
Abstract
Abstract
Resveratrol is a naturally occurring anticancer compound present in grapes and wine with antiproliferative properties against breast cancer cells and xenografts. Our objective was to investigate the metabolic alterations that characterize the effects of resveratrol in the human breast cancer cell lines MCF-7 and MDA-MB-231 using high-throughput liquid chromatography-based mass spectrometry. In both cell lines, growth inhibition was dose dependent and accompanied by substantial metabolic changes. For all 21 amino acids analyzed levels increased more than 100-fold at a resveratrol dose of 100 μM with far lower concentrations in MDA-MB-231 compared to MCF-7 cells. Among the biogenic amines and modified amino acids (n = 16) resveratrol increased the synthesis of serotonin, kynurenine, and spermindine in both cell lines up to 61-fold indicating that resveratrol strongly interacts with cellular biogenic amine metabolism. Among the eicosanoids and oxidized polyunsaturated fatty acids (n = 17) a pronounced increase in arachidonic acid and its metabolite 12S-HETE was observed in MDA-MB-231 and to a lesser extent in MCF-7 cells, indicating release from cell membrane phospholipids upon activation of phospholipase A2 and subsequent metabolism by 12-lipoxygenase. In conclusion, metabolomic analysis elucidated several small molecules as markers for the response of breast cancer cells to resveratrol.
Introduction
Although gene and protein expression in breast cancer cells after resveratrol treatment have been extensively profiled, there are no data about the metabolic alterations caused by this compound. In contrast to genetics and proteomics, the identification and quantification of specific metabolites in tumor cells provide high-resolution biochemical snapshots depicting the functional endpoints of the physiologic state of an organism, including the effects of drug disposure (Deberardinis et al., 2008; Weinberger and Graber 2005).
Studies conducted on laboratory animals and humans have reported a very low oral bioavailability of resveratrol based on extensive metabolism in gut and liver to several glucuronides and sulfates. In human breast cancer cell lines, however, resveratrol is exclusively metabolized to trans-resveratrol-3-O-sulfate. Surprisingly, in this setting the concentrations of resveratrol glucuronides were below the detection limits (Murias et al., 2008). Furthermore, recent data from our lab also demonstrate that trans-resveratrol-3-O-sulfate was about threefold less cytotoxic against the hormone-dependent MCF-7 and the hormone-independent MDA-MB-231 human breast cancer cell lines with IC50 values of about 200 μM, indicating that sulfation of resveratrol has only a minor effect on cell growth inhibition (Miksits et al. 2010). Therefore, we used these cell lines to investigate possible alterations in the cellular concentrations of amino acids, biogenic amines, eicosanoids, and polyunsaturated fatty acids after resveratrol application using a targeted metabolomic approach. This information is important as some small molecules analyzed in this study may act as markers for the anticancer activity of resveratrol.
Materials and Methods
Materials
Resveratrol (3,4′,5-trihydroxy-trans-stilbene, 99% GC) and dimethyl sulfoxide (DMSO) were obtained from Sigma-Aldrich (Munich, Germany). All other chemicals and solvents were commercially available, of analytical grade, and used without further purification.
Cell culture
MCF-7 and MDA-MB-231 breast cancer cells were purchased from the American Type Culture Collection (ATCC, Rockville, MD, USA). Both cell lines were grown in phenolred-free RPMI 1640 tissue culture medium including L-glutamine (PAN Biotech, Aldenbach, Germany), supplemented with 10% heat-inactivated fetal bovine serum (FBS) and 1% penicillin-streptomycin (Gibco Invitrogen Corp., Grand Island, NY, USA) under standard conditions at 37°C in a humidified atmosphere containing 5% CO2 and 95% air. Twenty-four hours before treatment, cells were transferred to a RPMI 1640 medium supplemented with 2.5% charcoal-stripped FBS (PAN Biotech, Aidenbach, Germany) and 1% penicillin–streptomycin. Cells were placed into 15-cm plates and allowed to attach overnight. Resveratrol was dissolved in DMSO and diluted with medium (final DMSO concentration <0.1%) to 5–100 μM. Experiments under each set of conditions were carried out in triplicate. Blank experiments contained DMSO in the medium in place of resveratrol. After 72 h, media were aspirated by suction and aliquots (100 μL) were analyzed by LC-MS/MS. In parallel, cells were scraped off, washed three times with phosphate-buffered saline, and lysed in ethanol/phosphate buffer (85/15 v/v) by repeating (three times) shock freezing in liquid nitrogen, and thawing. After centrifugation at 10,000 × g for 5 min, 10 or 20 μL of the supernatant (cytoplasm) was subjected to the LC-MS/MS quantification assays.
Cell growth inhibition
The effect of resveratrol (0–100 μM) on the in vitro growth of MCF-7 and MDA-MB-231 cells was evaluated after 72 h of resveratrol application under identical conditions (see above) using the CellTiter-Glo® Luminescent Cell Viability Assay (Promega, Madison, WI, USA) and a Victor™ microplate reader (Perkin-Elmer, Waltham, MA, USA) according to the manufacturer's instructions.
Targeted metabolomics
Using a high-throughput liquid chromatography-based mass spectrometry platform for targeted metabolomics, 54 analytes were quantified in cell pellets and in medium at Biocrates Life Sciences AG, Austria. Multiple reaction monitoring detection was performed using a 4000 Q TRAP tandem mass spectrometry instrument (Applied Biosystems, Bedford, MA, USA) to obtain concentration data, which were finally exported for statistical analysis. Metabolomics data were used as received from Biocrates. No data correction or removal of data points was applied. The experimental metabolomics measurement technique was carried out as previously described (Gieger et al., 2008).
Statistical analysis
Unless otherwise indicated, values are expressed as mean ±SD of three individual experiments. Statistical differences from control values were evaluated using the Students' paired t-test at a significance level of p < 0.05 using the Prism program (version 5.0, GraphPad Software Inc., San Diego, CA, USA).
Results
Amino Acids
Resveratrol significantly reduced cell viability in the cancer cell lines MCF-7 and MDA-MB-231, yielding IC50 values of 68.3 ± 2.6 and 67.6 ± 4.1 μM, respectively (data not shown). Cell growth inhibition was accompanied by substantial metabolic changes, which were dose dependent but different between both cell lines. After 72 h of cell growth in the presence of resveratrol, the concentrations in the medium of all 21 analyzed amino acids (19 proteinogenic, 2 nonproteinogenic) were substantially increased compared to resveratrol-free controls. For MDA-MB-231 cells, this effect was less pronounced than for the MCF-7 cell line (data not shown). In the presence of resveratrol, the maximum changes seen between resveratrol-treated cells and controls were 21-fold for serine in MCF-7 cell culture and 63-fold for methionine in MDA-MB-231 cells (Table 1). Significant increases in the synthesis of all amino acids under resveratrol treatment was also observed in the cytoplasm of MDA-MB-231 cells (up to 18-fold), whereas the concentrations of many amino acids, most notably aspartic acid, glutamine, glycine, and ornithine in MCF-7 cells were decreased (0.42 to 0.56-fold) (Table 1).
Values in bold indicate significant changes (p < 0.05).
n.d., not detectable.
Biogenic Amines and Modified Amino Acids
Metabolic changes in response to resveratrol were also seen for biogenic amines and modified amino acids (n = 16). As already observed for amino acids, much higher concentrations were seen in the medium of MCF-7 cells than in the MDA-MB-231 cell line. In cytoplasm, however, concentrations in both cell lines were very low or below the detection limit (Table 1). Most important, resveratrol significantly increased the synthesis of serotonin, kynurenine, spermidine, and spermine by up to fivefold in MCF-7 and up to 61-fold in MDA-MB 231 compared with controls (Fig. 1), indicating that resveratrol strongly interacts with cellular biogenic amine metabolism. Furthermore, resveratrol induced the oxidation of methionine to methionine sulfoxide by 1.7- and 5.24-fold in MCF-7 and MDA-MB-231 cells, respectively (Table 1). Phenylalanine and phenylethylamine (PEA) concentrations were below the detection limits in both cell lines.
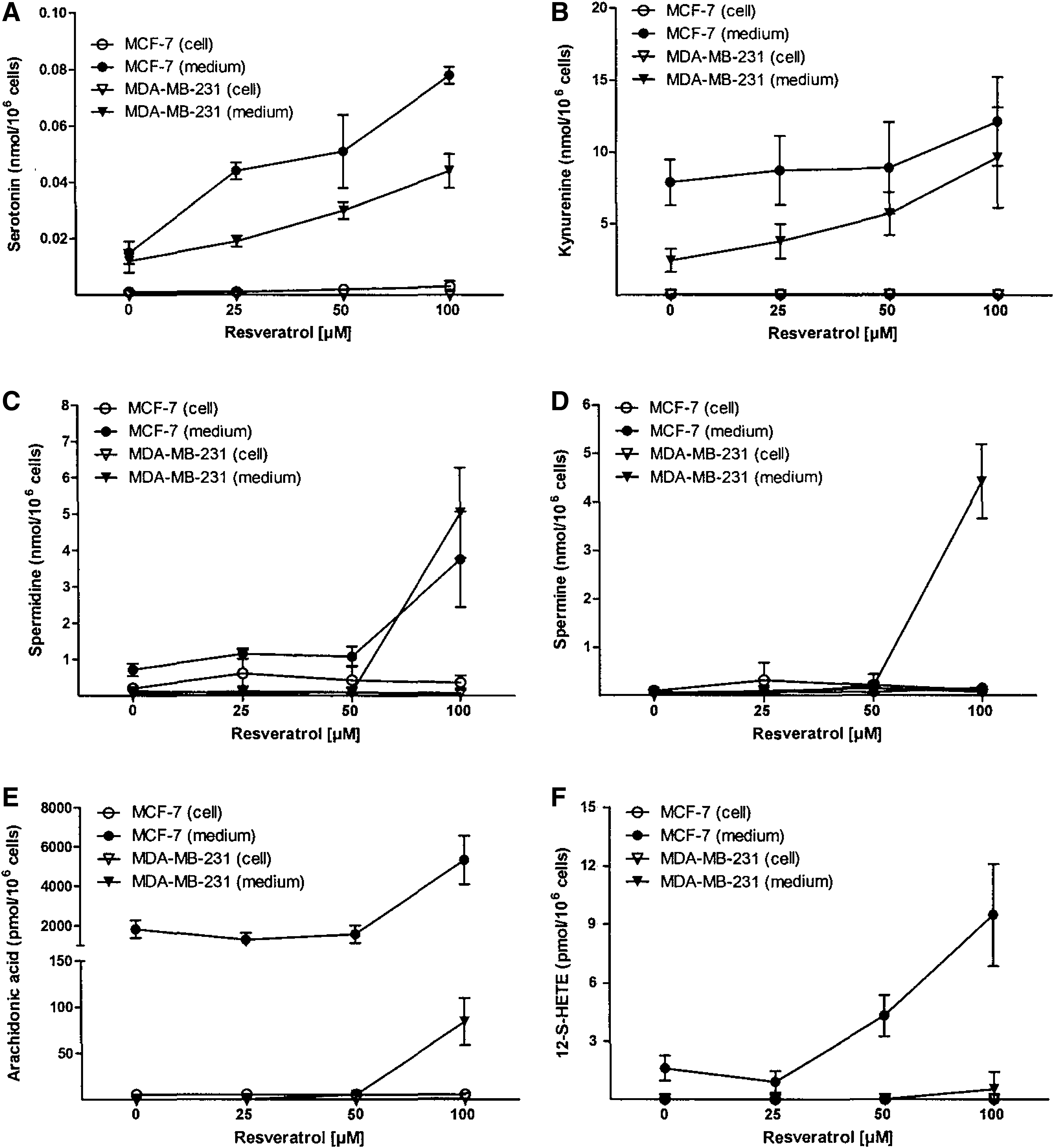
Induction of serotonin (
Eicosanoids and Oxidized Polyunsaturated Fatty Acids
Among the 17 analytes quantified, a marked increase in extracellular arachidonic acid and its metabolite 12S-HETE (12(S)-hydroxy-5Z,8Z,10,E14Z-eicosatetraenoic acid) was observed (Fig. 1). Concentrations of the linoleic acids metabolites 13S-HODE [13(S)-hydroxy- 9Z, 11E-octadecadienoic acid] and 9-HODE [(±)9-hydroxy-10E,12Z-octadecadienoic acid] were also increased by resveratrol. Remarkably, extracellular arachidonic acid concentrations rose 84-fold in MDA-MB-231 cells cultures, but only 2.9-fold in MCF-7 cells compared to control (Fig. 1). Also, resveratrol significantly reduced prostaglandin E2 (PGE2) levels in the medium of MDA-MB-231 cells (>99%), whereas the reduction in MCF-7 cells was less pronounced (65%) (Table 1). Several other oxidized polyunsaturated fatty acids and prostaglandins as well as leucotriene D4 and thromboxane B2 were below the detection limit.
Discussion
In the present study, we investigated the metabolic changes in two human breast cancer cell lines after resveratrol application (5–100 μM). These concentrations were chosen based on daily intake of resveratrol as beverage (red wine) or as dietary supplement (5–100 mg/day). By quantifying 54 analytes, we found that resveratrol significantly induced the synthesis of 21 amino acids with far higher concentrations in MCF-7 than in MDA-MB-231 cells. In both cell lines, all amino acids were substantially released from the cytoplasm into the medium, which is often caused by cell swelling and the occurrence of reactive oxygen species (Lambert, 2007). Resveratrol also profoundly modulated the polyamine biosynthesis in both cell lines. Tryptophan, serotonin, and kynurenine increased significantly in the presence of resveratrol, indicating that enzymatic conversion of tryptophan to the bioactive metabolite serotonin through tryptophanhydroxylase and to kynurenine through tryptophan-2,3-dioxygenase and monooxygenase was stimulated. Kynurenine was further metabolized to hydroxykynurenine3-hydroxy-kynurenine with much higher concentrations in the medium of MDA-MB-231 cells than in the MCF-7 cell line (Fig. 1).
Treatment of both breast cancer cell lines with resveratrol also stimulated the synthesis of putrescine and spermidine indicating activation of ornithine decarboxylase and spermidine synthase, respectively. Interestingly, synthesis of spermine from spermidine was stimulated in MDA-MB-231 cells but inhibited in MCF-7 cells. Because putrescine, spermidine and spermine are essential for a variety of cellular processes related to signal transduction, resveratrol-induced growth and differentiation changes in polyamine metabolism may be directly linked to cell vitality (Takao et al., 2006). Conversion of putrescine to the metabolically active polyamines spermidine and spermine occurs early during cell proliferation. It is mediated by S-adenosylmethionine decarboxylase (SADMC), the rate-limiting enzymes of polyamine biosynthesis. Similar to ornithine decarboxylase (ODC), SADMC activity is increased in proliferating cells (Milovic et al, 2000). In human colon adenocarcinoma CaCo-2 cells, resveratrol, and the analog (Z)-3,5,4′-trimethoxystilbene have been shown to reduce ODC and SADMC activities by depletion of the polyamines putrescine and spermidine, exerting their cytotoxic effects by depleting the intracellular pool of polyamines (Schneider et al., 2003; Wolter et al., 2003). In contrast to colon cancer cells, resveratrol stimulated putrescine and spermidine synthesis in MCF-7 and MDA-MB-231 cells, indicating that cell growth inhibition may rather be caused by high polyamine concentrations, which have also been to induce cell death (Takao et al., 2006).
Our study also showed a pronounced increase in extracellular arachidonic acid and its metabolite 12S-HETE at high resveratrol concentrations, indicating the release of arachidonic acid from cell membrane phospholipids upon activation of phospholipase A2. Arachidonic acid is subsequently converted to 12S-HETE through the action of 12-lipoxygenase. Increased levels of 12S-HETE may therefore indicate oxidative stress in tumor cells under resveratrol treatment (Nazarewicz et al, 2007). Furthermore, resveratrol also reduced prostaglandin E2 (PGE2) levels, thus confirming that this polyphenol is an inhibitor of cyclooxygenase 2 (Murias et al. 2004). In conclusion, we revealed several small molecules as novel markers for the anticancer activity of resveratrol. Further investigations are required to better understand the resveratrol-induced metabolic differences between hormone-sensitive and hormone-insensitive cell lines.
Footnotes
Acknowledgments
This study was supported by grants of the Jubiläumsfonds der Österreichischen Nationalbank (12600 to W.J.) and FWF (P21083-B11 to W.J.).
Author Disclosure Statement
No competing financial interests exist.