
Abstract
Abstract
Cadmium is a toxic heavy metal causing iron deficiency in the shoot and light sensitivity of photosynthetic tissues that leads to decreased photosynthetic performance and biomass production. Light intensity had strong impact on both photosynthetic activity and metal accumulation of cadmium-treated plants. At elevated irradiation, cadmium accumulation increased due to the higher dry mass of plants, but its allocation hardly changed. A considerable amount of iron accumulated in the roots, and iron concentration was higher in leaves developed at moderate rather than low irradiation. At the same time, the higher the irradiation the lower the maximal photochemical quantum efficiency. The decreased photochemical efficiency, however, started to recover after a week of Cd treatment at moderate light without substantial change in metal concentrations but following the accumulation of green fluorescent compounds. Both cadmium treatment and higher light caused the accumulation of flavonoids in leaf mesophyll vacuoles/chloroplasts, but accumulation of flavonols, fluorescing at 510 nm, was characteristic to cadmium stress. Therefore, flavonoids, which may act by scavenging reactive radicals, chelating Cd, and shielding against excess irradiation, play an important part in Cd stress tolerance of Populus, and may have special impact on its phytoremediation capacity.
Introduction
Numerous metabolic inhibitory effects of Cd are related to the perturbations of metal homeostasis of plants (Clemens, 2006; Solti et al., 2011). Strong Fe deficiency develops under Cd stress in the shoot (Larbi et al., 2002), whereas roots were shown to accumulate Fe (Shao et al., 2007). Iron translocation into the shoot is the most retarded process also in poplar (Fodor et al., 2005). In iron shortage induced by Cd treatment, the development of thylakoid membranes, and synthesis of chlorophylls (Chl) and Chl-protein complexes are highly retarded, resulting in modified composition of thylakoid membranes (Fagioni et al., 2009). The diminished efficiency of the photosynthetic electron transport was also attributed to either direct or indirect effects of Cd on photosystem II (PSII) (Faller et al., 2005; Qureshi et al., 2010; Sigfridsson et al., 2004). As a first effect of Cd, the nonphotochemical quenching by the out-of-function PSII reaction centers increases (Solti et al., 2009) similar to the plants suffering from photoinhibition (Hendrickson et al., 2005). It points out that the relative excess of light may also contribute to the inhibition of photosynthetic performance by Cd stress. However, we still have no information on how light intensity can affect the development of Cd toxicity symptoms and whether it is connected with the Cd-induced changes in the Fe homeostasis.
Photosynthetic pigments have dual roles in plants: their main task is to collect light for the photosynthetic reaction centers but dissipation of excess light to protect reaction centers against overexcitation, which would lead to the damage of photosynthetic electron transport, is of equal importance (Murchie and Niyogi, 2011). However, there are other pigments, which also protect cell components against high visible and UV irradiation damaging many biological molecules (Takahashi and Badger, 2010). In most of the higher plants, phenolic pigments, such as flavonoids and anthocyanins, accumulate under high light irradiation, which also have a predicted role in heavy metal stress as antioxidant or chelating compounds (Procházková et al., 2011). Cd is known to enhance the phenylalanine ammonia lyase (PAL) biosynthetic pathway (Herbette et al., 2006), which leads to the accumulation of phenolic compounds (Dai et al., 2006; Finger-Teixeira et al., 2010; Kováčik and Bačkor, 2007), as well as flavonoids and anthocyanin pigments in the leaves (Krupa et al., 1996). Flavonoids also contribute to Cd detoxification in the vacuoles based on their heavy metal binding ability (Bravo and Anacona, 2001). Nevertheless, the effect of these compounds is still not resolved in the long-term resistance against Cd stress.
Therefore, the processes of photosynthesis/light protection—Fe transport—Cd accumulation may be in close connection during natural Cd stress, which can strongly influence the capacity of poplar trees for phytoremediation. Here we investigate some aspects of their interactions.
Materials and Methods
Plant material
Experiments were performed on micropropagated poplar (Populus jacquemontiana var. glauca (Haines) Kimura cv. Kopeczkii) plants, grown in climate chamber with 14/10-h light/dark periods, 24/22°C and 70/75% relative humidity (VPD: 0.89/0.66 kPa) in quarter-strength Hoagland nutrient solution: 1.25 mM Ca(NO3)2, 1.25 mM KNO3, 0.5 mM MgSO4, 0.25 mM KH2PO4, 0.08 μM CuSO4, 4.6 μM MnCl2, 0.19 μM ZnSO4, 0.12 μM Na2MoO4, 11.56 μM H3BO3, and 10 μM Fe(III)-citrate as the Fe source. Light intensities were adjusted to 40 (“L40”), or 80 (“L80”), or 250 (“L250”) μmol photons m−2 s−1. In their four-leaf stage, plants were further grown under control conditions (“−Cd”) or treated with 10 μM Cd(NO3)2 (“+Cd”) for 3 weeks. The three to four normal leaves present at the beginning of the treatment (older leaves) were marked with negative numbers starting from the youngest one, while another four to five leaves developed under the treatment (younger leaves) got positive numbers starting from the oldest developing leaf. The part of the stem developed before or after the start of treatment is referred to as lower and upper stem, respectively.
Determination of ion content
Ion content was determined by total reflection X-ray fluorescence spectrometry. Digestion of air-dry plant organs was performed in a pressure-controlled microwave digestion system (MDS-2100 CEM Corporation, Matthews, NC). The analysis of the digested samples was carried out by an EXTRA IIA total-reflection X-ray fluorescence spectrometer manufactured by Atomika Instruments GmbH (Oberschleissheim, Germany). Molibdenum (Mo) and tungsten (W) microfocus X-ray tubes of a power of 50 kV and current intensity of 38 mA, equipped with high-energy cutoff filters (quartz-glass mirrors) and attenuation filters [for the molybdenum (Mo) tube: 200 mm Mo, 240 mm aluminum (Al); for the W(B) tube: 1,000 mm Al, 100 mm nickel (Ni)], were used for excitation. A silicon(lithium) [Si(Li)] detector of 80 mm2 in area recorded the fluorescence photons emitted by the elements of the samples. The integration time was 500 s for each sample. The energies of the analytical lines, used for the determination, were the following: Fe Kα 6.398 keV and Cd Kα 23.106 keV.
Determination of phenolic content
Cumulative absorbance between 280 and 367 nm after the digestion of leaf tissues in 62.5% methanol, 0.6 M HCl at 90°C for 60 min was used as the measure of total phenolic content (Goldschmid, 1954).
Leaf fluorescence measurements
Fluorescence induction measurements of leaf samples were performed using a PAM 101-102-103 Chlorophyll Fluorometer (Walz, Effeltrich, Germany) according to Solti et al. (2008). Maximal quantum efficiency of the photosystem II (PSII) Fv/Fm=(Fm−F0)/Fm was used to monitor the physiological status of the leaves.
Fluorescence imaging was performed by a compact flash-lamp imaging system (Szigeti, 2008) according to Solti et al. (2008). Shortly after, excitation of the adaxial side of the light-adapted leaves was carried out by xenon-lamp flashes (λexc=360–370 nm) at room temperature and the detection of fluorescence was performed by a CCD video camera at different wavelengths using appropriate interference filters. Corrections and arithmetical operations were performed by Camille 1.05 software (Photonetics, Kehl, Germany).
Room-temperature fluorescence emission spectra of intact leaves were measured between 400 and 700 nm (λexc=365- nm slit widths: 1 nm) by a Fluoromax-2 fluorimeter (Jobin Yvonne, France).
Fluorescence microscopy was performed on native thick cross-sections of poplar leaves without any fixation or embedding by an Olympus BH-2 fluorescence microscope using UG2 excitation filter (λexc=360–370 nm). Fluorescence was monitored using a λ=510–525-nm emission filter.
Results
Plants were grown at three different light intensities, which deeply influenced their growth, metal accumulation, and photosynthetic performance under Cd stress. The maximal quantum efficiency of PSII reaction centres (Fv/Fm) showed the most characteristic decline in parallel to the rising light intensity: although it did not decrease significantly under L40 treatment compared to −Cd plants, L80 or L250 caused moderate and marked decrease, respectively, by the end of the second week of treatment (Fig. 1). It is important to note that even L250 treatment did not cause any decrease in Fv/Fm of −Cd plants, indicating that this light intensity is not photoinhibitory.
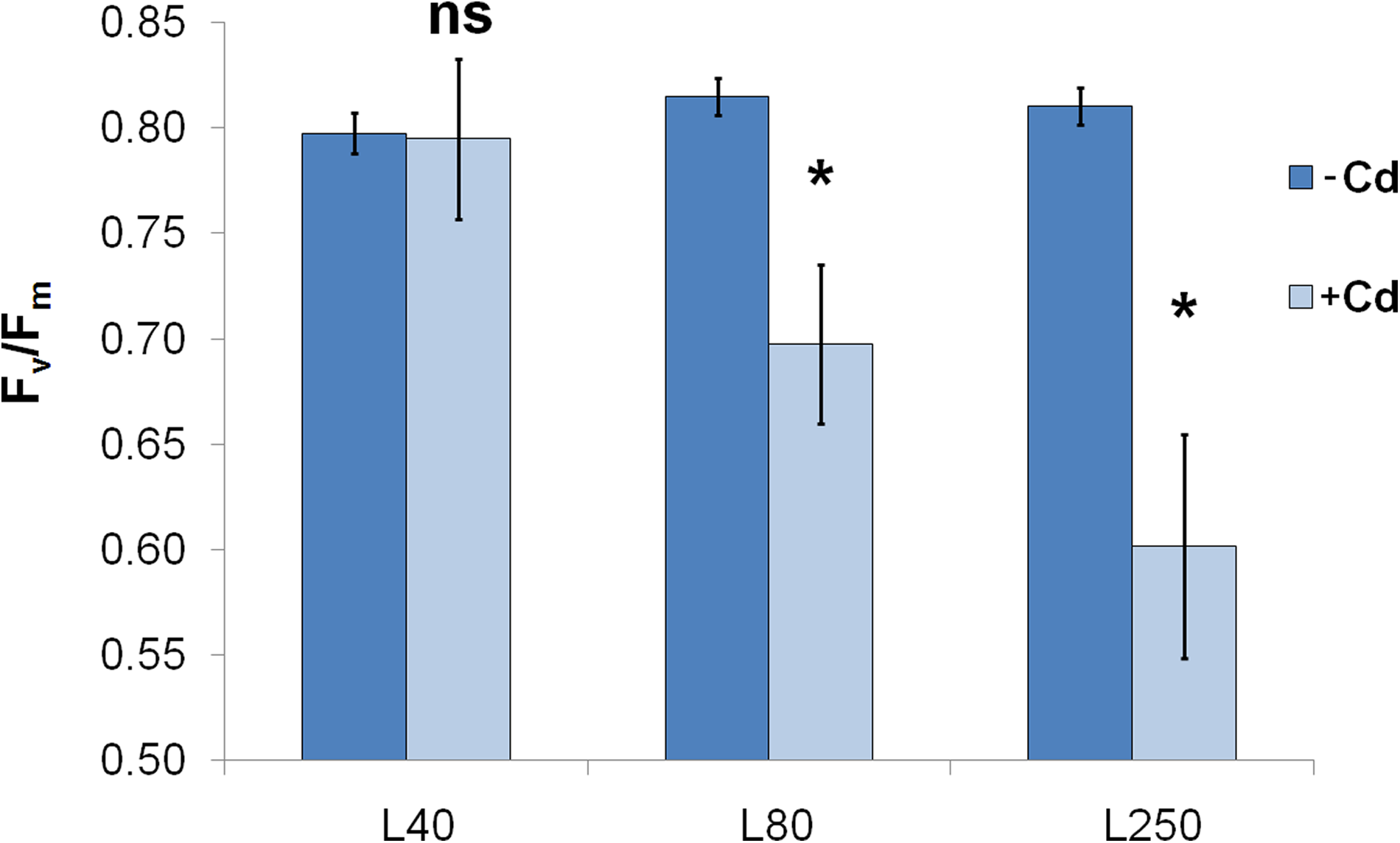
Maximal quantum efficiency of PSII reaction centres (Fv/Fm) of the +2 leaves in differently treated plants after 2-week treatment. Differences were: ns: nonsignificant, *Significant (Student's t-test, P<0.05), L40, L80, L250: light conditions during growth and treatment (daytime photon flux density in μmol m−2 s−1).
The rise in the light intensity affected the Cd accumulation in the organs. After 2 weeks of Cd treatment, plants grown at L80 accumulated 7.64±0.53 μmol Cd, and the total dry biomass of treated plants was 1.049±0.045 g. Although L250 treatment did not cause any significant change in the Cd accumulation of +Cd plants compared to L80+Cd treatment, L40 significantly decreased the Cd content together with an average of 45.5±4.0% retardation in the dry weight of plants compared to L80+Cd treatment (Fig. 2). However, different light treatments hardly changed the allocation of Cd among different organs; Cd accumulated mostly in root tissues (Fig. 2). Cd concentration was only slightly elevated in the part of plants developed under the treatment (Table 1). Longer treatment hardly changed the Cd content of plants (not shown).
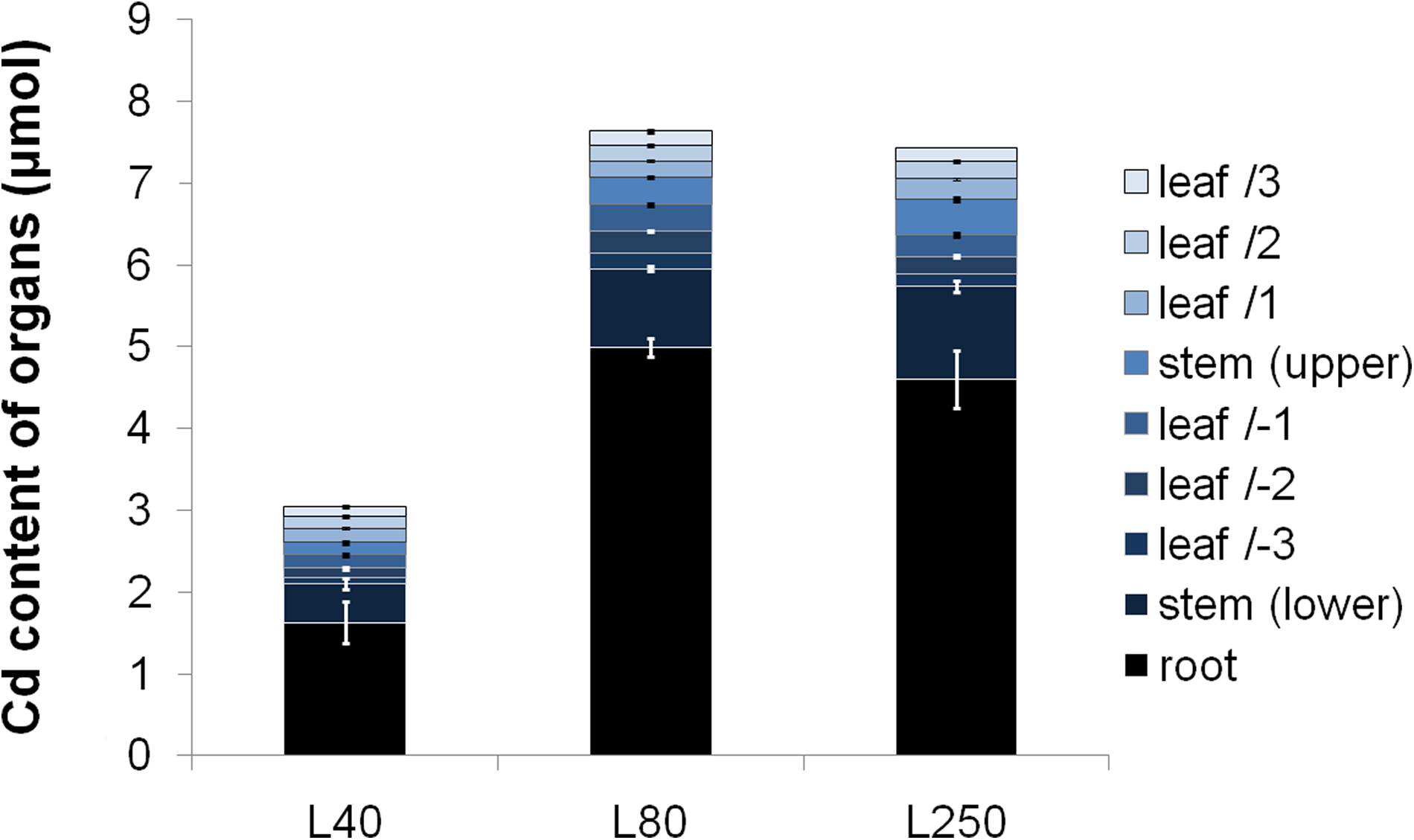
Cd allocation in Populus treated with 10 μM Cd at different light conditions for 2 weeks. Cd accumulation decreased significantly in the different organs under L40 treatment (ANOVA, p<0.05). Upper and lower refer to development before and during the treatment period, respectively.
L250 data are in the percentage of L40 treatment. Upper and lower refer to development before and during the treatment period, respectively.
Fe content of poplar plants was also strongly affected by light intensity. Under L40−Cd treatment, the Fe accumulation decreased compared to the L80−Cd treatment, whereas the Fe concentration did not change in agreement with the decreased biomass growth (Fig. 3 and Table 2). L250 treatment did not change the Fe accumulation but slightly decreased the Fe concentration compared to L80–Cd plants in accordance with enhanced dry weight growth. Under Cd stress, an elevated iron accumulation was observed in the roots, whereas the shoot organs contained significantly less Fe (Fig. 3). Both Fe accumulation and concentration decreased bottom-up in the shoot of +Cd plants, independently of irradiation conditions. L80+Cd-treated plants accumulated 32.57±2.28 μmol Fe in the root (204% of the L80–Cd plants), whereas only 0.66±0.01 μmol Fe was translocated to the shoot (52% of the L80–Cd plants). Compared to L80+Cd, L40+Cd treatment caused a significant decrease in Fe accumulation both in the root and the shoot: Fe content of the shoot was lowered but Fe accumulation in the root was comparable to that of L40–Cd plants. Under L250+Cd treatment, neither the root nor the shoot Fe accumulation changed compared to L80+Cd conditions. The Fe concentration was similar in L40+Cd and L80+Cd, but higher in L250+Cd leaves. The smallest difference in Fe concentrations were measured between +Cd/−Cd pairs under L250 conditions due to the decreased Fe concentration of L250–Cd plants. Three-week treatment did not change the iron content of organs any further (not shown).
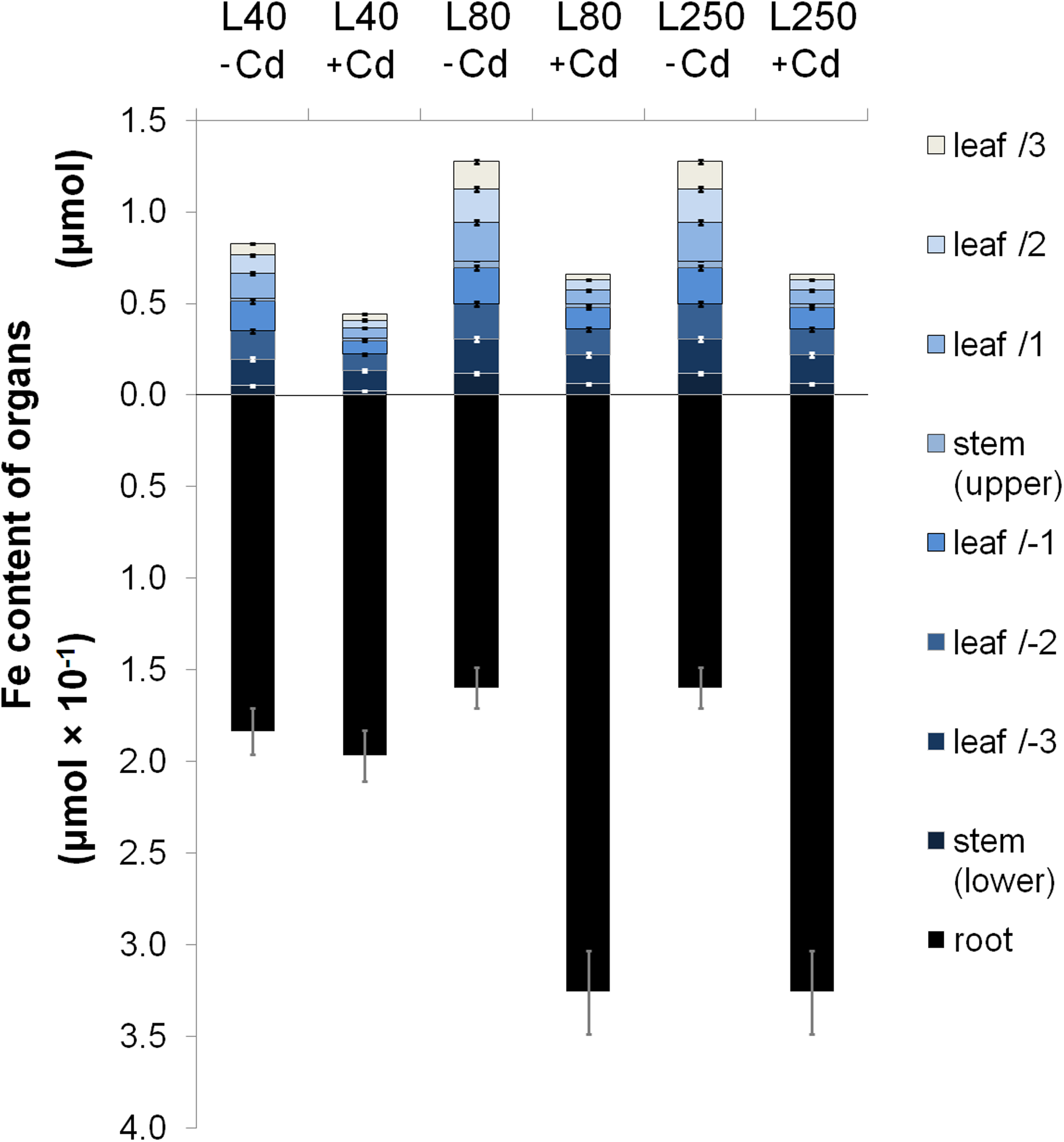
Fe allocation in Populus grown at different light intensities in the presence or the absence of Cd. Cd treatments as well as L40−Cd decreased the shoot iron content significantly (ANOVA, p<0.05). Upper and lower refer to development before and during the treatment period, respectively.
+Cd data are in the percentage of their −Cd pair. Upper and lower refer to development before and during the treatment period, respectively.
Studying the time course of Fv/Fm, a strong and significant decrease was found up to the 10th day of the L80+Cd treatment, where its value stabilized for a while, then as the treatment went on, Fv/Fm started to increase (Fig. 4), and it completely recovered to the level of −Cd plants after 3 weeks of Cd treatment. More or less similar changes were observed in the green fluorescence of leaves measured by multicolor fluorescence imaging (F520). In the leaf expansion period F520 decreased in both −Cd and +Cd-treated leaves. After the area expansion ceased, its level remained constant in −Cd leaves, but started to increase in +Cd leaves; a permanent increase was observed from the seventh to the eighth day of treatment. Similar to L80+Cd treatment, L250–Cd treatment, itself, also caused an increase in F520, whereas F440 did not show significant changes (not shown). In the total phenoloid content of leaves, a similar, significant increase was found in both L80+Cd and L250–Cd treatments (Fig. 5) where the ratio of increase in the measurable total phenoloid content (179.4% and 344.7% of L80–Cd values in L80+Cd and L250–Cd plants, respectively) was similar to the increase of green fluorescence of the leaves: cumulative F480–600 increased to 159.3% and 167.2% of the L80–Cd values in L80+Cd and L250–Cd plants, respectively, but F520/F440 changed to 289.0% and 177.7% of the L80–Cd values in L80+Cd and L250–Cd plants, respectively).
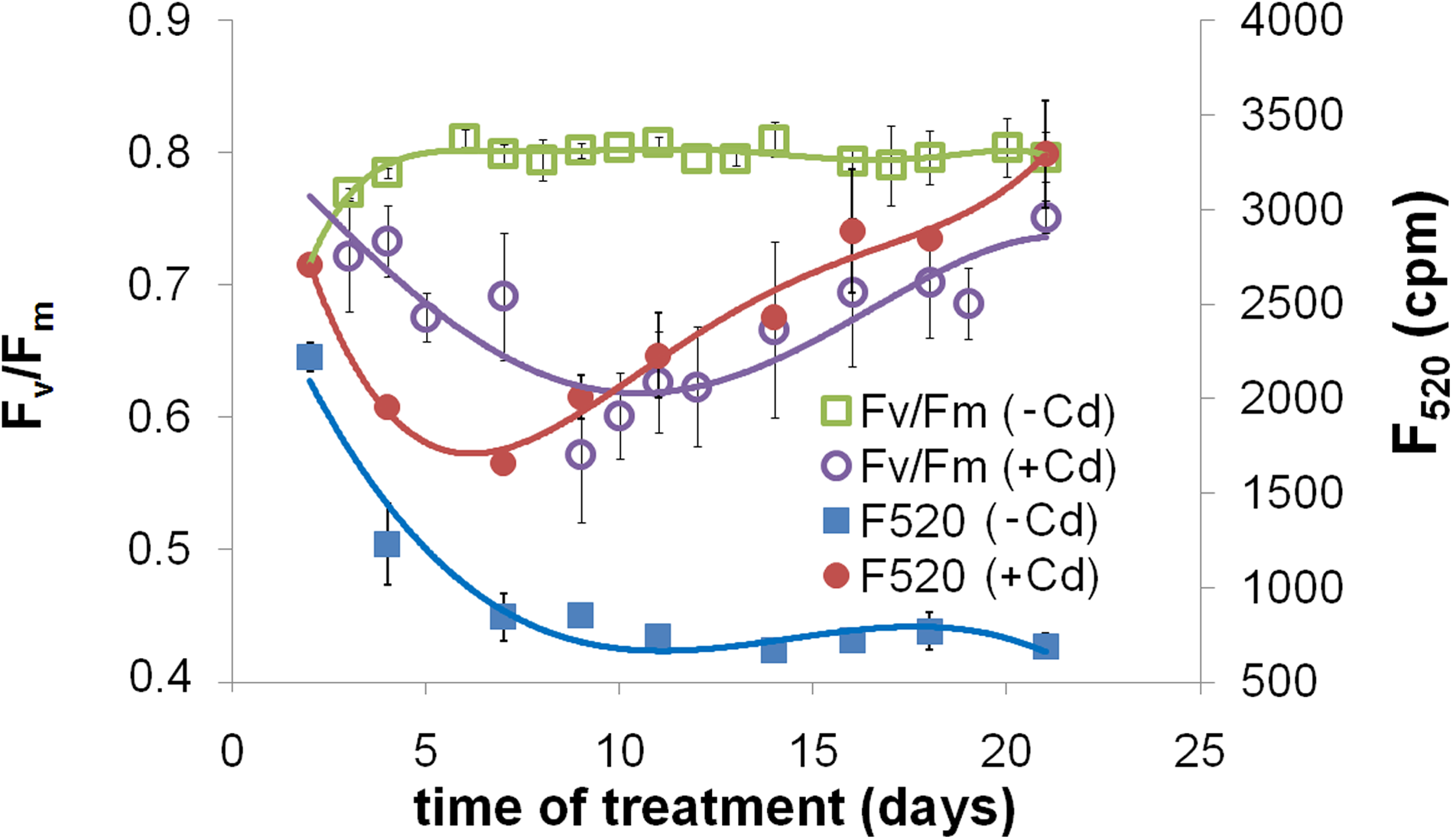
Time course of changes in the values of the Fv/Fm and F520 parameters in the +2 leaves of L80–Cd and L80+Cd plants.
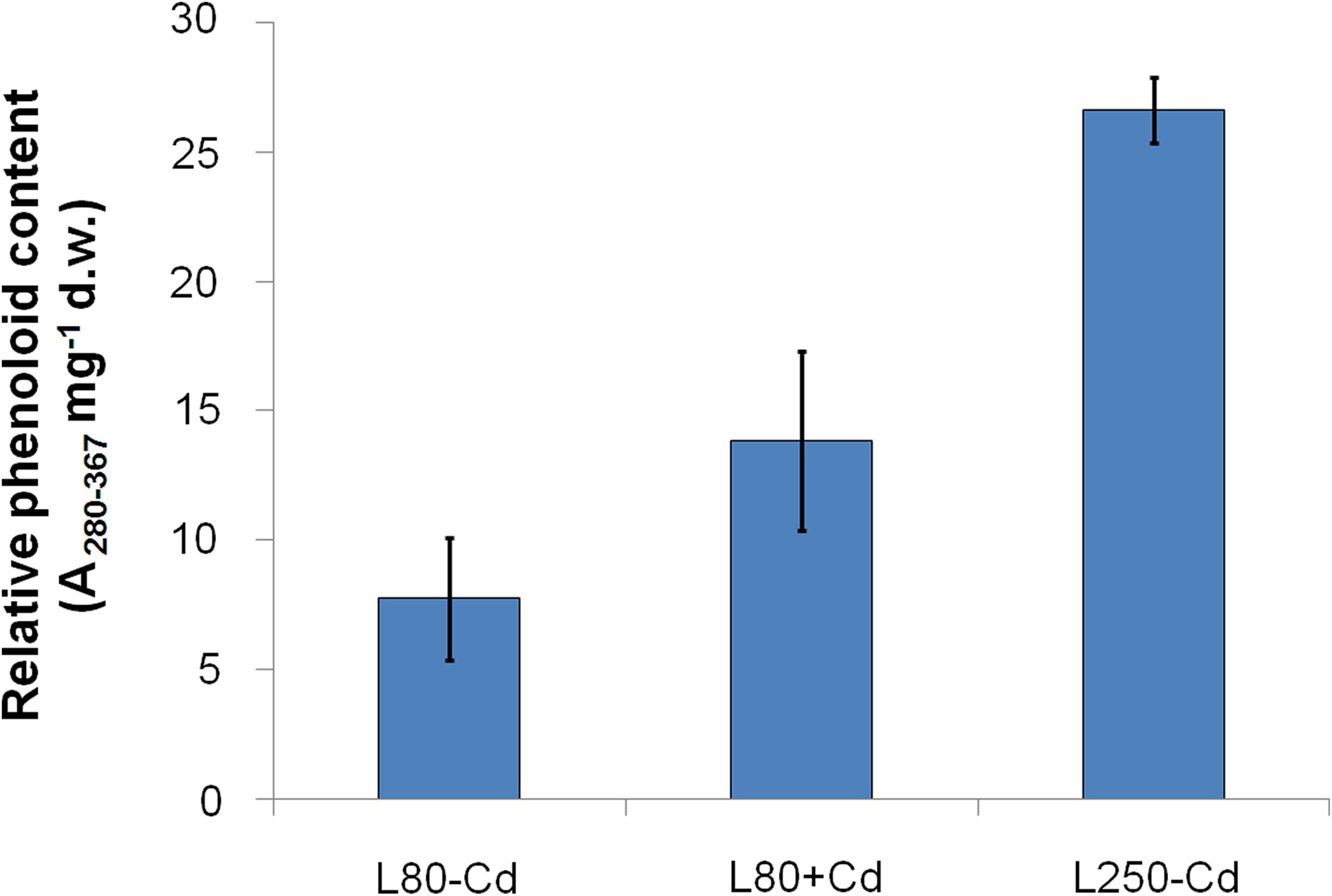
Total phenoloid content (cumulative absorbance between 280 and 367 nm mg−1 dry weight) of +2 leaves after 3-week treatment. Differences between L80−Cd and L80+Cd/L250−Cd leaves were significant (one-way ANOVA with Tukey-Kramer posttest, p<0.05).
In the fluorescence spectra of intact leaves, four major components showing fluorescence in the 500–600 nm regions at λex=365 nm were identified: an F510, an F530, an F550, and an F560 (Fig. 6A). The amount of these components, however, changed somewhat differently under L80+Cd and L250–Cd treatments (Fig. 6B). Although a strong increase in F510 was observed in L80+Cd leaves, and there was moderate increase in F530, F550, and F560, all the peaks except of F510 showed a strong increase, F560 being the most markedly elevated under L250–Cd treatment.
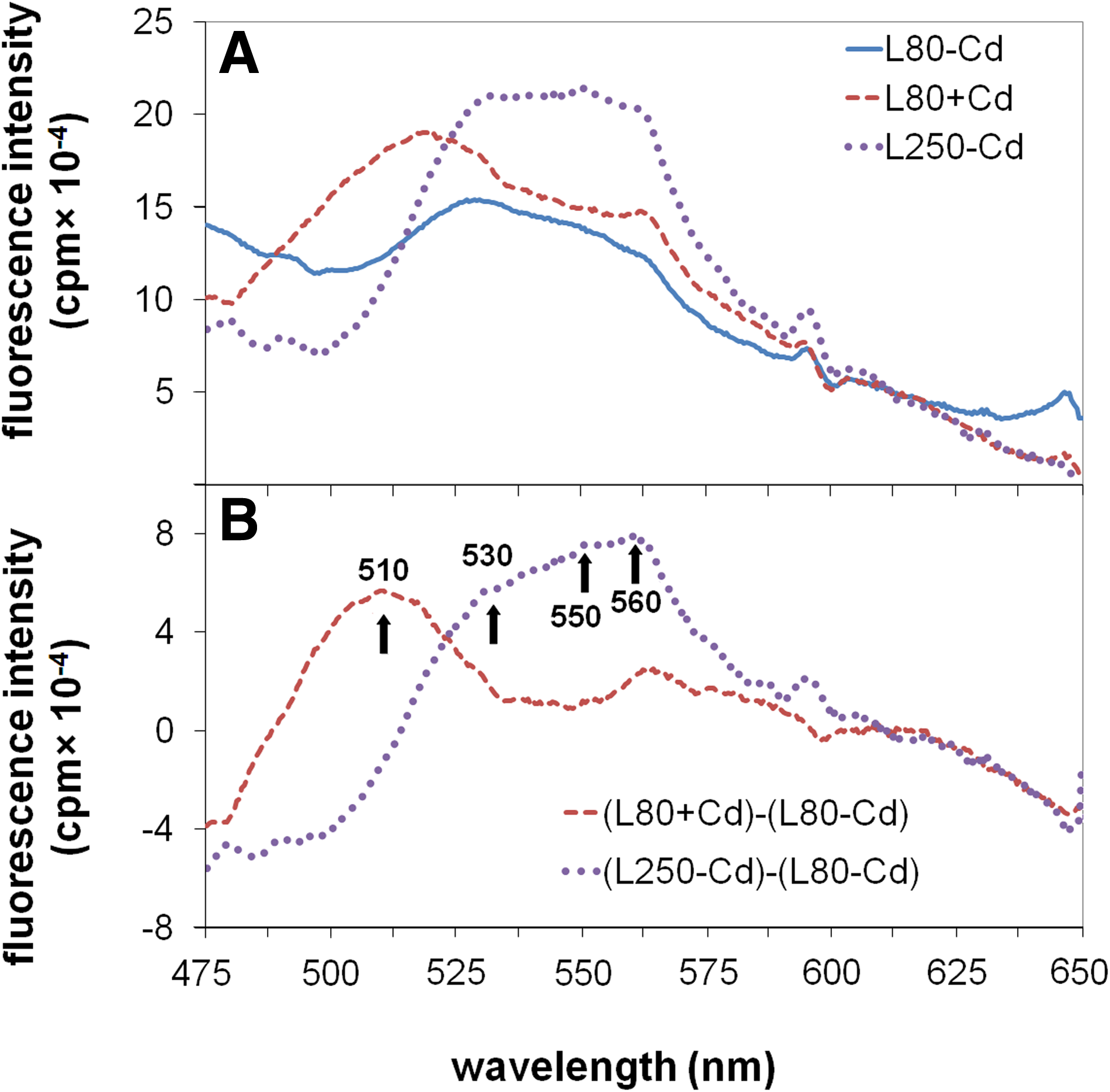
Native fluorescence spectra (λex=365 nm) of +2 leaves after 3-week treatment (
In the L80–Cd leaves, green fluorescence observed by fluorescence microscopy originated mainly from the cell walls, particularly from the secondary walls of the vascular elements and external walls of the epidermis (Fig. 7A). In the epidermis, green fluorescence originating from the vacuoles was not detected. Although the green fluorescence of cell walls also increased slightly, the compounds accumulated under L80+Cd stress and L250–Cd treatment appeared mainly in the vacuoles and chloroplasts of mesophyll cells (Fig. 7B,C) Both L80+Cd and L250–Cd treatments caused the thickening of the mesophyll layer.
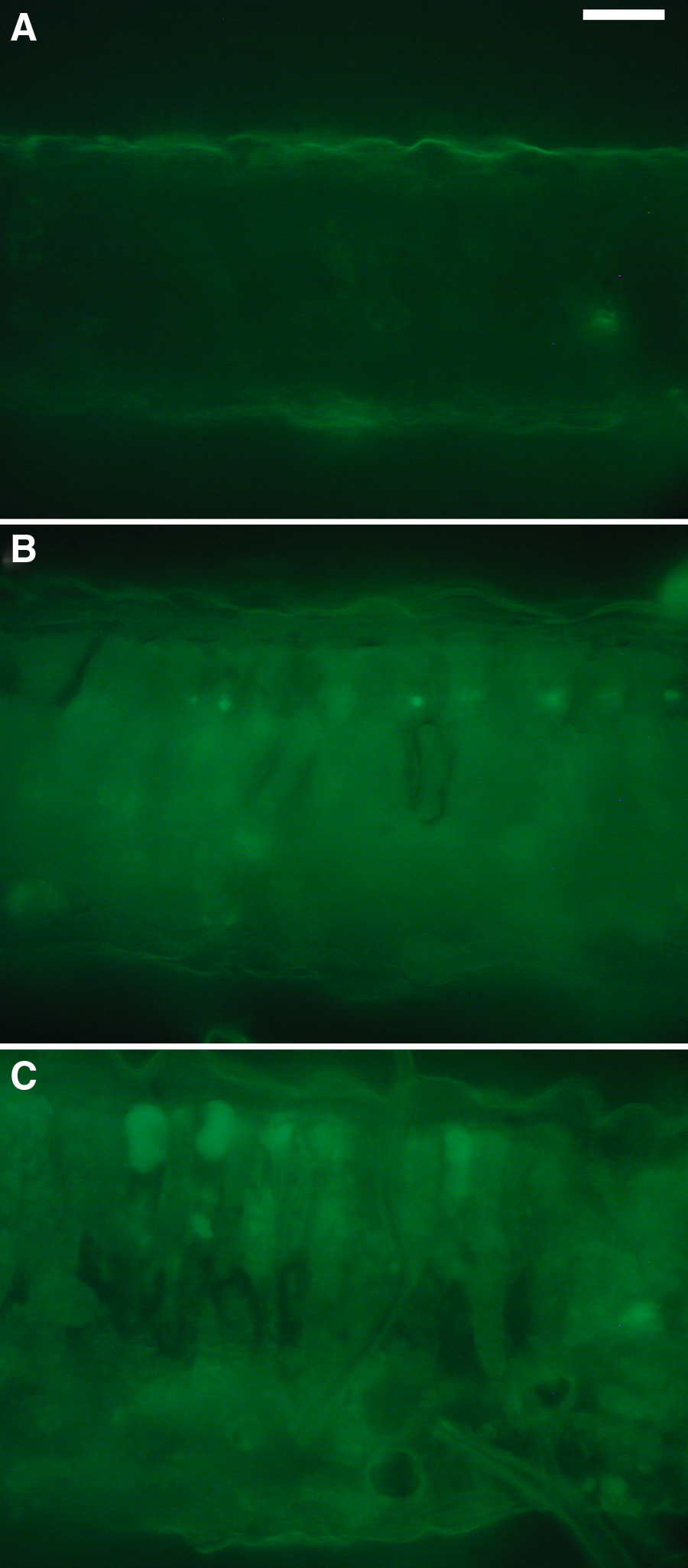
Green fluorescence (λfilter=510–525 nm) in native cross-section in +2 leaves of L80−Cd (
Discussion
Effects of Cd stress on Cd and Fe uptake and translocation, as well as on photosynthetic activity of poplar, were influenced by a light intensity-dependent manner. In Cd-treated plants, Cd uptake and translocation into the organs were enhanced by the increase in light intensity, whereas changes in the growth had only moderate effect on the Cd concentration in leaf tissues. It points to the main role of light-influenced biomass growth in the Cd accumulation capacity of poplar. Iron homeostasis under Cd stress was also affected by light intensity. Iron uptake into the roots (Chang et al., 2003) as well as Fe translocation, which also needs citrate as chelator for uptake into the xylem (Rellán-Álvarez et al., 2010), are energy-dependent processes. Thus, they were strongly retarded in L40–Cd plants due to their lower assimilation activity. At higher light intensities, Cd treatment massively enhanced the iron uptake into the roots, and although Cd markedly decreased the translocation of iron into the shoot, iron concentration moderately increased in leaves developed at higher irradiation compared to L40+Cd plants. The maximal quantum efficiency of PSII reaction centres (Fv/Fm), one of the most important parameters for the characterization of photosynthetic activity, can be affected directly by Cd binding, by Cd-induced iron deficiency, and high light stress (Lichtenthaler et al., 2007; Morales et al., 1991; Pietrini et al., 2010). In the case of iron deficient plants, however, low Fv/Fm was found to be an underestimation caused by the reduction of the PSII acceptor side in the dark (Belkhodja et al., 1998). Therefore, both higher Cd binding and low Fe concentration, depending on the compartmentalization of these ions in the leaves/cells, may contribute to the stronger inhibition of maximal PSII efficiency at higher light intensities.
The decrease in the PSII quantum efficiency in younger leaves under the coexistence of Cd treatment and elevated irradiation refers to a Cd-induced light sensitivity. Cd treatment is known to cause similar changes in PSII function as photoinhibition (Hendrickson et al., 2005; Solti et al., 2009). After the inhibitory phase, however, a recovery of PSII activity was observed, in parallel to or even following the accumulation of green fluorescent compounds in the leaves. The accumulation of these compounds could be observed both in L250–Cd plants and in L80+Cd plants. Lichtenthaler and Babani (2000) showed that blue and green fluorescence increase under many stresses. Green fluorescent compounds were shown to accumulate in the walls of leaf epidermis cells, the fluorescence of which was not modified by the reabsorption of chlorophylls, and also in the mesophyll cells (Mantha et al., 2000). Under iron deficiency, the green fluorescence originating from leaf mesophyll increased as a part of the large increase in blue fluorescence (Morales et al., 1994). The green emission of L250–Cd and L80+Cd leaves was slightly different from that of the green emission of the L80–Cd leaves originated only from the external cell walls of the epidermis, probably due to the broad fluorescence spectra of blue-emitting cell wall phenolics like ferulic acid (Morales et al., 1996).
The green fluorescence of leaves is known to originate mainly from the emission of cinnamoids, quercetin, and other flavonoids (Meyer et al., 2003), which all belong to the family of phenoloids. Among flavonoids, flavonols are the main emitters in the green region (Morales et al., 1996; Roshal et al., 1999), whereas flavins perform a higher wavelength, yellow region fluorescence (Cerovic et al., 1994; Morales et al., 1994). The synthesis of flavonoids is known to be induced by UV stress (Harborne and Williams, 2000). Flavonoid pigments (e.g., anthocyanins) were also shown to accumulate under heavy metal stress (Krupa et al., 1996). Similar to some other stresses, the polyphenol and phenoloid content of the mesophyll cell wall increases under Cd stress (Vollenweider et al., 2006). PAL is a key enzyme in phenoloid biosynthesis, the activity of which was shown to be increased under Cd stress (Kováčik and Bačkor, 2007). Among the first responses to Cd stress, the expression of flavonoid-3′-hydroxilase, a key enzyme in the quercetin biosynthesis pathway, is strongly elevated (Herbette et al., 2006). The constitutive expression of chalcone synthase, a key enzyme in flavonoid biosynthesis, leads to an extremely enhanced UV tolerance in Arabidopsis (Bieza and Lois, 2001), and flavonoid biosynthesis defective mutant lines showed decreased tolerance against Cd stress (Keilig and Ludwig-Müller, 2009). Both L250–Cd and L80+Cd treatments increased the green emission. The composition of the accumulated flavonoids seems to be different (Fig. 6). The F510 compound, which accumulated under L80+Cd treatment, may belong to flavonols (Cerovic et al., 2002; Morales et al., 1996), whereas F530–F560 compounds, which accumulated under L250–Cd but partially also under L80+Cd treatment, are most probably flavins (Cerovic et al., 1994; Morales et al., 1994).
Green fluorescent flavonoids are supposed to have a role in photoprotection by attenuating the irradiation (Götz et al., 2010). Although the primary role of F530–F560 compounds in the mesophyll of L80+Cd and L250–Cd plants is probably the protection against excess irradiation, the accumulation of F510 compounds, predominant under Cd treatment, may protect against the direct/indirect effects of Cd. Flavonoids are also known to behave as antioxidants, which can scavenge free radicals as well as reactive oxygen species, particularly in chloroplast (Agati et al., 2007). Cd itself is not redox active but—due to its interaction with the Fe metabolism—free ferrous ions may be produced that take part in Fenton reactions (Winterbourne, 1995) causing oxidative stress. Reactive oxygen species were shown to accumulate in cell wall and in chloroplasts under Cd stress (Zhang et al., 2009). In L80+Cd plants, the accumulation of green fluorescent compounds was also shown in the chloroplasts (Fig. 7). In addition, flavonols are known to chelate divalent metal cations, and metal chelation leads to the modification of their spectroscopic behavior (Roshal et al., 1999). As a strong flavonol accumulation was observed under L80+Cd treatment, role of flavonols in Cd chelation can be also postulated. Thus, F510 flavonol(s) may act as scavengers of reactive oxygen species and/or Cd chelators, whereas flavins fluorescing in the green–yellow region may help the protection against the relative excess of irradiation. Therefore, Cd-inducible flavonoids have key roles in the restoration of photosynthetic activity of leaves, and in the long-term tolerance against Cd stress.
Conclusion
Cd accumulation and the induced Fe-deficiency/incomplete Fe incorporation caused light sensitivity in Populus leaves even at moderate irradiation, which decrease the maximal efficiency of PSII. Protection of photosynthetic electron transport against photoinhibition by compounds absorbing in UV-A, and emitting in the green region can enhance the biomass growth. Therefore, flavonoids fluorescing in the green and yellow region, which are able to protect the sensitive photosynthetic structures by scavenging reactive radicals, chelating Cd, and shielding against excess irradiation, can play a relevant part in Cd stress tolerance of Populus, and may have special impact on its phytoremediation capacity.
Footnotes
Acknowledgments
We thank Zsuzsa Ostorics for technical assistance. This work was supported by the grant NN-84307 (ERA Chemistry–OTKA).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.