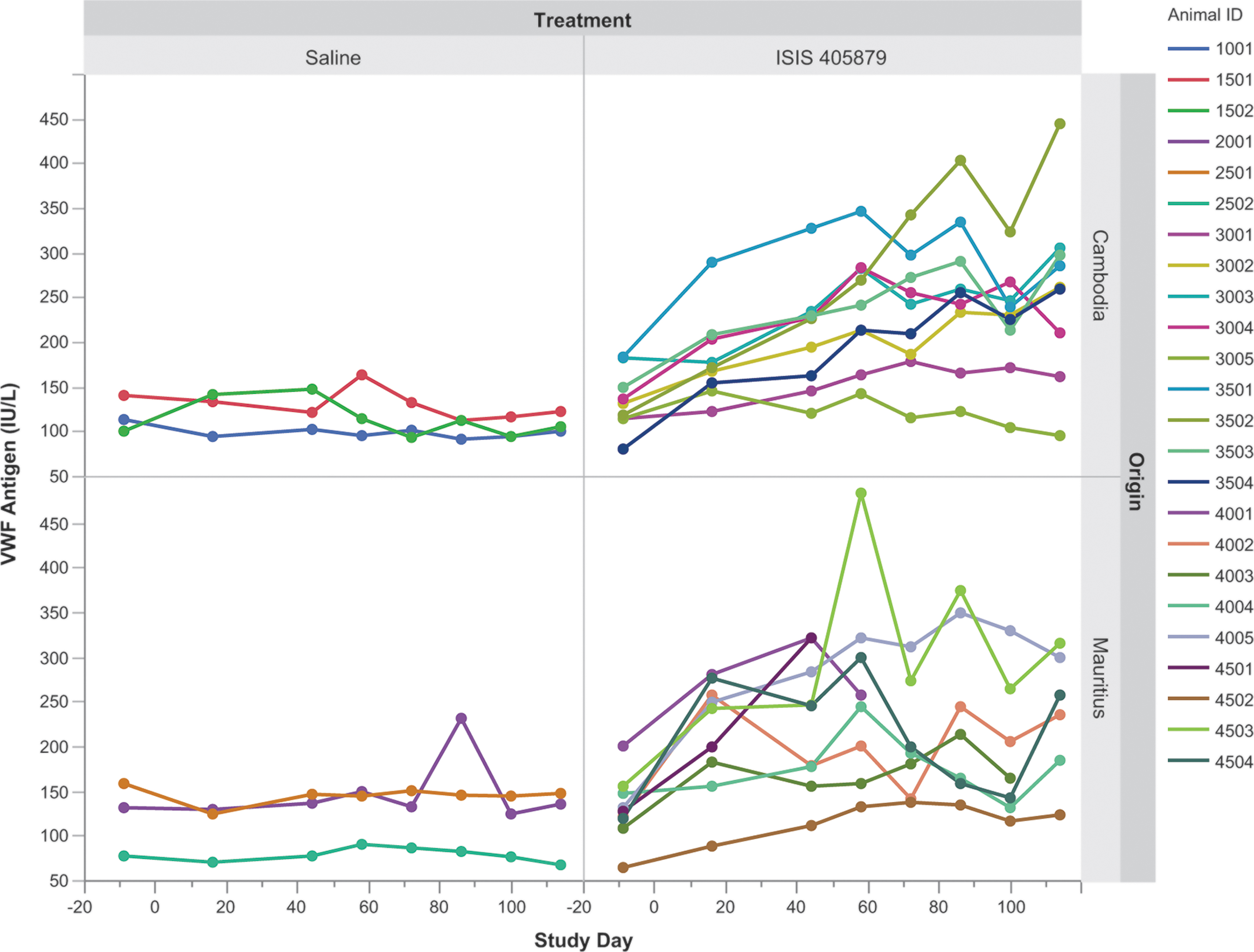
Time-course changes in serum vWF antigen level after 16 weeks of ISIS 405879 treatment. Individual data from each monkey are plotted. vWF, von Willebrand factor.
Changes in proinflammatory parameters
ISIS 405878-induced proinflammatory effect was assessed using a 15-analyte cytokine/chemokine panel, which included GM-CSF, IFN-γ, IFN-α, TNF-α, MCP-1, IL1-β, IL-4, IL-5, IL-6, IL-8, IL-10, IL-12, IL-13, IL-17A, and IL-21. Most of the analyte results were below the detection limits. Increases in MCP-1 were observed in selected monkeys from both sources, with the highest levels seen in one Asian-sourced (Cambodian origin) monkey (Monkey 3504 on Days 100 and 114) and one Mauritian-sourced monkey (Monkey 4002 on Day 100) (Fig. 10A).
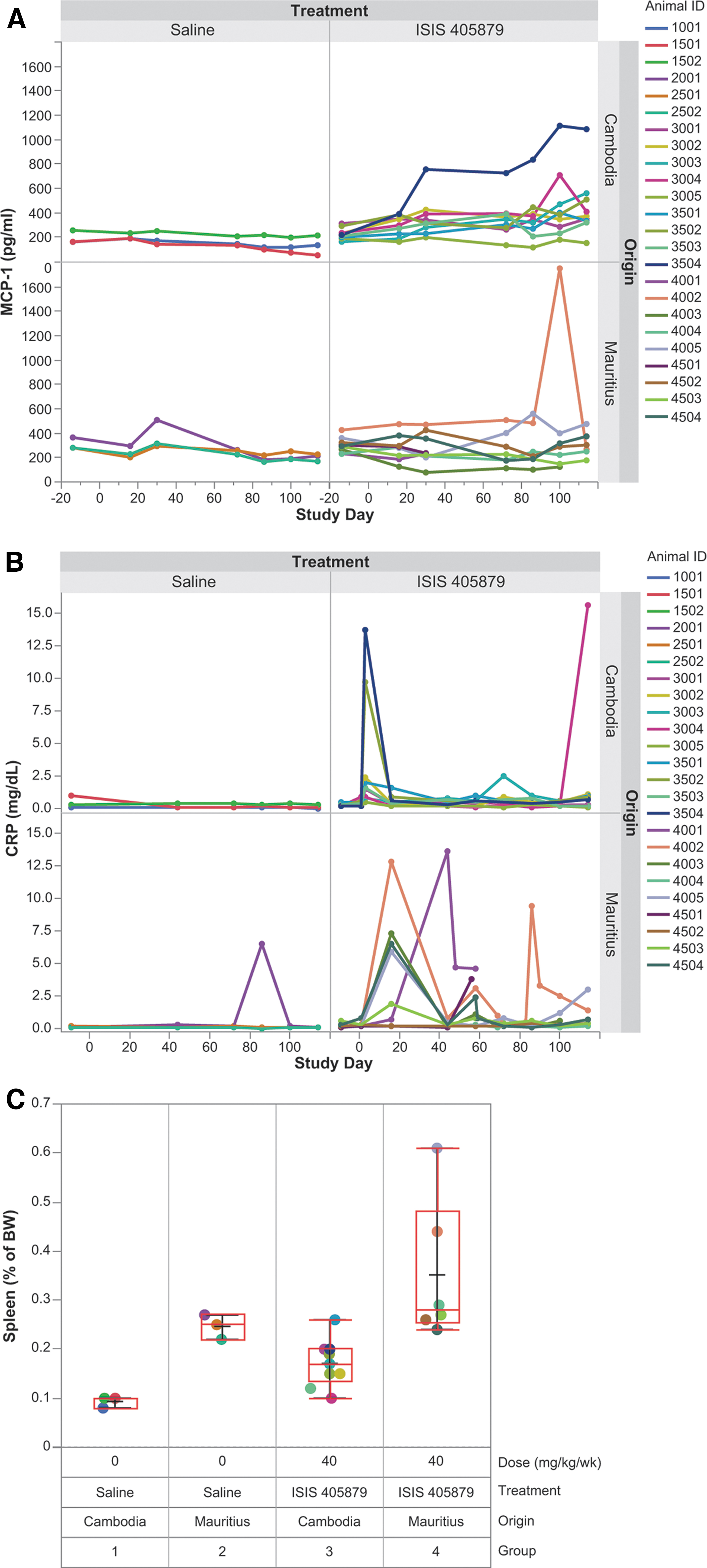
Changes in proinflammatory parameters after 16 weeks of ISIS 405879 treatment. Serum MCP-1
Additional proinflammatory parameters, such as C-reactive protein (CRP) and spleen weight, were also measured. A sporadic transient increase in CRP was observed in one Mauritian control monkey, and selected monkeys from both sources treated with ISIS 405879 (Fig. 10B). A ∼1.8- and 1.5-fold increase in spleen body weight ratio over control was observed in Asian-sourced (Cambodian origin) and Mauritian-sourced monkeys treated with ISIS 405879, respectively. Interestingly, the spleen weight of the control monkeys from the Mauritian source was about 2.6-fold of the Asian-sourced (Cambodian origin) monkeys in this study (Fig. 10C).
Discussion
The two phenotypes of PLT reductions in monkeys induced by 2′-MOE ASO treatment have been well characterized previously [3]. This article is focused on the potential mechanism of sporadic severe phenotype 2 PLT reduction (<50 K/μL) using cynomolgus monkeys from different sources. The key characteristics of this type of PLT reduction in monkeys or humans are dose- and sequence dependent with delayed onset (typically after 3–4 months of treatment), and reversible upon dose suspension and/or steroid treatment [3].
Anecdotal evidence suggests that Mauritian-sourced cynomolgus monkeys experience a higher incidence of ASO-induced severe TCP than monkeys from various sources in Asia. Ionis own experience with ISIS 405879, a 2′-MOE ASO previously developed as a treatment for hypercholesterolemia, demonstrated a relatively high incidence of severe TCP in Mauritian-sourced cynomolgus monkeys (Noveprim Ltd.) but not in the Asian-sourced (Chinese origin) monkeys after 3 months of treatment. This increased susceptibility of Mauritian-sourced monkeys to severe TCP could be due to their limited genetic diversity.
For example, fewer major histocompatibility complex haplotypes are present in the Mauritian-sourced cynomolgus monkeys [12], likely resulting from inbreeding due to the geographic isolation of the island of Mauritius [13,14]. The patterns of severe PLT reduction observed in the Mauritian-sourced monkeys appear identical to those observed in Asian-sourced monkeys after 2′-MOE ASO treatment, although with a greater incidence, and thus can be used as a model to study the mechanisms of severe PLT reduction.
The higher incidence of 2′-MOE ASO-induced severe TCP in Mauritian-sourced monkeys has further been confirmed in this investigative study, with seven of the nine Mauritian-sourced monkeys developing TCP but none in the Asian-sourced (Cambodian origin) monkeys after 16 weeks of treatment with ISIS 405879 at 40 mg/kg/week. The two crucial pieces of mechanistic evidence that emerged from this study were the deposition of C3d/C4d on monkey PLT surface as detected by flow cytometry and increased levels of serum anti-GPIIb/IIIa IgG antibody in the thrombocytopenic Mauritian-sourced monkeys as measured using Luminex bead array assay.
The marked increase in PLT-bound C3d/C4d was observed in all Mauritian-sourced monkeys with TCP but not in monkeys with normal PLT counts from either source. The peak levels of C3d/C4d deposition generally correlated with PLT nadirs for those monkeys that developed TCP. Additional data from a recent follow-up study also showed increased C3d deposition (and C4d to a lesser extent) on Asian-sourced (Cambodian origin) monkeys who developed TCP (data not shown). This suggests that the complement split product fixation on the PLT surface could be one of the mechanisms for ASO-induced PLT reduction in monkeys, regardless of the source of the monkeys.
The C3d/C4d fixation on the PLT surface is independent of ASO-mediated fluid-phase alternative complement activation in monkeys, a known class effect of 2′-MOE ASO in monkeys at high doses (approximately ≥10 mg/kg with SC administration) [15]. The increases in plasma Bb (alternative pathway complement activation marker), C3a, and decrease in total C3 after repeat treatment were seen in all monkeys given at 40 mg/kg/week ISIS 405879. There were no clear differences between Mauritian-sourced and Asian-sourced (Cambodian origin) monkeys.
The acute fluid-phase complement activation does not directly lead to acute PLT reduction, as demonstrated in this study as well as from our previous experiences [15,16]. Most importantly, there was no evidence of classical pathway complement activation in the fluid phase, as the C4a level was similar to the baseline after 16 weeks of ISIS 405879 treatment. These data suggested that the increased C3d/C4d deposition on the PLTs was not a direct consequence of ASO-induced fluid-phase alternative pathway complement activation.
There were also increases in PLT crossreactive immunoglobulins, such as anti-PLT IgG and anti-PLT IgM, in monkeys treated with ISIS 405879. The most well-correlated change with PLT reduction was the marked increase in serum anti-GPIIb/IIIa IgG in those monkeys that developed TCP. PLT reduction induced by anti-GPIIb/IIIa IgG can be either mediated by Fc-mediated phagocytosis or following classical pathway complement activation on the PLT surface by C3b-mediated phagocytosis and PLT lysis [17–19]. Activation of classical complement pathway on the PLTs has previously correlated with enhanced PLT clearance in patients with immune thrombocytopenic purpura (ITP) due to PLT autoantibody specifically to GPIIb/IIIa and GPIb/IX [20,21].
Increases in serum anti-PLT IgM and anti-PF4 IgM were also observed in ISIS 405879-treated monkeys from both sources. Although these changes were not as closely correlated with severe TCP, the increases were more pronounced in Mauritian-sourced monkeys in this study compared with Asian-sourced (Cambodian origin) monkeys. However, the increases in serum anti-PLT IgM and/or anti-PF4 IgM alone did not appear to be sufficient to cause a severe reduction in PLT counts in monkeys, as elevated levels of both antibodies were also seen in selected monkeys with normal PLT counts.
Increases in serum anti-PLT IgM and anti-PF4 IgM parameters were reported for monkeys treated with a similar 2′-MOE ASO (ISIS 104838) [7]. These increases in total serum IgM have been attributed to a polyclonal antibody response secondary to the nonspecific innate immune cell activation by the 2′-MOE ASO [16]. In addition, increases in CRP and spleen weight were attributed to the proinflammatory effects, and were comparable between the Asian (Cambodian origin)- and Mauritian-sourced monkeys given ISIS 405879 treatment.
The binding of anti-PLT IgM to PLTs could also activate classical complement pathway activation either in the form of the antibody or as part of an immune complex. PLT-bound C3 has been observed in some ITP patients, even though this increase in C3 deposition on the PLT is generally correlated with PLT-bound IgG [22–24]. A strong correlation between the amount of IgM and C3 on PLTs was also confirmed in a subset of ITP patients, suggesting the presence of C3 on PLT surface due to complement fixation by IgM [25,26].
Polyreactive natural IgM has been shown to initiate complement activation by PF4/heparin complexes through the classical pathway [26]. The ASOs may play a role in the interaction of anti-PLT IgM or anti-PF4 IgM with PLT and complement activation, but it is unclear to what extent based on the current results. Even though monkey PLT-bound IgG and IgM were not evident in this study (due to a technical issue of antibody crossreactivity), increased PLT-bound IgG (and to a lesser extent PLT-bound IgM) was later demonstrated in a follow-up study using a more optimized assay mostly in monkeys that developed TCP (data not shown).
The data from this study are consistent with previous findings, and further confirm that ASO-related TCP is not due to a direct bone marrow effect but rather the result of increased peripheral consumption of PLT [3,7]. Increased MPV levels were observed in all affected monkeys and correlated with megakaryocyte hyperplasia in most of the thrombocytopenic monkeys, suggesting normal bone marrow response to PLT reduction.
Both in vitro (P-selectin CD62P upregulation) and in vivo assays (CD62P upregulation, leukocyte-PLT aggregation, and release of PLT extracellular vesicles) performed with ISIS 405879 showed a lack of PLT activation at concentrations that are high multiples (> ∼100-fold) of clinically relevant Cmax plasma concentrations [28]. However, detecting PLT activation in vivo could be technically challenging as activated PLTs may be quickly cleared from circulation.
Consistent with previous studies, the data did not support the HIT-type mechanism for PLT reduction due to the lack of treatment-related anti-PF4 IgG increase. However, we reaffirmed the presence of treatment-emergent anti-PF4 IgM, especially in the Mauritian-sourced monkeys. The elevated anti-PF4 IgM reflects the increase in total IgM and is not clinically meaningful for the diagnosis of HIT. Increased vWF was seen previously in monkeys that developed the moderate phenotype 1 PLT reduction [7]. However, the data from this study suggested that the increase in vWF was likely a high-dose effect with 2′-MOE ASOs in monkeys and not necessarily related to the PLT changes.
Unlike the traditional drug-induced TCP, the mechanism for ASO-induced severe PLT reduction in monkeys is likely attributed to innate immune cell activation (e.g., monocytes and macrophages) as the result of ASO direct interaction with innate immune receptors, such as TLR9 and TRL7 [29]. The binding and activation are the greatest for CpG motif sequences but can happen to non-CpG ASOs at higher concentrations [30].
The innate immune cell activation is a known sequence- and dose-dependent effect for phosphorothioate-modified ASOs, and results in cytokine/chemokine release, B cell activation/proliferation, and polyclonal antibody production (including PLT crossreacting IgG and IgM) [7]. The severe PLT reduction in monkeys appears to be determined by epitope specificity or complement fixation on the PLTs.
Using the Mauritian-sourced monkey model makes it possible to study the potential mechanism (s) for ASO-induced TCP, since the incidence is very low in the Asian-sourced monkeys. Increases in PLT-bound C3d/C4d and serum anti-GPIIb/IIIa IgG signals were confirmed in both Mauritian -and Asian-sourced monkeys who developed severe TCP in the follow-up study, suggesting similar mechanisms for PLT reduction in those two different sources of the monkeys with ASO treatment. Thus, the Mauritian-sourced monkeys may be representative of the subset of Asian-sourced monkeys that are more sensitive to the ASO-induced TCP, likely due to some unknown predisposed conditions.
The causes for the high susceptibilities of Mauritian-sourced monkeys are still unclear. Preliminary data from the genetic profiling of Mauritian versus Asian-sourced (Cambodian origin) monkeys from this study have identified ∼30 “risk-increasing alleles” that were associated with severe PLT declines. These alleles have very low allele frequency in Asian-sourced monkeys but high in Mauritius-sourced monkeys at baseline (unpublished internal data).
The relevance of the current mechanism of C3d/C4d deposition on PLT in monkeys to the TCP observed in the clinic with the 2′-MOE ASOs is still unclear. Severe cases of TCP were reported previously with two different 2′-MOE ASOs (inotersen and volanesorsen) in two phase 3 clinical trials in two rare diseases, hereditary transthyretin amyloidosis with polyneuropathy and familial chylomicronemia syndrome, respectively [31].
Based on the specific context of these observations, it is likely that factors from the underlying disease conditions may contribute to the sensitivity to the TCP in these patients [4,6]. The implicated mechanisms for ASO-induced TCP in monkeys provide some general insights into the potential pathways in a monkey model with a very distinct genetic background. The information derived from this monkey model provides some guidance for testable hypotheses in humans, but there are likely differences in the exact mechanism between Mauritian monkeys and the specific patient populations studied.
The cause for the high incidence of ASO-induced TCP in Mauritian-sourced monkeys is still under investigation. The genetic profiling of Mauritian- versus Asian-sourced (Cambodian origin) monkeys from this study is ongoing. A follow-up study comparing the sensitivity of different sources of Mauritian-sourced monkeys (monkey farm effect) versus Asian-sourced (Cambodian origin) monkeys for 2′-MOE ASO-induced phenotype 1 (moderate) and phenotype 2 (severe) PLT reduction has just been completed. A clear understanding of the mechanism of these PLT phenotypes will enable us to identify and eliminate the ASO sequences that may lead to severe TCP in the clinical setting, and help monitor, manage, and mitigate the potential PLT effects.
Footnotes
Acknowledgment
The authors thank Tamika Holmes for her administrative support.
Author Disclosure Statement
L.S., J.S., T.Z., J.A.E., S.P.H., and P.N. are employees of Ionis Pharmaceuticals, Inc.; all hold stock in the company. A.W. and S.O. are Altasciences employees. B.R.C. is a Versiti, Inc., employee.
Funding Information
No funding was received for this article.