
Abstract
Abstract
Identification of more and more novel tumor antigens and autoantibodies will lead to the earlier diagnosis, better prognosis prediction, and more efficient therapy of cancer in the future. Immunoproteomics techniques have successfully been used for finding novel cancer biomarkers in different subgroups of cancer patients. HER2 is a marker for an aggressive breast cancer, particularly in node-positive (NP) cases. The aim of our study was to identify antigens eliciting a humoral immune response in HER2+ and HER2− NP breast cancers by two-dimensional electrophoresis (2D), Western blotting, and mass spectrometry. Sera from 18 women with newly diagnosed NP breast cancer (9 HER2+ and 9 HER2−) and 9 healthy volunteers were individually investigated for the presence of antibodies to MCF7 breast cancer cell line proteome. Reactive spots in 2D blots were matched to stained 2D gels. Twenty-eight of matched spots were identified by mass spectrometry. Among them were LDH-A, glyceraldehydes-3-phosphate dehydrogenase, enolase-α, phosphoglycerate dehydrogenase, proteasome 26S non-ATPase subunit 13, triosephosphate isomerase, hnRNP K, hsp27, hsp90, prohibitin, nucleophosmin, 14-3-3ɛ, PP2A regulatory subunit, and ribonuclease inhibitor-angiogenin. The five former antigens were more commonly reacted with sera from HER2+ cases, and the three latter antigens were more commonly reacted with sera from HER2− cases. Noteworthy, the antigenicity of the 28 spots showed a few differences when SBR3 cell line was used as the source of antigens. Although some of the identified antigens were previously defined as tumor antigens, others were novel. Further investigations for their utilizations as markers for breast cancer diagnosis, progression, and therapy are warranted.
Introduction
The talent of cancer cells for the aberrant expression of a large number of proteins (e.g., overexpressed, mutated, or improperly posttranslationally modified proteins) is a substantial rationale for the heterogeneity of the breast cancers (Kuramitsu and Nakamura, 2006). The aberrant expressions of estrogen receptor (ER) and HER2 are commonly used as biomarkers for breast cancer typing, prognosis, and targeted anti-ER or anti-HER2 therapies. However, the existence of a wide variation in prognosis and response to therapy among these breast cancer types stands for other aberrantly expressed proteins with significant roles on breast cancer outcome that are not well recognized (Gonzalez-Angulo et al., 2007).
The aberrant expression of proteins elicits the immune system for autoantibody production, and therefore the serum autoantibody profiles are different between healthy individuals and cancer patients (Hamrita et al., 2008; Tamesa et al., 2009). The presence of an autoantibody in cancer may be an indicator of a population of T cells against an amenable antigen for generating an anticancer immune response. Evidence suggests that boosting the immune response against these antigens generate an effective immune response to cancer (Philip et al., 2007). The hallmark of aberrantly expressed antigenic proteins is HER2, an oncoprotein that its targeting by specific inhibitory ligands, and also by boosting the immune response using HER2 vaccination has clinical application in HER2 overexpressing patients (Benavides et al., 2009).
Serological proteome analysis (SERPA), also called immunoproteomics and two-dimensional (2D) Western blot, is a common technique for the high throughput identification of tumor antigens (Hamrita et al., 2008). In this technique, proteins in tumor cell lysates are first separated by 2D gels, and then transferred onto membranes that are probed with sera from cancer patients or controls. Proteins reacting with antibodies are subsequently identified by mass spectrometry (MS). By the 2D Western blotting approach, which efficiently distinguishes posttranslationally modified proteins, or other immunoproteomics approaches, such as serological screening of tumor-derived cDNA expression libraries and phage display libraries, several putative biomarkers in various types of cancer have been uncovered (Chen et al., 2008; Hamrita et al., 2008). Using 7-cm medium pH-range (pH 4–8) strips in the first dimension of 2D Western blotting, several novel breast cancer antigens such as heterogeneous nuclear ribonucleoproteins (hnRNP K), prohibitin, and peroxiredoxin-2 were identified on the whole of breast cancer patients (Hamrita et al., 2008). However, by traditional 1D electrophoresis, it has been shown that the pattern of serum immunoreactivity against breast cancer cell lysates have differences along with the severity of the disease (Hamsher et al., 2007). HER2 overexpression and antigenicity occur in roughly 20% of breast cancer patients, and is associated with an aggressive disease and poor prognosis, particularly in axillary lymph node positive (NP) patients (Purdie et al., 2010). Although there are numerous proteomics studies regarding differentially expressed proteins between HER2+ and HER2− cancer cells in general (Zhang et al., 2005), the information regarding differentially expressed proteins or antigens between these two subsets in NP breast cancers is poor.
In the present study, 2D blots of beast cancer cell lines (using 18-cm wide pH-range (pH 3–10 nonlinear) strips in the first dimensions) were probed with individual sera from HER2+ and HER2– NP breast cancer patients and healthy individuals. Antigenic proteins were then identified by matrix-assisted laser desorption ionization-time of flight/time of flight (MALDI-TOF/TOF) MS. By this approach, differential immunogenic proteins between (1) the overall NP breast cancer patients and healthy individuals, and (2) HER2+ and HER2− NP cases were identified. At least, from a theoretical point of view, predominant antigens in a poor prognosis stage compared to a better prognosis stage (such as HER2+ vs. HER2− NP breast cancers) might be more aberrantly expressed and more culprit molecules in favor of cancer progression. Identification of such antigens can be the first step in better understanding of tumor immunology, biology, subtyping, prognosis, and designing targeted therapy by means of specific ligands or by efficient immunization.
Materials and Methods
This study was approved by the Ethics Committee of Shiraz University of Medical Sciences. All participants were informed that blood samples will be used for research projects, and their consent was obtained.
The breast cancer patients were recruited from Motahari clinic in Shiraz, Iran, and blood was taken prior to surgery or any other treatment. No patients had a history of an autoimmune disease. The mean age at onset of the disease was 47.6 ± 9.4 years, ranging from 35 to 60 years. Infiltrating ductal carcinoma and the involvement of more than two axilliary lymph nodes were confirmed histopathologically in all the patients after the surgery. Patients were at stage III disease with no detectable distant metastasis at diagnosis. The status of HER2 and ER were checked by immunohistochemistry (IHC) technique. Nine HER2+ and nine HER2− NP breast cancer patients were included in the study. Eight out of nine of HER2− and five out of nine of HER2+ patients were estrogen receptor (ER) positive. Using Hercep test, tumors were scored 0, 1+, 2+, and 3+. Only scores of 3+ were included in our HER2+ group.
The control group consisted of nine healthy volunteers (mean age 47.6 ± 9.2 years), confirmed by physical exam and mammography, residing in the same region as patients. They had no history of cancer or autoimmunity. Serum from each participant was aliquoted and stored at −20°C until required.
Cell culture
The MCF7 (an ER+, low-expressing HER2 cell line) and SKBR3 (an ER−, high expressing HER2 cell line) breast cancer cell lines were purchased from the Pasteur Institute of Iran. The cells were cultured in 150 cm2 flasks under standard conditions. The culture media contained RPMI 1640 medium, 10% fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin. The cells were grown to confluence before harvesting. Cell culture media and reagents were obtained from Sigma-Aldrich (Germany).
Protein preparation
The cultured cells were harvested by trypsinization, washed three times with cold phosphate-buffered saline (PBS), and lysed in a lysis buffer containing 7 M urea, 2 M thiourea (Merk, Germany), 4% CHAPS, 2% IPG buffer 3–10, and 40 mM dithiothreitol (DTT). The cell lysates were incubated at room temperature for 2 h, and then centrifuged. The supernatants were collected and stored at −70°C. Protein concentration was determined using the 2D Quantitation kit. All the chemicals, except where stated, were purchased from GE Healthcare (Sweden).
2D Western blotting
Isoelectric focusing (IEF) was carried out using a Protean IEF Cell system (Bio-Rad, Hercules, CA, USA). Approximately 120 μg (for 2D Western blotting runs) or 500 μg (for gel-staining runs) of protein extracts were mixed with rehydration solution contained 7 M urea, 2 M thiourea, 2% CHAPS, 20 mM DTT, and 0.5% IPG buffer 3–10 nonlinear (NL). The mixtures were loaded on IPG strips (18 cm: pH 3–10 NL; GE Healthcare) by in gel rehydration for 13 h at 50 V. The loaded IPG strips were focused at 70–75,000 Vh. Once IEF was completed, the focused strips were equilibrated for 15 min in an equilibration buffer (50 mM Tris-HCl (GE Healthcare), pH 8.8, 6 M urea, 30% glycerol, 2% SDS (GE Healthcare) containing 65 mM DTT. This was followed by further 15 min incubation in the equilibration buffer containing 135 mM iodoacetamide (GE Healthcare).
The equilibrated strips were placed on the top of a SDS polyacrylamide gel, and embedded with 0.5% agarose (GE Healthcare). The second dimension SDS-PAGE (12%) was performed at a constant current of 20 mA/gel with Protean II xi 2D cell system (Bio-Rad), until the tracking dye reached the bottom of gels. A low-molecular SDS-PAGE weight marker (Fermentas, Lithuania) was used in the second dimension as molecular mass reference. Proteins were visualized by a modified Coomassie Brilliant Blue (CBB) staining method named blue silver (Candiano et al., 2004). Gels were scanned using a GS-800 scanner (BioRad) at 300 dpi resolution.
For 2D Western blotting, proteins from 2D gels derived from either MCF7 or SKBR3 lysates were transferred onto PVDF membranes by a semidry blotter (BioRad) for 1.5 h at 25 V. The membranes were then blocked with 5% skimmed milk (Fluka, Milwaukee, WI, USA) in 0.05% PBS-Tween (PBS-T) for 2 h and washed two times with PBS-T for 20 min. The blocked membranes were incubated overnight at 4°C with individual sera diluted 1/300 in PBS-T. Some membranes were probed with pooled sera from nine HER2+, nine HER2−, or nine controls. The blots derived from MCF7 lysates were probed with all studied serum samples and pooled sera, but blots derived from SKBR3 lysates were probed with individual sera from four persons in each group. After incubation with primary antibody, membranes were washed four times each 5 min. The washed membranes were incubated for 1.5 h with diluted (1/4,000) secondary antibody (antihuman, IgG conjugated with horseradish peroxidase; Sigma) in PBS-T. Again, membranes were washed five times, each time for 5 min. To visualize antigenic components, membranes were incubated in diaminobenzeden and H2O2 for 15 min. MCF7 blots were repeated twice.
Blots were analyzed using the Prodigy SameSpots version 1.0 software (Nonlinear Dynamics, UK). Briefly, MCF7 blot images were initially aligned by placing 21 manual vectors. Using these vectors as starting points, the software performed alignments by generating the automatic vectors for each image and showed them in the alignment review stage where the user can correct misalignments. To view the results according to group, the blot images derived from MCF7 cell line were set up in three separate groups (HER2+, HER2−, and control groups). Spots that present just in one blot were deleted in the “View Results” stage. After matching the blots, images of CBB stained gels and blots derived from SKBR3 lysates were added, and by selecting “use existing spot detection” bottom, the detected spots in the MCF7 blots were matched to the gels and SKBR3 blots.
The MCF7 and SKBR3 gels were also compared regarding to the expression of the detected spots by the Prodigy software. Spots that were differentially expressed more than twofold between MCF7 and SKBR3 groups (three gels in each group) were considered as differentially expressed protein spots.
Mass spectrometry
Immunoreactive protein spots were manually cut from 2D gels derived from MCF7 lysates, and sent in a 1.5-mL vial for MALDI-TOF/TOF MS analysis to the United Kingdom (Department of Biology, The Proteomics & Analytical Biochemistry Labs, University of York, UK). The peptide mass fingerprinting (PMF) and tandem mass spectrometry (MS/MS) information were searched against the National Center for Biotechnology Information nonredundant (NCBInr) database, using the Mascot search engine (Matrix Science, UK). Mass tolerance settings of 250 ppm for parent ion and 0.5 Da for fragment ions were applied. One missed cleavage per peptide was allowed. Carbamidomethylation of cysteine and oxidation of methionine residues were set as fixed and variable modifications, respectively. Peptide scores greater than 37 were considered as significant (p < 0.05).
Results
2D blots of MCF7 lysates were screened with individual sera from nine healthy individuals and 18 newly diagnosed NP breast cancer patients (HER2+ and HER2− subgroups), and with three pooled sera from HER2+, HER2−, and control groups. Each immunoblot was repeated two times with the same result. After blotting, the membranes were stained with Ponceau S for watching the blot efficiency. The blotted gels were also silver stained for watching the reproducible patterns of proteins in each blot. It is worth mentioning that after blotting, a minute amount of proteins still remained in the gels, especially in the high molecular weight regions. The reactive spots in MCF7 blots were matched to each other and also to the MCF7 CBB-stained gels using Prodigy software. Five 2D CBB stained gels with proteins obtained from independent experiments were run for the MCF7 cell line, and the patterns of proteins were found to be highly reproducible. Thirty-eight protein spots that reacted with at least two patients' sera, and generally with minimal reactivity with controls' sera, were sent for MS analysis. Of them, 28 spots were identified by a significant score. The locations of these 28 spots are shown in Figure 1. For verification of our protein identification, prior to identification of proteins, the spots 1, 2–4, 7, 9–12, 21, 23, and 26 were matched to the previously published MCF7 proteome (Ou et al., 2008). Our MS analyses identified the same proteins. We assumed that spot 2 must be a mixture of peroxiredoxin-6 and hsp27. Interestingly, our MS analysis identified spot 2 as the mixture of these two proteins. As a sample, the MS/MS data of enolase (spot 3) is indicated in Supplementary File 1.
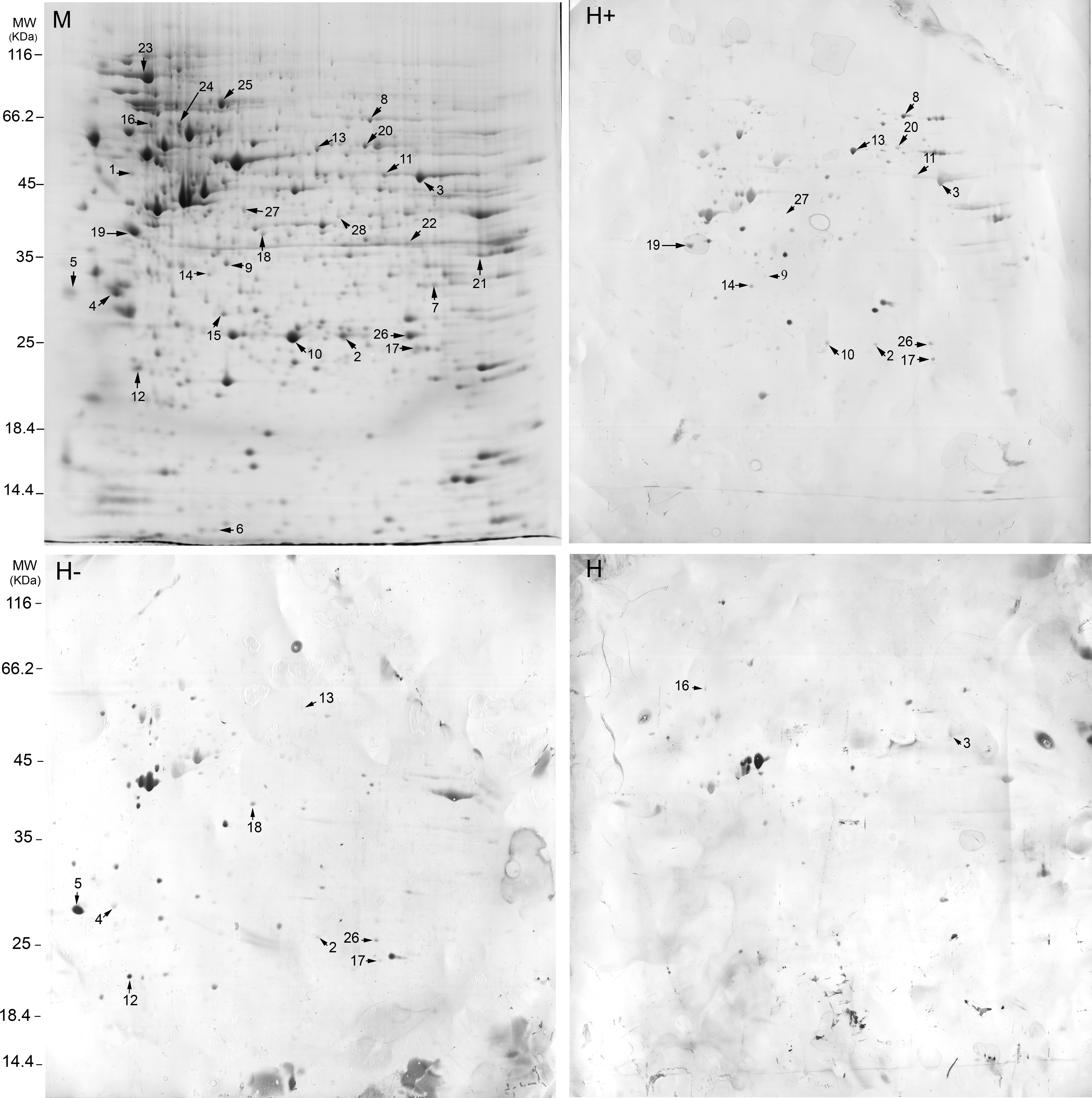
Representatives of MCF7 breast cancer cell proteins separated by 2DE using IPG strips PH 3–10 nonlinear that Coomassie stained (M), or probed with sera from HER2+ breast cancer patients (H+) (patient 5 in HER2+ group in Table 2), HER2− breast cancer patients (H−) (patient 4 in the HER2− group in Table 2), and healthy controls (H) and visualized by diaminobenzeden. Immunoreactive spots that are identified by MS are numbered. Spot numbers are related to Figures 2 and 3 and Tables 1 and 2.
The descriptions of the 28 identified spots were displayed in Table 1. All HER2− patients, except patient 6, were ER+. In the HER2+ group, patients 1, 5, 7, 8, and 9 were ER+. Others were ER− (Table 2). The profiles of antigenic proteins in patients were not the same. The profile of identified antigens in each patient is shown in Table 2. Spot 3 was partly visible on some blots probed with controls' sera. Others had no visibility or represented as faint spots with controls' sera. Stress-induced phosphoprotein 1 (STI1), pyrophsphatase 1, lactate dehydrogenase A (LDHA), glyceraldehyde-3-phosphate dehydrogenase (GAPDH), phosphoglycerate dehydrogenase (PHGDH), and proteasome 26S non-ATPase subunit 13 isoform were more commonly observed in blots of HER2+ cases compared to HER2− cases (>2), while 14-3-3ɛ, protein phosphates 2A regulatory subunit (PP2AR) and ribonuclease inhibitor-angiogenin were more common immunoreactive in HER2− cases (>2). The ratio of the immunoreactivity of other spots in two groups of our patients did not exceed 2 (ratio ≤2). Spots 3 and 11 had a marked increase in intensity in HER2+ cases, despite the fact that the numbers of patients in each group did not differ remarkably (Supplementary File 2). Enlarged sections of STI1, PHGDH, proteasome 26S non-ATPase subunit 13, LDHA, ribonuclease inhibitor-angiogenin, PP2AR, enolase (spot 3), enolase (spot 11), isocitrate dehydrogenase-3 (IDH3), nucleophosmin, eukaryotic translation initiation factor-3 (eIF3) in each person in one replicate of MCF7 blots were indicated in Supplementary File 2. These closed up sections have been obtained from the Prodigy software that calibrates images according to the reference.
National Center for Biotechnology Information nonredundant (NCBInr).
exp, experimental; pre, predicted.
2D blots of MCF7 lysates were also reacted with pooled sera from HER2+ and HER2− breast cancer patients and healthy individuals to realize the main features of antigens. Although spots 6, 7, 9, 10, 13–15, 17, 18, 20, 23, 25, and 27 were invisible, LDHA, GAPDH, STI1, enolase (spot 11), enolase (spot 3), translationally controlled tumor protein (TCTP), nucleophosmin, hnRNPK, hsp27 (spot 2), eIF3, triosephosphate isomerase (TPI) (Fig. 2), acidic leucine-rich nuclear phosphoprotein 32 (pp32), 14-3-3ɛ, PP2AR, and ribonuclease inhibitor-angiogenin were visible. The first four antigens were just observed in blots probed with sera from HER2+ cases (Fig. 2), while the last four antigens were observed in blots probed with sera from HER2− cases. Among the 28 spots, enolase (spot 3) was the only one that was visible, although very faint, on blots probed with sera from healthy individuals (Fig. 2).

Enlarged sections of MCF7 2D Western blots probed with pooled sera from healthy individual (H), HER2− (H−), and HER2+ (H+) breast cancer cases. Glyceraldehyde-3-phosphate dehydrogenase (spot 22), lactate dehydrogenase A (spot 21), stress-induced phosphoprotein 1 (spot 8), enolase in spot 11 are visible just in blots probed with sera from HER2+ cases, whereas enolase in spot 3 and triosephosphate isomerase (spot 26) are visible in blots of both group of patients. Enolase in spot 3 is visible in blots probed with sera from healthy individuals.
Sera from four HER2+ (patients 1, 2, 4, and 8 in Table 2) and four HER2− patients (patients 1, 2, 5, and 7 in Table 2) were reacted against 2D blots derived from SKBR3 cell lysates to recognize the possible different antigenic patterns between different cell lines with different levels of HER2 and ER expressions. The 28 identified antigenic spots were matched to the equal positions on 2D immunoblots from SKBR3 lysates. We observed invisibility of electron transfer flavoprotein α subunit (ETFA) (spot 7) (patient 2 in the HER2− group) in SKBR3 blots. Consistently, this spot was not visible in SKBR3 gels (Fig. 3). Among the 28 spots, nucleophosmin (spot 19) was also overexpressed (more than twofold) in MCF7 gels compared to SKBR3 gels, but LDHA and pyrophosphatase in SKBR3 gels. Their visibilities were somewhat different between MCF7 and SKBR3 blots. As a sample, the comparison of nucleophosmin (patients 2 and 5 in the HER2− group and patient 8 in the HER2+ group in Table 2) between MCF7 and SKBR3 blots is shown in Figure 3.

The reactivities of electron transfer flavoprotein α subunit (ETFA) (spot 7) and nucleophosmin (spot 19) with sera in MCF7 blots (M-blot) and SKBR3 blots (S-blot). These spots were overexpressed (more than twofold) in MCF7 gels (M-gel) compared to SKBR3 gels (S-gel). p refers to the patient number in Table 2. P2 and p5 are in HER2− group and p8 is in HER2+ group.
Discussion
Breast cancers are heterogeneous even in an individual patient in different disease stages, which are partly attributed to the ability of cancer cells for the employment and aberrant expression of a large number of key molecules involved in tumorigenesis (Gonzalez-Angulo et al., 2007; Kuramitsu and Nakamura, 2006). The aberrant expressions of proteins stimulate the immune system for the antibody production. There is a consensus that the composition of autoantibody profile in each patient might be a clue for cancer individualized therapy (Philip et al., 2007). 2D Western blotting seems to be the easiest method for globally finding antigens in each patient. By this method, we identified 28 antigenic spots, generally with a low reactivity with controls' sera. Patients groups were comprised of HER2+ and HER2− NP breast cancer cases. HER2, an overexpressed antigen in roughly 20% of breast cancer patients, is a marker for response to targeted therapy by anti-HER2 antibodies, and also for poor prognosis in breast cancer particularly in NP cases (Purdie et al., 2010).
The tyrosine kinase HER2 controls many aspects of tumorigenesis such as metabolism, cell cycle, and proliferation (Zhang et al., 2005). We found that several glycolytic enzymes and known poor prognostic markers in breast cancer were more commonly reacted with sera from HER2+ cases. A characteristic of cancer cells is enhanced glycolysis, a mechanism that may help cancer cells to proliferate in hypoxic environments, such as conditions within tumors, and to escape from apoptosis. A correlation between the glycolytic phenotype and tumor invasiveness has also been suggested (Diaz-Ruiz et al., 2010). Consistently, overexpressions of glycolytic enzymes are frequent reports in most types of cancers as well as breast that are perturbed in HER2+ cells (Zhang et al., 2005). For the first time, we observed the antigenicicty of GAPDH (3/9 of HER2+ and 1/9 of HER2− cases), LDH-A (5/9 of HER2+ and 2/9 of HER2− cases), and also enolase-α (spot 3 in 7/9 of cases in each subgroup, and spot 11 in 4/9 and 3/9 of HER2+ and HER2− cases, respectively) in breast cancer, the three glycolytic enzymes whose antigenicities have been observed in other types of cancer, and they were recommended as diagnostics, prognostics, and/or therapeutics for cancer patients (Chang et al., 2006, Nakanishi et al., 2006). Although the numbers of sera from HER2+ cases with reactivity against enolase-α did not differ from HER2− cases in the present study, however, a trend of increase in intensity of enolase-α spots in HER2+ cases was observed in blots (Supplementary File 2), and spot 11 was disappeared from blots that were probed with pooled sera from HER2− cases (Fig. 2). It seems that the determination of a cutoff point for the right interpretation of enolase-α immunoreactivity, and possibly other spots, is necessary. In this regard, a scoring method based on intensity of immunohistochemical staining and determination of a cutoff point revealed an association between enolase-α expression status and poor prognosis in lung cancer (Chang et al., 2006). Another glycolytic antigen in our study was TPI, an enzyme whose immunoreactivity has been reported in breast cancer as well as other cancers (Shukla et al., 2009; Tamesa et al., 2009). The frequency of TPI immunoreactivity between sera from HER2+ and HER2− patients showed no difference (five patients in each subgroup). Similarly Tamesa et al., in Japanese, found no differences in TPI immunoreactivity between sera from HER2+ and HER2− breast cancer patients (Tamesa et al., 2009).
In HER2+ NP breast cancer, PHGDH antibody was detected in 55% (5/9), but 22% (2/9) in HER2− cases (Supplementary File 2 and Table 2). A recent publication indicates that overexpression of this enzyme is markedly associated with shorter time to relapse and survival in breast cancer. PHGDH is involved in biosynthesis of serine, an amino acid that is necessary for cell proliferation (Pollari et al., 2010). Autoantibody against this protein has been reported in lung cancer (Nakanishi et al., 2006).
The more occurrences of autoantibodies against known aberrantly expressed, poor prognostic markers and/or putative therapeutic targets in sera from HER2+ patients compared to HER2− patients deserves further attention on other predominant antigenic proteins in HER2+ patients, including pyrophosphates, STI1, and proteasome 26S non-ATPase subunit 13. The expression or antigenicity of none of them has been investigated in breast cancer so far. The aberrant expression of the two former antigens has been shown in other types of cancers, and STI1 has also been recognized as a tumor antigen in ovarian cancer (Kim et al., 2010; Kuramitsu and Nakamura, 2006). Although overexpressions of several subunits of proteasomes have been reported in cancer (Kuramitsu and Nakamura, 2006; Tan et al., 2005), the expression of proteasome 26S non-ATPase subunit 13 is not clear in cancer. This subunit is a component of the 19S regulatory particle (RP). 19S RP is the key regulatory component of the 26S proteasome by which the majority of proteins are degraded (Tan et al., 2005). Proteasome 26S non-ATPase subunit 13 was antigenic in 44.4% (4/9) of HER2+ and 11.1% (1/9) of HER2− cases (Table 2). Interestingly, the only HER2− case with reactivity against this protein was our only HER2 and ER-negative case. HER2 and ER-negative breast cancers also represent a subgroup of the disease with poor prognosis (Purdie et al., 2011).
The frequencies of immunoreactivities of sera from HER2− cases against 14-3-3ɛ, PP2AR, and ribonuclease inhibitor-angiogenin were more than those from HER2+ cases (>2). From a theoretical point of view, PP2AR and ribonuclease inhibitor-angiogenin might suppress tumorigenesis. 14-3-3ɛ has also been shown to have some interactions with PP2A (Dickson et al., 2009; Dougherty and Morrison et al., 2004; Wong et al., 2010). 14-3-3 proteins through binding to proteins containing phosphothreonine or phosphoserine motifs participate in phosphorylation-dependent protein–protein interactions that control multiple pathways during tumorigenesis such as signal transduction, apoptosis, cell cycle progression, DNA replication, and cytoskeletal dynamics. 14-3-3 proteins have multiple isoforms with distinct tissue localizations and isoform-specific functions (Dougherty and Morrison et al., 2004). Among them, it has been proven that 14-3-3γ gene polymorphism is associated with breast cancer risk only in HER2− cases (Olson et al., 2011). Recent studies now indicate that the serine/threonine PP2A is an important regulator of 14-3-3 binding interactions (Dougherty and Morrison, 2004). PP2A is a major serine/threonine protein phosphatase implicated in the negative control of cell growth and division by dephosphorylating specific substrates. It has been shown that this phosphatase is inactivated in HER2+ cells (Wong et al., 2010). A subunit of PP2A hetrotrimer holoenzyme is PP2AR whose mutations have been recently linked to breast cancer development and progression (Wong et al., 2010). Whether these genes alterations are the cause of PP2AR aberrant expression and antigenicity in our study needs more investigations. Ribonuclease inhibitor-angiogenin expression and antigenicity in cancer tissues are not well understood in cancer. Angiogenin induces neovascularization, and plays a role in the establishment, growth, and metastasis of tumors. Ribonuclease inhibitor-angiogenin binds to angiogenin to modulate its activity and neovascularization (Dickson et al., 2009).
The ratios of immunoreactivity of HER2+/HER2− or HER2−/HER2+ patients' sera against other identified spots were ≤2. They were hnRNP K, hsps, prohibitin, nucleophosmin, glyoxalase, IDH3, eIF3, pp32, chaperonin containing TCP1, TCTP, and ETFA (Table 2).
The autoantibody against hnRNP K, heat-shock protein (hsp) 27, hsp90, hsp70, prohibitin, and nucleophosmin were previously reported in 30–50% of breast cancer patients (Brankin et al., 1998; Conroy et al., 1998; Hamrita et al., 2008), which are in keeping with our findings (Table 2). Theses antigens are involved in key process of tumorigenesis and their aberrant expressions are also noted in breast cancer (Brankin et al., 1998; Conroy et al., 1998; Hamrita et al., 2008). In addition to diagnostic values, immunoreactivities against hsp27, hsp90 and nucleophosmin were shown to also have prognostic values (Brankin et al., 1998; Conroy et al., 1998).
The antigenicities of glyoxalse, IDH3, eIF3, pp32, chaperonin containing TCP1 and TCTP have not been reported in breast cancer. The aberrant expression of the four former antigens have been reported in breast cancer and the two latter antigens in other types of cancer (Kadkol et al., 2001; Kuramitsu and Nakamura, 2006, Silvera et al., 2010; Zang et al., 2004; Zhang et al, 2005; Zhu et al., 2008). The glyoxalase pathway leads to the detoxification of methylglyoxal, a cytotoxic byproduct of glycolysis. Methylglyoxal causes protein and DNA modification and subsequently cell death. The glyoxalase overexpression has been observed in several types of cancer as well as breast particularly in HER2+ cases (Zhang et al., 2005). We found its antigenicity in 44.4% (4/9) of HER2− and 33.3% (3/9) of HER2+ cases. Similarly, although TPI overexpression is more pronounced in HER2+ cells (Tamesa et al., 2009), but its antigenicity was irrespective of HER2 status. IDH3 is a mitochondrial enzyme that catalyzes the third step of the citric acid cycle: the conversion of isocitrate to alpha-ketoglutarate. The mutation in other isoforms of IDH has been linked to cancer that its real mechanism is not clear (Yan et al., 2009). eIF3 has translation initiation factor activity during protein synthesis through its interaction with the small ribosomal subunit and the eIF4F. eIF3 is increased in various cancers such as breast. This overexpression is thought to promote tumorigenesis through the stimulation of global protein synthesis (Silvera et al., 2010). The antigenicity of eIF3 has been reported in liver cancer (Chen et al., 2008). pp32 plays important roles in many cellular processes including regulation of chromatin remodeling, transcription, RNA transport, transformation, and apoptosis. pp32 is described as a new tumor suppressor. Its mutation and underexpression have been shown in prostate cancer. However, in breast cancer, pp32 is an abundant protein (Kadkol et al., 2001). When MCF7 blots were probed with pooled sera, pp32 spot was reacted with sera from HER2− cases, but not with sera from HER2+ cases (data not shown). It has been previously shown that MCF7 cell line is a good source for antigens in breast cancer (Hamsher et al., 2007). MCF7 is a low-expressing HER2 cell line with high expression of ER. We checked some of our sera against 2D blots from SKBR3 cell line that highly expresses HER2 with no ER expression. We compared the positions of 28 immunoreactive spots in MCF7 blots with paired positions in SKBR3 blots. ETFA was not present in both SKR3 gels and blots. The differential expression (more than twofold) of LDHA, pyrophosphatase, and nucleophosmin were observed between SKBR3 and MCF7 blots, and their immunoreactivities were just partly different between SKBR3 and MCF7 blots. Further investigation with other breast cancer cell lines with different levels of cancer biomarkers such as MDA-MB-231 (ER− and HER2−) is required to recognize more antigens in different types of breast cancer.
Most of current widely tumor markers such as CA125 are detectable in normal individuals and their raise from a cut off point is considered as a cancer biomarker (Medeiros et al., 2009). Our identified antigens had no or slight reactivity with healthy sera, indicating that by making a suitable cutoff point in ELISA test, these antigens may be potent diagnostic serum biomarkers. However, predominant antigens in HER2+ cases may have lower frequencies in overall breast cancer patients, because HER2 overexpression occurs in less than 20% of breast cancer patients (Purdie et al., 2010). Therefore, predominant antigens in HER2+ cases may have lower frequencies in overall breast cancer patients. This fact may make them trivial as diagnostics in commercial tests. Moreover, all patients studied here were NP and relatively in advanced stages. There is a possibility that patients in earlier stages do not produce autoantibodies against these proteins. Nonetheless, these antigens still shed more light on breast cancer immunology and biology. We found differential antigenicities of some known prognostic markers and some novel molecules between HER2+ and HER2− NP breast cancer patients, the two subgroups of patients that were previously shown to have substantial differences in prognosis (Purdie et al., 2010). These molecules may prove to have prognostic and targeted-therapeutic values in future studies. For example, we found the antigenicity of proteasome 26S non-ATPase subunit 13 in breast cancer patients, particularly in HER2+ cases. Different classes of proteasome inhibitors targeting different subunits of proteasome complex are evaluating in clinical trials (Dupire and Coiffier, 2010). Whether proteasome 26S non-ATPase is suitable for a targeted intervention in breast cancer needs more investigation.
2D Western blotting may be helpful for giving a hint on aberrantly expressed proteins and possibly response to a particular therapy. This method may not need protein identification by MS. As mentioned, our MS protein identification revealed the same proteins as in previously published MCF7 proteome (Ou et al., 2008).
Conclusions
In conclusion, we identified several antigens in NP breast cancer patients by 2D Western blotting. Some of them were previously known to be antigenic in cancer, whereas others were novel. The frequency of known breast cancer antigens in our study was partly similar to those in reports where their investigations were conducted by the ELISA method in a larger number of patients. It remains to be identified the exact frequencies of our novel antigens by ELISA in a larger number of patients, and their potential uses as prognostics and therapeutics for NP breast cancer patients.
Footnotes
Acknowledgments
This work was funded by grants from the Shiraz University of Medical Sciences (87-4303 and 89-01-16-2548) and the Institute for Cancer Research (ICR-87-503).
Author Disclosure Statement
All authors declare no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.