
Abstract
Abstract
Osmostress triggers profound adaptive changes in the physiology of the cell with a great impact on gene expression. Saccharomyces cerevisiae has served as an instructive model system to unravel the complexity of the stress response at the transcriptional level. The main signal transduction pathways like the HOG (high osmolarity glycerol) MAP kinase cascade or the protein kinase A pathway regulate multiple specific transcription factors to accomplish large changes in the expression pattern of the genome. Transcription profiling and proteomic studies give us an idea about the impact of osmostress on gene expression and the overall protein composition. Recent genome wide location studies for several transcription factors and signaling kinases involved in the transcriptional stress response shed light on the genomic organization of the osmostress response at the level of the dynamic association of regulators with chromatin. Finally, global surveys of mRNA stability complete our picture of the mechanisms underlying the massive reprogramming of global gene expression, which leads to efficient adaptation to osmotic stress.
Background
The Impact of Osmotic Stress on Global Gene Expression: Reprogramming the Genome Upon Stress
Several global transcription profiling experiments have been performed to characterize the changes of gene expression in the yeast genome upon hyperosmotic and salt stress (Gasch et al., 2000; Ni et al., 2009; Posas et al., 2000; Rep et al., 2000; Yale and Bohnert, 2001). A remarkable feature of the transcriptional osmostress response is its extremely transient nature. Moreover, the time course of the transcriptional response depends critically on the severity of the osmotic stress (Posas et al., 2000; Rep et al., 1999a; Yale and Bohnert, 2001). A “typical” upregulated osmoresponsive yeast gene will be fully induced within 5–10 min upon mild salt stress (0.4 M NaCl), whereas the same gene will have its induction maximum much later (30–60 min) upon severe salt stress (1 M NaCl). A likely explanation for this phenomenon is that osmotic stress interferes with protein binding to chromosomal DNA, and therefore generally with transcription due to the critical ionic strength in the nucleus in the first instances of the osmotic insult (Proft and Struhl, 2004). In this scenario, a higher salt concentration will prolong the adaptation phase until efficient transcription can occur, and therefore delay the expression of defense genes under more severe conditions.
Genome-wide expression analyses give us an idea about the impact of osmotic stress on global transcription. However, the estimated number of osmoresponsive genes depends on the intensity and kinetics of the osmotic stress applied to the cells and the cutoff chosen to define a significantly stress-regulated locus. Strongly upregulated genes represent 3–7% of the genome (200–350 different genes) (Posas et al., 2000; Rep et al., 2000; Yale and Bohnert, 2001). A very recent, kinetic study of global gene expression raises this number to 23% of the genome (nearly 1,400 genes) including many moderately regulated genes over a time course of salt stress (Ni et al., 2009).
The classification of these activated genes into biological functions gives important information about the different physiological processes that are remodeled upon osmostress (Fig. 1). Genes involved in glycerol, trehalose, and glycogen metabolism are activated because of the osmolyte function of the metabolites, which has been shown to be critical for glycerol as the main osmolyte (Hohmann, 2002a). Sugar transporters and genes involved in sugar metabolism are targeted, which might reflect the need to modify the sugar supply to the cell in hyperosmotic conditions. A major group of osmoinducible genes is functionally connected to the defense of oxidative stress, which is caused by hypertonic conditions (Koziol et al., 2005). In the same vein, several nuclear genes encoding mitochondrial functions are upregulated, which have important antioxidant functions upon osmostress (Pastor et al., 2009). The great number of osmoinducible genes involved in redox metabolism further highlights the importance of redox maintenance as a manner to combat the oxidative stress caused by salt stress. Protein denaturation represents yet another toxic effect of osmostress, which seems to be counteracted by the induction of genes encoding numerous heat-shock proteins and chaperones. A significant number of transcriptionally activated genes has functions in signal transduction. Less understood is the importance of the selective induction of cell surface proteins, and still we do not know the physiological role for many yeast genes that show robust osmoinducibility.
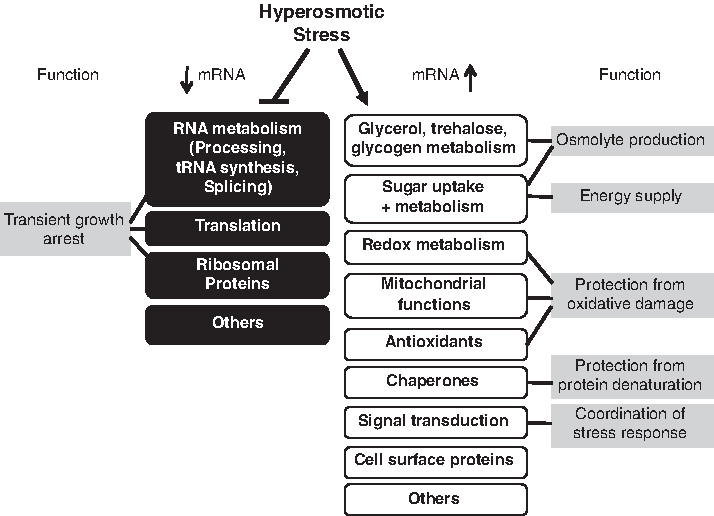
Overview of the differentially expressed gene clusters upon hyperosmotic stress.
On the other hand, a large set of genes shows an opposite regulation and their transcript levels diminish upon osmostress (Gasch et al., 2000; Rep et al., 2000). One main group of downregulated genes is involved generally in growth-related processes or RNA metabolism like RNA processing, splicing, translation, and tRNA synthesis. The second cluster of osmorepressed genes encodes ribosomal proteins. The general shutdown of ribosomal biogenesis, translation, and RNA metabolism likely reflects the transient growth arrest provoked by osmostress. It is important to note that the number of downregulated transcripts can be as high as the upregulated transcripts, and affects more than 1,300 genes according to kinetic expression profiling experiments upon salt stress (Ni et al., 2009).
Taken together, transcription profiling of the yeast osmostress response reveals the complexity of the physiological readjustments necessary to adapt to the stress. We next focus at how stress signaling modulates the genomic expression profile via specific transcriptional activators and repressors in such a profound way as described by the various transcriptomic surveys. However, not all changes of mRNA abundance will be attributable to changes in the transcription rate. Especially subtle variations in the mRNA levels might be explained by posttranscriptional regulation, which will be discussed in the last chapter of this review.
Changes in the Chromatin Landscape Upon Osmotic Stress: Dynamic Association of Transcription Factors, Signaling Kinases and Chromatin Modifiers to the Genome
A great number of specific transcription factors (TF) is involved in coordinating the reprogramming of gene expression upon osmotic stress. This is not surprising because many genes respond in different ways to this stimulus, as has been described by genomic expression analyses. In general, the activity of these transcription factors is modulated by upstream signaling pathways to adapt the transcription pattern upon stress (Fig. 2). Many studies on specific osmostress-regulated gene promoters in yeast have identified multiple mechanisms that confer stress activated transcription, which will be discussed below. In the end of this article we will also discuss the mechanisms that lead to a controlled repression of gene expression upon stress.

The principal signaling compounds and specific transcription factors involved in the yeast osmostress response. Signaling molecules are depicted in blue, representing the Hog1 MAP kinase, the Sch9 protein kinase, protein kinase A (PKA), and the protein phosphatase calcineurin. Transcription factors are depicted in green, representing all regulators with experimentally confirmed implication in osmostress regulation. The mechanisms leading to modulation of Yap6 and Sch9 functions upon osmostress are currently unknown, and therefore represented by dotted arrows. Also, in cases were TFs are placed under control of the Hog1 MAP kinase by genetic rather than direct biochemical evidence, this is indicated by interrupted lines. See text for further details.
The HOG (High Osmolarity Glycerol) MAP kinase signal transduction pathway has the highest impact on osmostress regulated transcription (de Nadal et al., 2002; Westfall et al., 2004). This is reflected by the fact that loss of function of its terminal MAP kinase Hog1 leads to severe osmosensitivity and to impaired transcriptional activation at most osmo-inducible genes (Posas et al., 2000; Rep et al., 2000). Hog1 is rapidly activated by phosphorylation upon hyperosmotic stress and translocates to the nucleus where it orchestrates the genomic transcription program at multiple levels (de Nadal and Posas, 2010). It is important to note that changes of the gene expression pattern upon osmostress occur in a transient manner. This implies that signaling components and TFs involved in the osmoresponse change their activities only for a very limited time before they return to their initial state. This is well documented by the highly transient nuclear localization of Hog1, which is reverted immediately after the execution of the transcriptional program within a few minutes (Ferrigno et al., 1998; Reiser et al., 1999).
Hog1 directly contacts and phosphorylates the specific transcription factors Sko1, Hot1, and Smp1 (Alepuz et al., 2003; de Nadal et al., 2003; Proft et al., 2001; Proft and Serrano, 1999; Rep et al., 1999b). Sko1 is a transcriptional repressor/activator of the bZIP family, and the only TF identified so far within the osmostress response with repressor function under normal growth conditions and activator function upon stress. Hot1 and the MADS-box family member Smp1 belong to structurally different classes of TFs and activate transcription upon stress. The Hog1 MAP kinase regulates even more specific TFs, as genetic evidence identifies the Msn1, Msn2, Msn4, and Yap4 activators as downstream targets of the HOG pathway (Martinez-Pastor et al., 1996; Nevitt et al., 2004; Rep et al., 1999b; Schuller et al., 1994). It is important to note that with the exception of the highly homologous Zn-finger transcription factors Msn2/4 and the modest similarity between Msn1 and Hot1, the TFs acting downstream of HOG are structurally unrelated, which already indicates that each TF might recognize discrete sets of binding sites in the genome.
Regulation of TF activity by Hog1 goes far beyond the phosphorylation of the TF. In many cases, the TF forms a stable complex with the MAP kinase at the 5′-upstream chromosomal region of osmoresponsive genes, which can be experimentally shown by in vivo chromatin immunoprecipitation (ChIP) (Alepuz et al., 2001; Proft and Struhl, 2002). The stable association of Hog1 with chromatin is restricted to osmostress-regulated loci and is general as Hog1 can be crosslinked to osmostress-induced promoters along the yeast genome in global location analyses (ChIP–Chip) (Pascual-Ahuir et al., 2006; Pokholok et al., 2006; Proft et al., 2006). Interestingly, Hog1 association with the genome can be achieved by different sets of TFs depending on the specific stress-regulated promoter. This has been described by the analysis of the osmoinducible GPD1, CTT1, or GRE2 genes, at which the MAP kinase gets recruited by either Hot1, Msn2/4, or Sko1, respectively (Alepuz et al., 2001; Proft and Struhl, 2002).
Recruitment of Hog1 to promoters is necessary to coordinate chromatin remodeling and assembly of the transcriptional preinitiation complex upon stress. Once stably associated with chromatin, Hog1 has diverse functions in the stimulation of gene expression. The MAP kinase can directly recruit the RNA polymerase II machinery in the case of Hot1-regulated genes (Alepuz et al., 2003). This is further demonstrated by the observation that Hog1, when artificially tethered to promoter DNA, can stimulate transcription. Furthermore, Hog1 is directly or indirectly involved in the recruitment of chromatin modifying or basal transcription complexes to stress-activated promoters. These include the Rpd3 histone deacetylase, the SAGA coactivator complex, the SWI/SNF chromatin remodeling complex, and the Mediator complex (de Nadal et al., 2004; Proft and Struhl, 2002; Zapater et al., 2007). Most of these components are generally required for efficient stress-activated transcription, and mutants in the Rpd3, SAGA, and Mediator complexes are osmosensitive (de Nadal et al., 2004; Zapater et al., 2007). However, we do not know whether the recruitment of all transcriptional coactivator complexes is equally important for osmoinducible gene expression or whether some complexes are more important at specific subsets of stress genes. Interestingly, it has been found that although Mediator is more generally required for activated transcription upon osmostress, the function of SAGA is much more pronounced under severe stress conditions (Zapater et al., 2007). Therefore, the differential recruitment of basal transcription complexes might be a way of more dynamically adapt to different stress situations. The role of Hog1 in the control of activated expression is not restricted to transcriptional initiation, as it physically interacts with RNA pol II and crosslinks to whole transcribed regions upon stress (Pascual-Ahuir et al., 2006; Pokholok et al., 2006; Proft et al., 2006). One of its functions during transcriptional elongation is the recruitment of the RSC chromatin remodeling complex to osmoinducible ORFs to allow for efficient histone eviction during transcriptional elongation. RSC seems to be globally involved in osmostress activated transcription as rsc mutants are generally osmosensitive (Mas et al., 2009).
Other signaling compounds have been implicated in the transcriptional osmostress response in yeast. This is the case for the Sch9 protein kinase, which is a central regulator of nutrient responsive growth and longevity (Pascual-Ahuir and Proft, 2007a; Smets et al., 2010). Upon osmotic stress and similar to Hog1, Sch9 associates with specific stress-regulated promoters via the transcription factor Sko1 and contributes to induced gene expression together with the Hog1 MAP kinase (Pascual-Ahuir and Proft, 2007b). Loss of Sch9 function leads to osmosensitivity and to impaired transcription of stress genes. It remains to be determined what impact Sch9 has on the genomic level regarding osmostress activated gene expression.
The protein kinase A (PKA) signaling pathway additionally plays important roles in the transcriptional adaptation to hypertonic stress. Although the principle function of the PKA pathway has been described with the adaptation from respiratory to fermentative metabolism (Santangelo, 2006; Thevelein et al., 2000), PKA has direct implications in the regulation of gene expression upon stress. In general, high PKA activity under favorable growth conditions promotes cell growth, glycolysis, and progression through the cell cycle, while stress resistance is downregulated. PKA activity is directly linked to osmostress-activated gene expression by the Msn2 and Msn4 transcription factors (Martinez-Pastor et al., 1996; Schmitt and McEntee, 1996). Msn2/4 are negatively regulated by PKA phosphorylation and retained in the cytoplasma under normal conditions (Gorner et al., 1998, 2002). General stress conditions, including osmostress, lead to a release from this negative control, nuclear accumulation of Msn2/4, and the subsequent transcriptional activation of a subset of stress-responsive genes. Interestingly, the Sko1 transcription factor also appears to be under PKA control. PKA activity acts through specific phosphorylation sites in Sko1, promotes nuclear import of Sko1, and interferes with Sko1-regulated gene expression (Pascual-Ahuir et al., 2001; Proft et al., 2001). It is important to note that like Hog1 and Sch9, also the catalytic PKA subunits Tpk1 and Tpk2, have been found to associate with the chromatin of target genes, mainly at highly transcribed regions (Tpk1) and ribosomal gene promoters (Tpk2) (Pokholok et al., 2006). It remains to be determined, whether differential recruitment of Tpk proteins to the genome upon stress is a general manner of regulating gene expression.
Another signal transduction route that contributes to osmostress adaptation is the Ca2+/calmodulin–calcineurin pathway. Calcineurin is a heteromeric protein phosphatase with two redundant catalytic subunits, Cna1 and Cna2, in yeast. The calcineurin pathway is activated mainly by saline stress, and not generally by osmotic stress. Consequently, calcineurin mutants are hypersensitive to cation specific stresses (Hirata et al., 1995; Mendoza et al., 1994; Nakamura et al., 1993). Genomic profiling showed that calcineurin affects the expression of a large number of genes in yeast (Yoshimoto et al., 2002). Saline stress regulated gene expression is achieved by calcineurin-dependent dephosphorylation of the transcriptional activator Crz1 (Matheos et al., 1997; Mendizabal et al., 2001; Stathopoulos and Cyert, 1997). In this case, dephosphorylation of Crz1 by stress-activated calcineurin promotes the nuclear import of the activator and stimulated transcription from Crz1 bound promoters. Crz1 is oppositely regulated by PKA, which by phosphorylation inhibits its nuclear localization and transcriptional activation (Kafadar and Cyert, 2004).
Additional specific transcriptional activators such as Skn7 or Yap6 have to be considered to define the array of different TFs contributing to osmostress activated transcription. The Skn7 response regulator and transcription factor is regulated by upstream components of the HOG pathway (Ketela et al., 1998; Li et al., 1998). Its main physiological function, however, is clearly related to the adaptation to oxidative stress (Krems et al., 1996; Lee et al., 1999; Morgan et al., 1997). Yap6 is yet another transcriptional activator that adds to the yeast osmoadaptation. However, Yap6 has only been shown to regulate transcription upon salt stress when artificially overexpressed (Mendizabal et al., 1998). Thus, its physiological relevance to osmoadaptation remains to be shown.
During the adaptation to hyperosmotic stress, a large set of genes globally associated with cell growth and ribosomal biogenesis/function is transiently downregulated. This coordinated response reflects the stop of cell proliferation caused by the stress situation. Taken into account that the gene clusters negatively regulated at their mRNA levels contain many extremely highly transcribed genes, the overall impact of this repression is considerable. This is especially relevant for the ribosomal protein genes (RP), which account for up to 50% of the overall transcription events in a yeast cell under favorable growth conditions (Warner, 1999). Therefore, the mechanisms underlying the controlled shutdown of RP gene expression upon stress are fundamental for our understanding of the cellular stress adaptation. Although this phenomenon has not been studied specifically for osmostress, it occurs generally under many different stress conditions and the molecular details will be explained here. High RP gene expression is achieved by the binding of specific transcription factors Rap1, Fhl1, and Ifh1 to basically all RP gene promoters in the yeast genome (Lieb et al., 2001; Rudra et al., 2005; Schawalder et al., 2004; Wade et al., 2004). Although Rap1 and Fhl1 association is constitutive, Ifh1 binding to the RP promoters via Fhl1 is rapidly reversed upon various stress treatments. Therefore, the dynamic association of the Ifh1 protein seems to be the critical event for efficient RP gene transcription. Furthermore, RP gene expression and ribosome biogenesis is regulated by the TOR (target of rapamycin) and PKA signaling pathways (Cardenas et al., 1999; Neuman-Silberberg et al., 1995; Powers and Walter, 1999). Consequently, it has been shown that Ifh1 association at RP gene promoters is activated by TOR and PKA. On the other hand, conditions that slow down the growth rate and inhibit TOR and PKA activity, favor the nuclear import of the Crf1 repressor and its association with Fhl1 to shut off RP gene transcription (Martin et al., 2004).
We can summarize that transcriptional regulation upon osmostress involves a large number of specific TFs that modulate gene expression by various mechanisms (Fig. 3). In some cases like Sko1, the TF is constitutively bound to its target promoters and confers repression or activation dependent on the stress condition. In other cases like Hot1, Msn2, Msn4, or Crz1, TF binding to the genome almost exclusively occurs upon stress. We can anticipate a complex and dynamic scenario when we aim at resolving the transcriptional program at the genomic level.
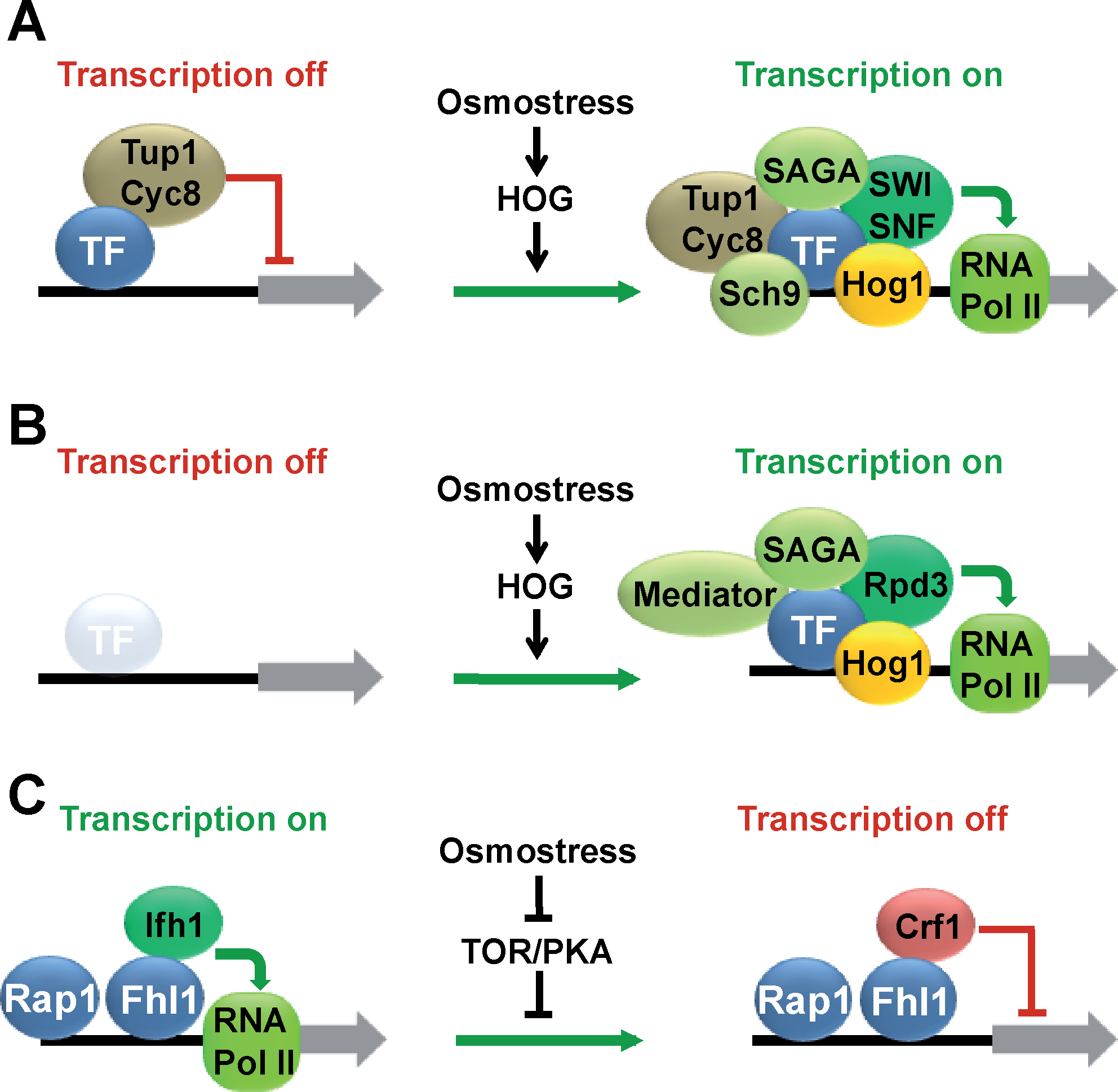
Mechanisms of transcriptional regulation upon osmotic stress. (
The Global Organization of the Transcriptional Program upon Osmostress: Learning from Genomic Location Analyses
Complex transcriptional responses have been successfully studied in yeast by the use of chromatin immunoprecipitation (ChIP) in combination with microarrays (ChIP–Chip) (Borneman et al., 2006; Simon et al., 2001). Because ChIP identifies the physical targets of transcription factors independently of the expression level, it provides the binding maps for all participating TFs, which finally can be correlated with the gene expression profiles observed under osmostress. Additionally, combining the global gene expression profiles from single and double mutant strains has been proven to efficiently dissect the complex transcriptional network operating upon osmotic stress (Capaldi et al., 2008). The genomic binding sites for the TFs primarily involved in the adaptation to osmotic stress (Sko1, Hot1, Smp1, Msn1, Msn2, Msn4, Yap4, Yap6, Skn7, and Crz1) have been mapped by ChIP (Harbison et al., 2004; Ni et al., 2009; Proft et al., 2005). A recent exhaustive study shows that it is critical to monitor TF binding in a dynamic manner along the osmotic shock to capture the complexity of the osmostress response (Ni et al., 2009).
There are several important conclusions that can be drawn from the genomic location analyses and global transcriptional profiling of TFs involved in osmoadaptation: (1) the binding of a given TF does not correlate with stress-induced gene expression in many cases. Although Sko1, Hot1, or Msn2 seem to preferentially bind to promoters of osmoinducible genes, other factors like Msn1, Yap4, or Yap6 associate with both induced and repressed genes (Capaldi et al., 2008; Ni et al., 2009; Proft et al., 2005). These data reveal an unanticipated complexity of TF binding and gene expression upon osmostress. (2) There is considerable overlap of the binding patterns of several TFs of the osmostress response, especially for Sko1, Msn2, Yap4, Yap6, and Skn7 (Ni et al., 2009). This indicates that there is a certain degree of redundancy in the transcriptional program upon osmostress and opens up the interesting possibility that specific expression patterns are achieved by specific combinations of TFs bound at the same promoter (Fig. 4). Partially redundant regulation of gene expression has also been identified by exhaustive transcriptional profiling of mutant strains, identifying Sko1, Hot1, Msn2, and Msn4 as the main contributors of Hog1 activated gene expression (Capaldi et al., 2008). (3) Different TFs show dynamic binding patterns during the osmostress response (Ni et al., 2009). The binding of a given TF along the stress is not uniform for all target genes. This observation adds to the complexity of the transcriptional response, as it might be that the dynamic formation of different TF combinations fine tunes the transcriptional output during the adaptation process. (4) The TFs involved in the osmostress response form a highly interconnected transcription network. Sko1 has been identified as one of the most upstream components in this network, which binds to the promoters of several other TFs directly involved in the osmostress response. All of the “core” TFs (Sko1, Hot1, Smp1, Yap4, Yap6, Skn7, Msn1, Msn2, and Msn4) seem to be extensively interconnected by binding to their gene promoters forming a hierarchical network (Ni et al., 2009). These network motifs are likely to have important consequences for the transcriptional output (Alon, 2007). For example, a feed-forward loop of several TFs could impose different timing or stress sensitivities to the expression of distinct target genes depending on the position of the TF within the regulatory cascade (Fig. 4). Additionally, the osmostress regulated TFs target many other TF encoding genes (Ni et al., 2009), and for most of them their implication in the stress response is not obvious. These data provide evidence for a very complex transcriptional network, which operates upon osmotic stress.
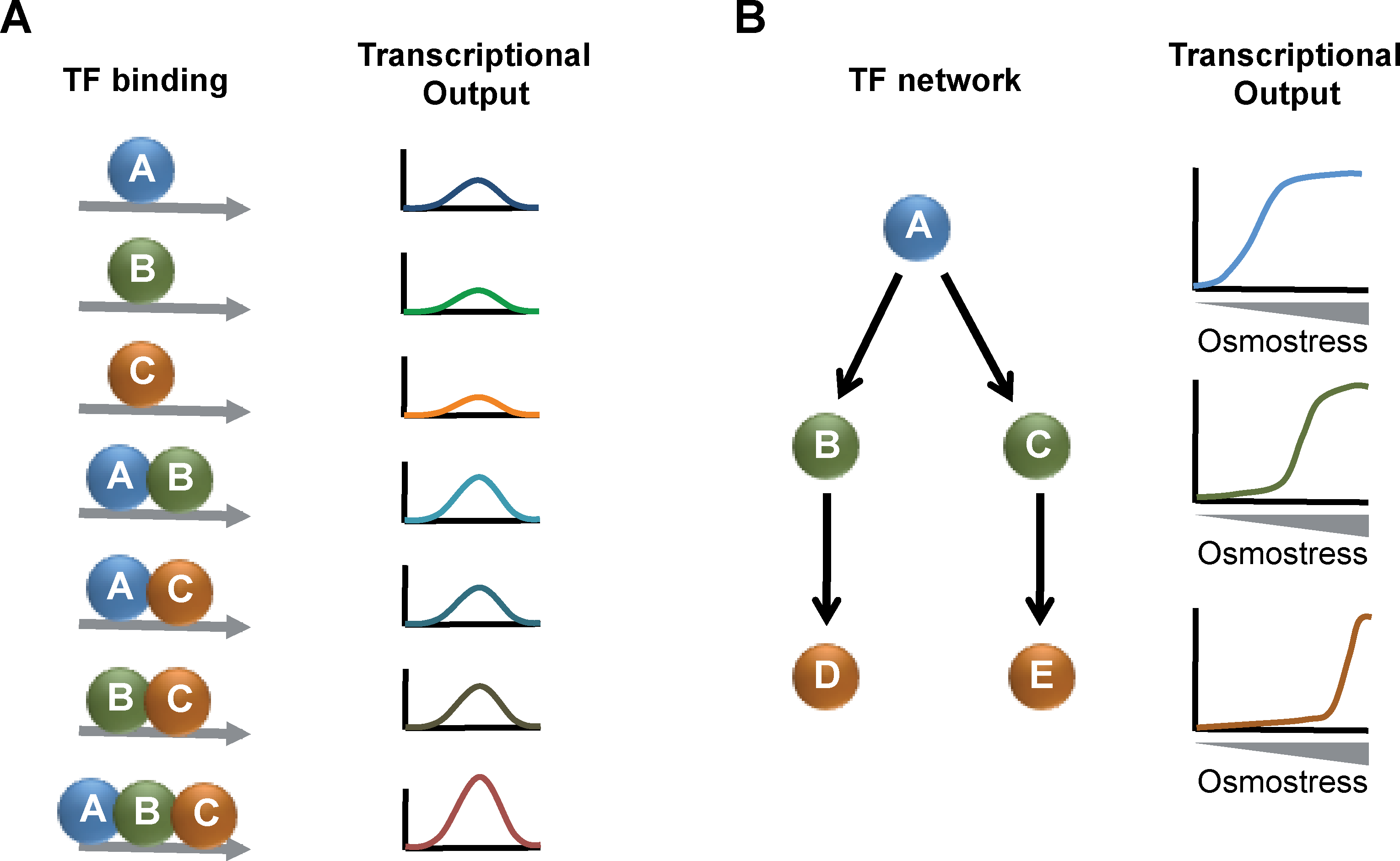
Possible regulatory functions of the transcription factor network operating upon osmostress. (
Regulation of mRNA Stability upon Osmotic Stress: Yet Another Layer of Global Control
The changes in gene expression patterns upon environmental stimuli are not only the result of differential rates of transcription. To understand how the cell dynamically adjusts its protein composition upon stress, we have to take into account additional regulation at the posttranscriptional level. Osmotic stress causes the transient inhibition of translation initiation during the first phase of stress exposure, which is then overcome dependent on the function of the Hog1 MAP kinase (Uesono and Toh, 2002). Therefore, the cell has to make sure that osmoinducible mRNAs are preferentially translated during the early phase, whereas osmorepressed mRNAs are retained from active translation. This has been shown at the genomic scale, where transcriptionally upregulated mRNAs are predominantly associated with ribosomes, whereas osmo-repressed mRNAs tend to accumulate in a nontranslating pool during stress (Melamed et al., 2008). The molecular mechanisms underlying this translational control are currently unknown.
Modulation of mRNA stability upon osmotic stress has been reported for specific yeast genes (Greatrix and van Vuuren, 2006; Hilgers et al., 2006). Moreover, two genomic studies have been performed very recently addressing the question how differential mRNA stability contributes to the changes in gene expression upon osmostress in yeast (Molin et al., 2009; Romero-Santacreu et al., 2009). Kinetic surveys of mRNA stabilities during osmostress show that the mRNA turnover is highly regulated and coordinated with the modulation of the transcription rates. Typical stress activated genes show a stabilization of their mRNAs during the initial response (Molin et al., 2009; Romero-Santacreu et al., 2009). Apparently, by enhancing transcription and mRNA stability the cell potentiates the expression of the activated stress defense genes. Interestingly, the same mRNAs are destabilized in the later recovery phase of osmostress adaptation, contributing in this way to the transient nature of the osmostress response. The mRNAs related to cell proliferation like the RP genes seem to be stabilized in the transition from the immediate shock phase to the recovery phase, presumably to allow for efficient growth after the immediate adaptation (Molin et al., 2009). These data show that the gene expression program upon osmostress is the result of extensive coordination of transcriptional activity and mRNA turnover. Further investigation will be needed to unravel the molecular mechanisms that are responsible for this coordinated regulation of mRNA stability upon osmotic stress.
Conclusions and Future Perspectives
The hyperosmotic stress response in yeast is an attractive model for many research groups, because the simple stress treatment triggers a complex and highly coordinated defense. The lessons learned with yeast are important to understand stress regulated gene expression in higher eukaryotes. This is particularly true for the evolutionarily conserved function of MAP kinases in stress-activated gene expression (Sheikh-Hamad and Gustin, 2004). For example, the Hog1 homolog p38 is responsible for large transcriptional reprogramming in mammalian cells (Ferreiro et al., 2010), and many mechanistic details about its function is derived from pioneering work in yeast (de Nadal and Posas, 2010). Specifically, mammalian p38 has direct functions in the assembly of active promoter complexes during developmental processes such as skeletal myogenesis and, similar to Hog1, targets distinct TFs and the SWI–SNF chromatin remodeling complex while intimately recruited to the chromatin of the regulated genes (Lluis et al., 2005; Rampalli et al., 2007; Serra et al., 2007; Simone et al., 2004).
Over the past years a very detailed knowledge has emerged that very well describes the signal transduction pathways, transcription factors, and molecular mechanisms underlying the gene specific transcriptional regulation upon osmostress in yeast. We have learned that the cell employs a complex regulatory network to ensure a wide reprogramming of gene expression. The combination of genome scale transcription profiling with global location analyses has raised a number of interesting models for the functions of this network, which can be experimentally addressed in the future: how do dynamic combinations of TFs create specific transcriptional readouts at different sets of osmoresponsive promoters? Do the different TFs depend on each other for their transcriptional activator/repressor function? How does the interconnection of the TFs establish different timing or stress thresholds to different classes of osmostress responsive genes? Also, the large number of TF encoding genes whose expression is targeted by osmostress needs further attention to address whether more TFs have to be implied in specific aspects of the osmostress response. An emerging research field is the regulation of mRNA turnover and its importance for osmostress adaptation. Here, future experimental approaches have to dissect the molecular mechanisms and the function of signal transduction pathways leading to differential mRNA stability upon osmotic stress.
Footnotes
Acknowledgments
We apologize to all colleagues whose work is not cited here because of space limitation. The work of our laboratory is supported by grants from Ministerio de Educación y Ciencia (BFU2005-0174), Ministerio de Ciencia e Innovación (BFU2008-00271), and Generalitat de Valencia (ACOMP07-198). F.M. is recipient of an FPI predoctoral fellowship from Ministerio de Educación y Ciencia.
Author Disclosure Statement
No competing financial interests exist.