
Abstract
Abstract
The global burden of dengue continues to worsen, specifically in tropical and subtropical countries, and has evolved as a major public health problem. We investigated the changes in serum proteome in dengue fever (DF) patients from a dengue-endemic area of India to obtain mechanistic insights about the disease pathogenesis, the host immune response, and identification of potential serum protein biomarkers of this infectious disease. In this study, serum samples from DF patients, healthy subjects, and patients with falciparum malaria (an infectious disease control) were investigated by 2D-DIGE in combination with MALDI-TOF/TOF MS. The findings were validated with Western blotting. Functional clustering of the identified proteins was performed using PANTHER and DAVID tools. Compared to the healthy controls, we found significant changes in the expression levels of 48 protein spots corresponding to 18 unique proteins (7 downregulated and 11 upregulated) in DF patients (p<0.05). Among these differentially-expressed proteins, 11 candidates exhibited different trends in dengue fever compared to falciparum malaria. Importantly, our results suggest that dengue virus infection leads to alterations in expression levels of multiple serum proteins involved in diverse and vital physiological pathways, including acute phase response signaling, complement cascades, hemostasis, and blood coagulation. For the first time we report here that the serum levels of hemopexin, haptoglobin, serum amyloid P, and kininogen precursor, are altered in DF. This study informs the pathogenesis and host immune response to dengue virus infection, as well as the current search for new diagnostic and molecular drug targets.
Introduction
Since a range of serum proteins exhibit rapid alterations in expression patterns in response to disease condition and direct correlation with disease progression, investigation of pathogen-induced alterations of the human serum proteome has been found to be effective for understanding disease pathogenesis, host immune response, and for the identification of potential disease-related markers in different parasitic, bacterial, and viral infectious diseases, including falciparum and vivax malaria (Kassa et al., 2011; Ray et al., 2012a), leptospirosis (Srivastava et al., 2012), severe acute respiratory syndrome (Chen et al., 2004), and swine fever virus infection (Sun et al., 2011). Although over the last decade several omics-based studies have been conducted to explore the underlying molecular mechanisms associated with disease pathogenesis and the host immune response in dengue virus infections (Albuquerque et al., 2009; Thayan et al., 2009), the overall pathobiology of this disease remains largely unexplored.
Analysis of the blood proteome is an effective approach to decipher disease pathobiology and to identify diagnostic/prognostic marker proteins, since this biological fluid has constant intimacy with different body parts, and contains several classes of proteins released by diseased tissues (Issaq et al., 2007). Plasma is the liquid part of unclotted blood after removal of solid cellular components, while serum is plasma devoid of fibrinogen and other clotting factors. Previous studies have shown that differences in overall protein diversity of heparinized plasma and serum are negligible, aside from a lack of fibrinogen in the later (Zimmerman et al., 2012), and thus these fluids are often considered equivalent for many assays (Ladenson et al., 1974). Nevertheless, due to the removal of fibrinogen and other clotting factors, the overall protein content of serum is lower than that of plasma, which may be advantageous for the analysis of analytes present in trace amounts (Denery et al., 2011; Kronenberg et al., 1998; Ray et al., 2011).
Conventionally, dengue infection is diagnosed by employing dengue-specific IgM antibody, which detects disease after 5–10 days in primary dengue virus infection (Gubler and Meltzer, 1999). Emerging molecular diagnostic methods, viral isolation, or PCR, are effective for early diagnosis of dengue infection, but these techniques are expensive and time-consuming, and too costly for use as routine tests in most developing countries (Singh et al., 2010). Thus the establishment of early detection surrogates can help to meet the need for less expensive and better diagnostics and more effective therapy. The present study was conducted to investigate alterations in the serum proteome in dengue fever (DF) patients compared to healthy subjects in a dengue-endemic area of India (Mumbai, Maharashtra). Serum proteomic alterations in another clinically-relevant mosquito-borne infectious disease, falciparum malaria, have recently been investigated by our group (Ray et al., 2012b). To make the present study more comprehensive and informative, serum levels of the identified proteins, which exhibited altered expression levels in DF (compared to healthy subjects), were further examined in falciparum malaria patients (Ray et al., 2012b). The inclusion of falciparum malaria as a disease control in this comparative proteomic analysis helped to assess the specificity of the identified candidate proteins in DF, and to distinguish the generic febrile responses as well. While the foremost aim of this proteomic study was to investigate disease pathogenesis and host responses in dengue fever, another possible outcome could be the identification of early detection surrogates for this vector-borne viral infection.
Materials and Methods
Description of the study subjects
This study was undertaken with the approval of the Institutional Ethics Committee of Seth GS Medical College and KEM Hospital, Mumbai, and written informed consent was obtained from each participant prior to participation in the study and the sample collection process. Blood samples were collected from patients (n=6) suffering from DF diagnosed on the basis of clinical examination and confirmed by kit-based solid phase immuno-chromatographic assay using the dengue IgG/IgM cassette (DNG 431; Atlas Link Biotech, Anhui, China), and PCR-based assays following manufacturer's instructions. Blood samples were collected in the defervescent stage from DF patients with 2–5 days of fever. Patients were categorized as DF according to the WHO severity grading scale (Dengue hemorrhagic fever: diagnosis, treatment, prevention, and control; World Health Organization, 1997). Demographic, epidemiological, and clinical details, together with the past history of disease in all patients selected for this proteomic study, are documented in Supplementary Table S1.1 (see online supplementary material at http://www.liebertonline.com). Additionally, blood specimens were collected from age- and sex-matched falciparum malaria (FM) patients (n=8) as febrile disease controls, and healthy subjects (n=8), to perform comparative proteomic analysis. Patients with uncomplicated, non-severe FM diagnosed by microscopic examination and confirmed through rapid diagnostic testing (RDT) were enrolled for this proteomic study (Supplementary Table S1.2; see online supplementary material at http://www.liebertonline.com). All the subjects were aged 20–50 years. Sample collection, serum separation, and processing were performed as previously described (Ray et al., 2012a).
Serum sample processing and two-dimensional-difference gel electrophoresis (2D-DIGE)
Sample processing and 2D-DIGE were performed as described previously (Ray et al., 2012a). 2D-DIGE gels were scanned using an Ettan DIGE Imager scanner (GE Healthcare, Waukesha, WI) at 40 μm resolution using specific wavelengths and filters for Cy2, Cy3, and Cy5 dyes. Dye swapping was performed during labeling of the test and control samples to eliminate any dye effects. Comparative analysis for relative protein quantification across all the dengue, malaria, and control samples (HC), was performed using ImageMaster 2D Platinum 7.0 DIGE software (GE Healthcare).
In-gel trypsin digestion and mass spectrometry
The identity of differentially-expressed proteins (p<0.05) was established using an AB Sciex 4800 MALDI-TOF/TOF (AB Sciex, Foster City, CA) mass spectrometer. In-gel digestion and mass spectrometric analysis was performed as described previously (Ray et al., 2012a). The combined MS and MS/MS peak lists were searched using GPS™ Explorer software version 3.6 (AB Sciex). Protein identification was performed by MS/MS ion search using the MASCOT version 2.1 (http://www.martixscience.com) search engine against the Swiss-Prot database. Searches were carried out with the following parameters: all entries taxonomy, trypsin digestion with one missed cleavage, carbamidomethylation of cysteine residues as a fixed modification, oxidation of methionine residues as a variable modification, and mass tolerance 150 ppm for MS and 0.4 Da for MS/MS. Identified proteins having at least two unique matched peptides were selected for further analysis.
Western blot analysis
Western blot analysis was performed as described previously (Gollapalli et al., 2012). Briefly, protein concentration in each sample (DF and FM patients and HC) was estimated using the 2D-Quant kit (GE Healthcare), and BCA Protein Assay kit (Thermo Fisher Scientific, Pittsburgh, PA), following the manufacturers' instructions prior to the Western blotting experiment. Extracted serum proteins from HC, FM, and DF subjects were separated by 12% SDS-PAGE (50 μg per lane) and transferred onto PVDF membranes under semi-dry conditions using an ECL semi-dry transfer unit (GE Healthcare). Western blotting was performed using monoclonal/polyclonal antibodies against clusterin (sc-8354), hemopexin (sc-134443), and haptoglobin (sc-71207; all from Santa Cruz Biotechnology, Santa Cruz, CA), and appropriate secondary antibodies conjugated with HRP. ImageQuant software version 5.0 (GE Healthcare) was used for quantitation of signal intensity of the bands in the Western blots.
Protein networks and functional analysis
Functional pathway analysis was performed with the serum proteins showing altered expression levels in the DF patients using the Protein ANalysis THrough Evolutionary Relationships (PANTHER) system, version 7 (http://www.pantherdb.org; Thomas et al., 2006), and the Database for Annotation Visualization and Integrated Discovery (DAVID), version 6.7 (http://david.abcc.ncifcrf.gov/home.jsp; Huang et al., 2009a, 2009b), to identify associated gene ontology terms, biological processes, molecular functions, and physiological pathways, for better understanding of the biological roles of these identified proteins and their correlation with disease pathobiology.
Results
Study population profiles
The mean age of the DF patients selected for this proteomic analysis was 31 years (SD 9.71 years; range 20–46 years; median 30 years). The HC and FM patient populations were of comparable age distribution, with an average age of 26.50 years (SD 6.28 years; range 20–34 years; median 25.50 years), and 30.4 years (SD 9.60 years; range 20–45 years; median 28.50 years), respectively, to maintain uniform population profiles for differential protein expression analysis.
Differential expression of serum proteins in dengue viral infection
This comparative proteomic analysis was performed to identify differentially expressed serum proteins in DF patients. 2D-DIGE profiling identified 48 statistically significant (p<0.05) differentially-expressed protein spots in DF patients compared to the HC. Among the 48 differentially-expressed protein spots, 25 spots were downregulated (ranging from 1.20- to 3.26-fold), while the remaining 23 spots were upregulated (ranging from 1.27- to 3.93-fold; Supplementary Table S2; see online supplementary material at http://www.liebertonline.com). MALDI-TOF/TOF MS analysis revealed the identity of 26 protein spots (Supplementary Table S3 and Supplementary Fig. S1A; see online supplementary material at http://www.liebertonline.com); however, the rest of the spots remained unidentified due to their very low intensity and insufficient amounts of detectable peptides. The 26 protein spots identified by MS correspond to 18 (7 downregulated and 11 upregulated) unique differentially-expressed proteins (Table 1, Fig. 1, and Supplementary Fig. S1; see online supplementary material at http://www.liebertonline.com). Some of our identified differentially-expressed proteins such as apolipoprotein A-IV, complement C3, clusterin, and α1-antichymotrypsin were found to be consistent with an earlier plasma proteomic study performed on severe DF patients from an endemic region of Brazil (Albuquerque et al., 2009). Additionally, one of our identified targets, α1-antitrypsin, has previously been reported to be upregulated in serum samples of DF and DHF patients (Thayan et al., 2009). Correlations obtained between our results and those of previous proteomic studies enhanced the confidence of this analysis. Moreover, quite a few differentially-expressed proteins identified in this study, including serum amyloid P, haptoglobin, complement C4, complement factor H, hemopexin, and kininogen precursor have not been reported in earlier proteomics studies of DF. Some of these identified targets, although not reported in the context of dengue fever, can be correlated with DF disease pathogenesis on the basis of their known biological functions.
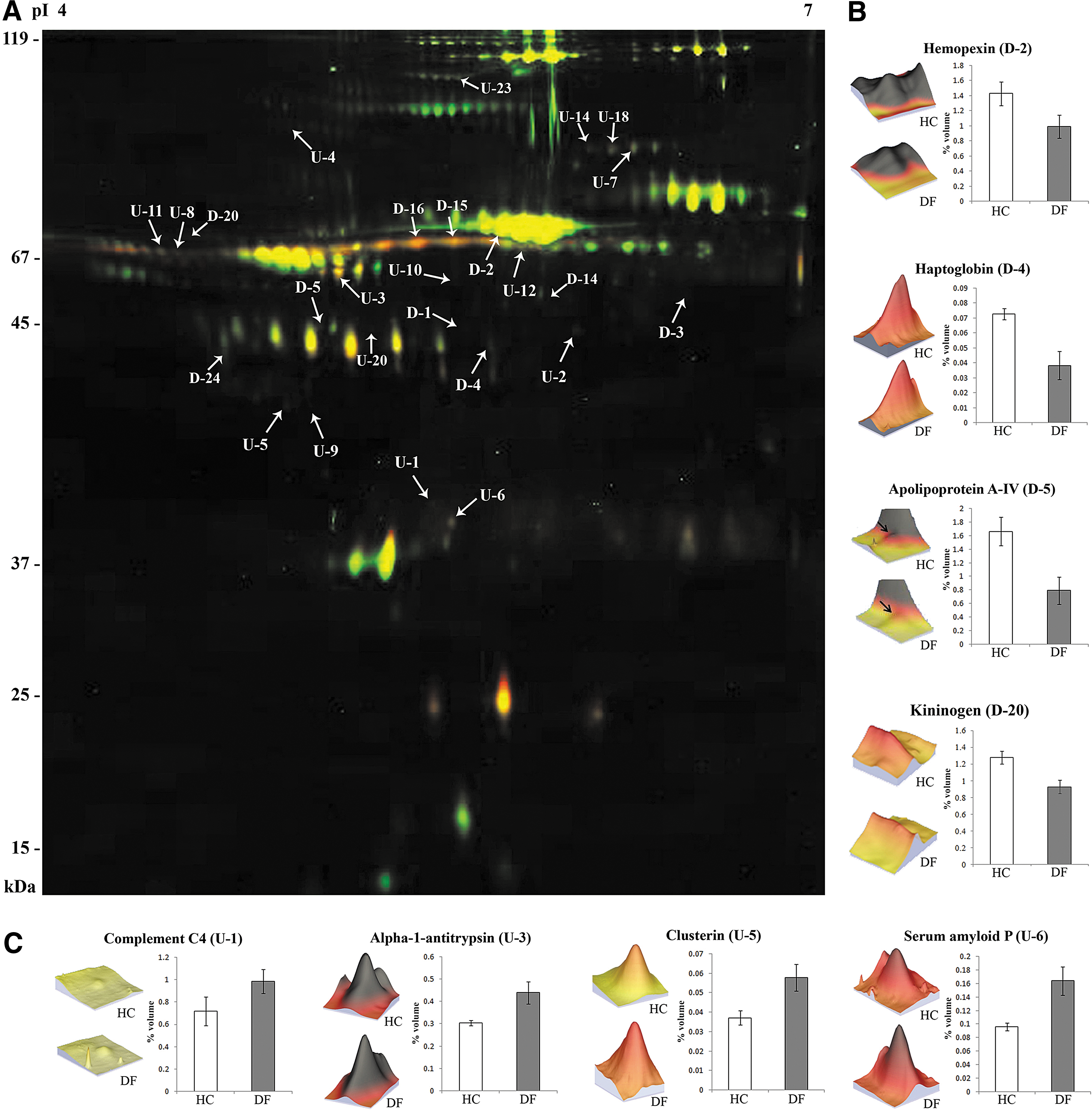
Differential expression of serum proteins in dengue fever (DF) patients identified by 2D-DIGE. (
For proteins with multiple spots in the 2D-DIGE gels, representative spot detail is provided. Exact values for each spot are provided in Supplementary Table S3.
A, protein binding; B, lipid binding; C, cell surface binding; D, antioxidant activity; E, chaperone binding; F, metal binding; G, drug binding; H, DNA binding; I, carbohydrate/sugar binding; J, peptidase/protease inhibitor activity; K, catalytic activity; L, lipid metabolism/transport; M, hemoglobin binding; N, enzyme regulator activity; O, enzyme inhibitor activity; P, sterol transporter activity; Q, complement activation; R, antigen binding; S, receptor binding.
Since proteomic analysis of DF in isolation cannot provide sufficient insight to understand the disease's pathogenesis, further comparative analysis was performed by including another mosquito-borne infectious disease, falciparum malaria, to establish the specificity of the identified dengue infection-related serum markers. There were four serum proteins that exhibited opposite trends of differential expression in DF patients and malaria patients (compared to healthy subjects), while seven differentially-expressed proteins identified in the DF patients exhibited no significant expression alteration in falciparum malaria patients (Supplementary Table S4 and Supplementary Fig. S1B; see online supplementary material at http://www.liebertonline.com).
Validation of differentially-expressed proteins
Validation of three interesting differentially-expressed proteins, clusterin, hemopexin, and haptoglobin was performed using Western blotting. CBB staining of the SDS-PAGE gels and Ponceau staining of the transferred blots containing the resolved proteins was performed to substantiate equal loading of the samples. Selection of the proteins for validation was done on the basis of fold changes, possible functional association of the proteins with dengue pathobiology, and availability of the required antibodies. Western blot analysis demonstrated overexpression of clusterin and downregulation of hemopexin in DF patients compared to healthy controls, while opposite trends of differential expression were observed in the malaria patients for these two serum proteins (Fig. 2). Reduced expression levels of haptoglobin were observed in both DF and malaria patients (Fig. 2). The relative intensity of clusterin was found to be 69.15±7.15, 32.68±7.34, and 102.37±6.74, in HC, FM, and DF patients, respectively. In the case of hemopexin, the trend of differential expression was the opposite, 49.95±5.76, 83.68±3.34, and 26.37±1.74, in HC, FM, and DF patients, respectively. The relative intensity of haptoglobin in healthy and febrile disease controls was found to be 49.70±3.34 and 18.35±1.34, while the DF patients showed a mean value of 28.82±3.57 (data are presented as mean±SE [relative intensity×104]; n=6; p<0.001 by Student's t-test; Fig. 2B). Similar trends of differential expression of the target proteins with comparable fold-change values were found in Western blots and 2D-DIGE gels (Supplementary Fig. S2; see online supplementary material at http://www.liebertonline.com), which validated the initial findings obtained from comparative gel-based proteomic analysis.

Western blot analysis of clusterin, hemopexin, and haptoglobin, from serum of dengue fever (DF) and falciparum malaria (febrile control) patients and healthy controls (HC). (
Alteration of physiological pathways in dengue fever
Functional pathway analysis using PANTHER revealed the association of differentially-expressed proteins in DF patients with the blood coagulation system (p=0.04; Supplementary Table S5.1; see online supplementary material at http://www.liebertonline.com), and exhibited involvement with 10 biological processes, including responses to stimuli (19.60%), metabolic processes (19.60%), immune system processes (19.60%), and cell communication (8.70%; Fig. 3A). PANTHER analysis classified the identified proteins in six protein classes (Fig. 3B), and five major GO functions [binding (39.10%), receptor activity (8.70%), enzyme regulatory activity (26.10%), catalytic activity (13%), and transporter activity (13%)] have been found to be associated with the identified differentially-expressed proteins (Fig. 3C). Interestingly, complement and coagulation cascades were also identified in DAVID analysis, under the KEGG category (p=5.01E−12; 38.89%; Supplementary Table S5.2), while different complement cascades (p=5.4E−06; 22.23%; Fig. 3D and Supplementary Fig. S3; see online supplementary material at http://www.liebertonline.com), and hemostasis (p=0.009; 22.23%), were identified in the Biocarta and Reactome categories, respectively.
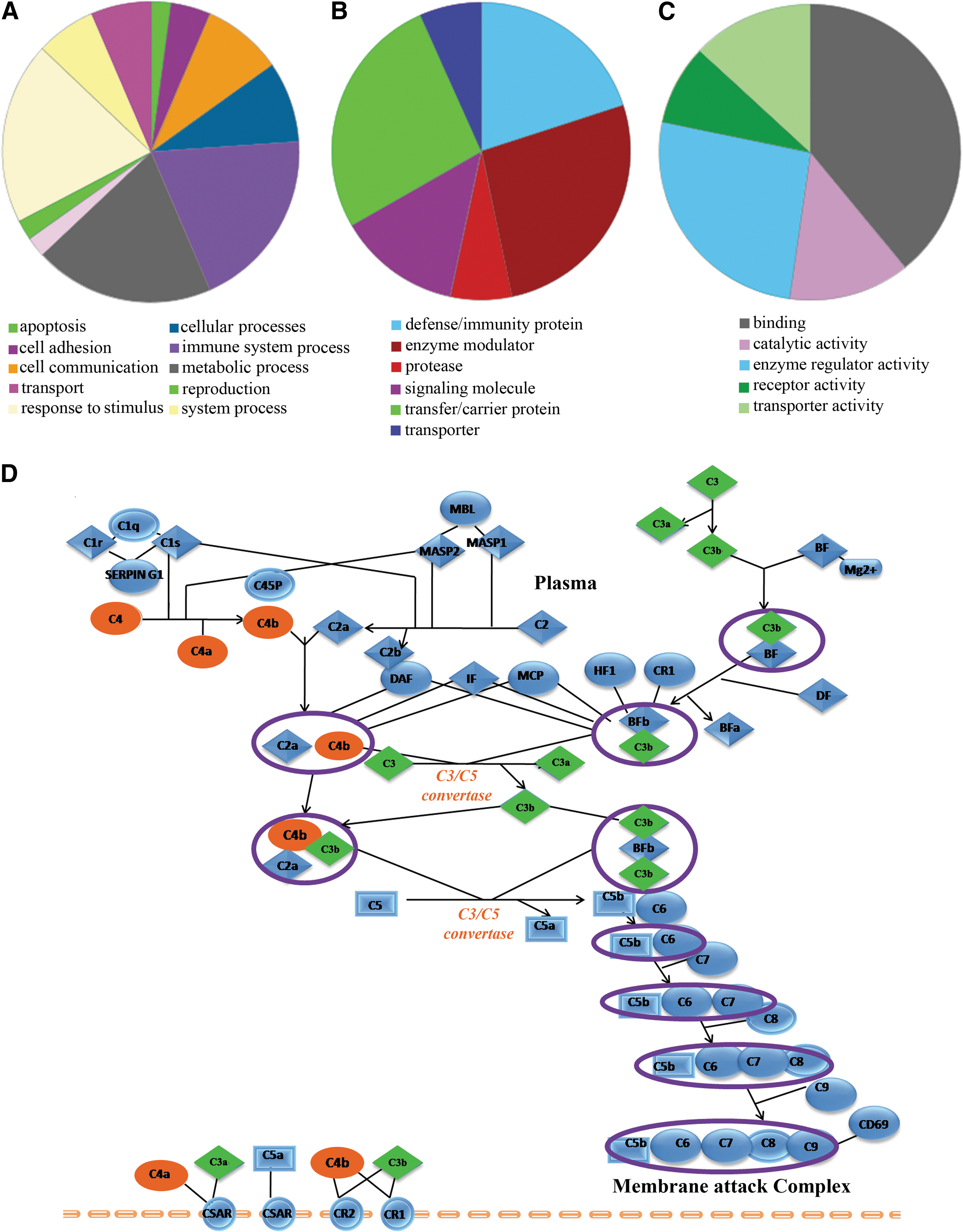
Functional clustering and physiological pathways associated with the differentially-expressed proteins identified in dengue fever (DF) patients. Pie charts show the biological processes (
Discussion
In this study we investigated human serum proteome alterations due to dengue fever, and identified altered expression levels of multiple proteins (Table 1) associated with coagulation and complement cascades, and inflammation-mediated acute phase signaling, probably as a consequence of this viral infection. There are earlier reports on the profiling of the plasma of dengue fever patients (Albuquerque et al., 2009), and serum of dengue hemorrhagic fever patients (Thayan et al., 2009) from other parts of the world. To the best of our knowledge, we report here the first comprehensive analysis describing the serum proteomic alterations seen in dengue fever patients from an endemic area of India. Compared to the previous proteomic studies, we have identified a larger number of differentially-expressed serum proteins in dengue fever, and performed functional pathway analyses employing these proteins to better understand the biological context of the proteins and their correlation with DF pathogenesis. Furthermore, in this proteomic analysis, in addition to healthy controls, serum samples from patients with another clinically-related vector-borne infectious disease, falciparum malaria, were analyzed to differentiate the generic febrile responses.
Dengue and malaria have significant overlapping endemic areas in the tropical and subtropical countries, and have similar transmission seasons. Although simultaneous transmission of both of these vector-borne infections is rare, there are quite a few published reports describing dual infections with dengue and malaria in endemic areas (Bhalla et al., 2006; Charrel et al., 2005; Kaushik et al., 2007; Ward, 2006). In addition, similarities in the clinical manifestations of dengue and malaria often make it difficult to identify the etiology of these two mosquito-borne infectious diseases. Accurate discrimination between these two infections and early diagnosis are essential for providing the proper treatments, which are quite different for the two diseases, in spite of the similarities in symptoms and signs. In light of the similarities between dengue viral and plasmodial infections in terms of symptoms, signs, and etiology, falciparum malaria patients were included as febrile disease controls in this comparative serum proteomic analysis to evaluate the specific host responses and proteomic alterations in dengue fever. Interestingly, a few serum proteins, such as serum amyloid P, kininogen, and complement factors H and C3 exhibited altered expression levels in DF patients, but not in falciparum malaria patients (Supplementary Table S4 and Supplementary Fig. S1B; see online supplementary material at http://www.liebertonline.com). Serum amyloid P (SAP), a member of the pentraxin family, exhibited a 1.7-fold increased expression in DF patients (Table 1). Earlier in vitro studies have shown that SAP can inhibit influenza A virus infection through binding of terminal N-acetyl-galactosamine, mannose, and glucuronic acid residues of the virus particle in a calcium-dependent process (Andersen et al., 1997). Reduced serum levels of kininogen were observed in DF patients, which may be due to the limited proteolysis of this precursor molecule for generating bradykinin to trigger inflammatory reactions via activating endothelial cells, and to participate in tissue hyper-responsiveness and local inflammation (Kaplan et al., 2002). Induction of the release of bradykinin leads to vascular leakage, and the subsequent hypotension and shock observed in severe dengue fever patients (Regoli and Barabé, 1980).
Activation of the complement cascades (classical and alternative), and the lectin-dependent pathways with the altered expression levels of C4, C3, C3a, C5a, and factor H, have been observed in DF patients (Table 1 and Supplementary Fig. S3 and Supplementary Table S5; see online supplementary material at http://www.liebertonline.com). An association of complement activation with dengue fever has been previously reported (Albuquerque et al., 2009; Avirutnan et al., 2011; Nascimento et al., 2009; Shaio et al., 1992); nevertheless, its exact relationship with disease pathogenesis and the mechanisms of activation remain unknown. Complement activation plays a vital role in protection against dengue viral infection through modulating adaptive immunity and activating virus-specific antibodies (Avirutnan et al., 2008). Activation of the complement system might be triggered by immune complexes generated in response to flavivirus nonstructural protein 1 (NS1) (Green and Rothman, 2006). Interestingly, a few complement factors and associated regulatory proteins such as clusterin, complement C4, and properdin factor B, have shown opposite trends of differential expression in DF and febrile controls (compared to HC; Supplementary Table S4 and Supplementary Fig. S1B; see online supplementary material at http://www.liebertonline.com). Elevated serum levels of clusterin were observed in this study (Fig. 2). This complement inhibitory factor interacts with the free soluble form of dengue virus (DV) NS1, and the NS1/Clu interaction may lead to the plasma leakage associated with DV infections (Kurosu et al., 2007). Avirutnan and associates have demonstrated that NS1-mediated complement activation involves vascular leakage, probably through the generation of anaphylatoxins and SC5b-9 during the severe manifestations of dengue viral infection (DHF and DSS; Avirutnan et al., 2006). According to a previous proteomic study, higher serum levels of dengue NS1 protein are observed in DHF patients compared with DF patients, indicating its possible correlation with disease severity (Thayan et al., 2009). Since the outcome of a productive infection depends on the balance between complement activation and evasion, detailed investigations of the functional aspects of complement factors and regulatory proteins could provide potential therapeutic targets.
Consistent with previous reports, our analysis also revealed altered expression of a few members of the blood coagulation cascade in DF patients (Huang et al., 2001). Although modulation of coagulation in the presence of disseminated intravascular coagulation (DIC) is most commonly associated with DHF, systemic host inflammatory and coagulation activation may occur in DF patients in the absence of severe hemorrhagic manifestations (Avila-Aguero et al., 2004). Dengue virus infection can also lead to endothelial cell injury (Butthep et al., 1993), which in turn may cause activation of the coagulation system and alteration of hemostasis in a direct or indirect way (Stemerman et al., 1984). However, the relevance of the activation of the coagulation system to the pathogenesis of dengue virus infection is not clear. The dengue fever patients analyzed in this proteomic study were suffering from primary infection, without any hemorrhagic phenomena (DHF) or shock (DSS). Further investigation of the modulation of the coagulation system and its associated plasma/serum proteins in dengue hemorrhagic fever could provide interesting insights into the disease's pathogenesis.
Serum proteins that exhibited similar trends of differential expression in DF and febrile controls were mostly acute phase proteins (APPs), and may be regular indicators of blood infection and inflammation. These APPs are found to be involved in inflammation-mediated acute phase signaling, and different host-adaptive and host-defense mechanisms. For instance, haptoglobin (Hp) was significantly downregulated in DF and FM patients compared to the healthy controls (Table 1). Hp possesses anti-inflammatory activity and innate phenotype-dependent antioxidant activity, it removes free hemoglobin (Hb) released during hemolysis, and it plays an important role in stimulating angiogenesis and has an inhibitory role in inflammation (Quaye, 2008). Downregulation of Hp reflects reduced levels of free Hp in patient sera, which is probably due to Hb-Hp complex formation and subsequent clearance by macrophages and monocytes. Elevated serum levels of different serine protease inhibitors, including α1-antitrypsin (1.44-fold), α1-antichymotrypsin (2.44- to 2.61-fold), and inter-alpha-trypsin inhibitor heavy chain H4 (1.5-fold), were observed in the dengue patients as well as in febrile controls. Over-expression of α1-antichymotrypsin and α1-antitrypsin have previously been reported in dengue fever and dengue hemorrhagic fever (Albuquerque et al., 2009; Thayan et al., 2009), and other viral infections such as SARS-coronavirus (Wan et al., 2006) and hepatitis B virus (Tan et al., 2011). Upregulation of these protease inhibitors might be a consequence of the host response to prevent the inflammation and vascular damage triggered by dengue virus infection.
Conclusions
We report here a comprehensive serum proteome analysis of DF patients from an urban population of India, and identified significantly altered expression of several serum proteins with various biological functions. This is the first study demonstrating human serum proteome alterations in DF in an Indian population. To the best of our knowledge, altered serum levels of serum amyloid P, haptoglobin, complement factors C4 and H, hemopexin, protein C6 orf165, and kininogen precursor, have not been reported in previous proteomics studies of DF. Some of our identified targets, including complement factors C4 and H, kininogen precursor, and clusterin, are interesting due to their functional relevance in DF pathogenesis, and thus warrant further research as potential inflammation-related biomarkers of DF. Future studies of these identified proteins in DHF and DSS may also show biologically relevant correlations with disease severity. Our observations will help to illuminate the pathogenesis and host immune response to DF, and may help in the search for new diagnostic and molecular drug targets to fight dengue virus infection.
Footnotes
Acknowledgments
We acknowledge the support of Dr. Urmila Thatte, Dr. Nithya Gogtay, Dr. Santosh Taur, and Tulip Jhaveri, of the Department of Clinical Pharmacology, Seth GS Medical College and KEM Hospital for clinical sample collection. We would also like to thank Dr. Rapole Srikanth and Snigdha Dhali from the National Centre for Cell Science (NCCS), Pune, for the help in performing the MALDI TOF/TOF MS experiments, and Dr. Priyanka Parte and Sumit Bhutada from the National Institute for Research in Reproductive Health (NIRRH), Mumbai, for their support in executing the 2D-DIGE experiments. A critical reading of the manuscript by Dr. Sayantan Ray of the Department of Medicine, Medical College, Kolkata, is also gratefully acknowledged. This research was supported by a start-up grant (09IRCC007) from the IIT Bombay to S.S.; S.R. was supported by an IIT Bombay fellowship. This study was undertaken with the approval of the Institutional Ethics Committee of Seth GS Medical College and KEM Hospital.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.