
Abstract
Abstract
Leptospirosis is one of the most important zoonoses. Leptospira interrogans serovar Lai is a pathogenic spirochete that is responsible for leptospirosis. Extracellular proteins play an important role in the pathogenicity of this bacterium. In this study, L. interrogans serovar Lai was grown in protein-free medium; the supernatant was collected and subsequently analyzed as the extracellular proteome. A total of 66 proteins with more than two unique peptides were detected by MS/MS, and 33 of these were predicted to be extracellular proteins by a combination of bioinformatics analyses, including Psortb, cello, SoSuiGramN and SignalP. Comparisons of the transcriptional levels of these 33 genes between in vivo and in vitro conditions revealed that 15 genes were upregulated and two genes were downregulated in vivo compared to in vitro. A BLAST search for the components of secretion system at the genomic and proteomic levels revealed the presence of the complete type I secretion system and type II secretion system in this strain. Moreover, this strain also exhibits complete Sec translocase and Tat translocase systems. The extracellular proteome analysis of L. interrogans will supplement the previously generated whole proteome data and provide more information for studying the functions of specific proteins in the infection process and for selecting candidate molecules for vaccines or diagnostic tools for leptospirosis.
Introduction
The genome of the pathogenic L. interrogans serovar Lai was first sequenced in 2003, and this sequence has been a useful tool for studying biology and pathogenesis of Leptospira, especially at the molecular level. Now, another six strains of Leptospira have been sequenced (Bulach et al., 2006; Nascimento et al., 2004; Picardeau et al., 2008; Zhong et al., 2011), and draft sequences for more than one hundred strains have been deposited in GenBank. These studies have greatly facilitated our knowledge of Leptospira physiology and pathology at the genomic, transcriptomic (Hesterlee, 2001), and proteomic levels (Adler et al., 2011). Extracellular proteins are important components of bacterial biology with many critical functions, such as nutrient acquisition, cell-to-cell communication, detoxification of the environment, and attacking potential competitors (Tjalsma et al., 2004). Moreover, the extracellular proteins could function as virulence factors in pathogenic bacteria (Lei et al., 2000; Tjalsma et al., 2004). Although many reports on leptospiral proteomics, from the outer membrane to the whole cell (Cao et al., 2010; Forster et al., 2010; Nally et al., 2011; Sakolvaree et al., 2007; Thongboonkerd et al., 2009; Vieira et al., 2009), have provided useful information, the precise identity and biological significance of the extracellular proteome remains largely unexplored. Moreover, the secretion apparatus used to deliver proteins to the extracellular environment by Leptospira is completely unknown. In this study, we analyzed the protein-related delivery systems based on whole-genome and whole-proteome data and further characterized the extracellular proteome in L. interrogans serovar Lai by liquid chromatography-tandem mass spectrometry (LC-MS/MS). This study contributes to a comprehensive overview of the secretion components and provides new candidates for further analysis of leptospiral virulence and protein function.
Materials and Methods
Preparation of protein-free C-70 medium
C-70 medium was modified from PF medium as described (Bey and Johnson, 1978) with additional supplementation of growth factor of solution A (5 mL per 1 L medium). The solution A was prepared (milligrams per 1 L of distilled water) as: vitamin B12 (4.00), benzene derivatives (4.00), vitamin B5 (8.00), L-glutathione (20.0), vitamin B6 (20.0), D-biotin (40.0), vitamin B3 (40.0), bitamin B1 (400), and L-asparagine (4000).
Culture conditions
The L. interrogans serovar lai type strain 56601 was obtained from the Institute for Infectious Disease Control and Prevention, Beijing, China and maintained by serial passage in hamsters to preserve its virulence. Leptospira that had been passaged in vitro for fewer than three generations were cultured in liquid Ellinghausen-McCullough-Johnson-Harris (EMJH) medium at 37°C or liquid-modified protein-free C-70 medium at 28°C under aerobic conditions for 48 h to early mid-phase at a density of approximately 108/mL. The cultures were centrifuged at 10,000 g for 10 min at 4°C to pellet the cells.
The growth of Leptospira in C-70 and EMJH medium was counted with a Petroff-Hausser chamber. Briefly, Leptospira were diluted to a density of 5×107cells/ml and cultured in liquid C-70 and EMJH medium at 28°C under aerobic conditions. Leptospira were enumerated using a dark-field microscope at every 6 h. Triplicate samples were counted.
Extraction of extracellular leptospiral proteins
Leptospira isolated from hamster and passaged in liquid C-70 medium in vitro for fewer than three generations were cultured in liquid C-70 medium at 28°C under aerobic conditions for 48 h to early mid-phase at a density of approximately 108/mL. 5×1010–1011 cells were harvested by centrifugation at 10,000 g for 10 min at 4°C and the supernatants were collected. The supernatant was filtered by a 0.22 μm filter unit (Millipore) to remove residual cells. Extracellular proteins were prepared by concentrating the supernatant with a 5000 Da MWCO filter (Sartorius, Viviflow 50), followed by precipitation with 4 volumes of acetone at 4°C overnight. The pellet was collected after centrifugation at 23,400 g for 30 min and dissolved in ddH2O. The extracellular proteins were monitored for purity by immunoblotting using antibodies against LA_2512, which is described elsewhere (Haake and Matsunaga, 2002).
LC-MS/MS analysis
The extracellular protein sample was digested to peptides by trypsin. An Ettan™ MDLC system (GE Healthcare) was applied to desalt and separate the tryptic peptide mixtures. In this system, the samples were desalted on RPtrap columns (Zorbax 300 SB C18, Agilent Technologies) and then separated on a RP column (150 μm i.d., 100 mm length, Column Technology Inc., Fremont, CA). Mobile phase A was 0.1% formic acid in HPLC-grade water and mobile phase B was 0.1% formic acid in acetonitrile. A 20 μg sample of tryptic peptide mixture was loaded onto the columns, and separation was performed at a flow rate of 2 μL/min using a linear gradient of 4%–50% phase B for 120 min. A FinniganTM LTQTM linear ion trap MS (Thermo Electron) equipped with an electrospray interface was connected to the LC setup to detect the eluted peptides. Data-dependent MS/MS spectra were obtained simultaneously. Each scan cycle consisted of one full MS scan in profile mode, followed by five MS/MS scans in centroid mode with the following Dynamic ExclusionTM settings: repeat count 2, repeat duration 30 sec, exclusion duration 90 sec. Each sample was analyzed in triplicate.
Data analysis
MS/MS spectra were automatically searched against the L. interrogans serovar Lai genome database (downloaded from GenBank http://www.ncbi.nlm.nih.gov/) using the BioworksBrowser rev. 3.1 (Thermo Electron, San Jose, CA). Protein identification results were extracted from SEQUEST out files with BuildSummary. Only tryptic peptides were considered, and up to two missed cleavages were allowed. The mass tolerances allowed for the precursor ions and fragment ions were 2.0 Da and 0.8 Da, respectively. The protein identification criteria used were Delta CN (≥0.1) and cross-correlation scores (Xcorr, one charge ≥1.9, two charges ≥2.2, three charges ≥3.75).
In silico analysis
The results were analyzed using several bioinformatic tools. SignalP 3.0 (http://www.cbs.dtu.dk/services/SignalP/) was used for signal peptide detection. SecretomeP (http://www.cbs.dtu.dk/services/SecretomeP/) was used to identify non-classical secretion pathway proteins. TMHMM (http://www.cbs.dtu.dk/services/TMHMM) was used for transmembrane structure detection. The SpLip program, kindly provided by D. Hakke (Research Service, Veterans Affairs, Greater Los Angeles Healthcare System, Los Angeles, CA), was used to detect leptospiral lipoproteins. Psortb (http://www.psort.org/psortb/index.html), cello (http://cello.life.nctu.edu.tw/) and SoSuiGramN (http://bp.nuap.nagoya-u.ac.jp/sosui/sosuigramn/sosuigramn_submit.html) were used to predict the localization of the identified proteins.
RNA extraction and real-time PCR detection
In vitro total RNA was extracted from L. interrogans using Trizol reagent (Roche). In vivo samples were collected from hamster livers. The hamsters were inoculated with approximately 108 L Leptospira and sacrificed at 72 h. Total RNA was extracted from the liver using Trizol reagent (Roche). Contaminating DNA was eliminated with RNase-free DNaseI (Roche), and the resulting RNA was purified using the RNeasy kit (Qiagen). The purified RNAs were converted to cDNA using the First Strand cDNA Synthesis Kit (Fermentas). Primers to amplify genes encoding extracellular proteins were designed with Beacon Designer software and are listed in Supplementary Table S1 (Supplementary Material is available online at www.liebertpub.com/omi). The gene transcript levels were normalized to the level of L. interrogans 16S, as described previously (Matsui et al., 2012).
Results and Discussion
Extracellular proteomic analysis of L. interrogans serovar Lai
Previous proteomic studies have mostly focused on leptospiral outer membrane proteins or vesicles. EMJH and Korthof's medium are commonly used to culture Leptospira, but both contain BSA or serum. Growth in a protein-free medium is preferable to obtain the pure secretion proteome. C-70 is the modified medium to culture the Leptospira without protein component in this study. The growth of Leptospira in C-70 medium reached the stationary phage at about 60 h, which was similar to that of EMJH (Fig. 1), although a slightly lower cell yield was achieved in C-70 than in EMJH medium. For preparing the extracellular protein, Leptospira were cultured in C-70 medium to early mid-phase, and the supernatant was collected as the secretion proteome and the purity was checked with SDS-PAGE and Western Blot (Fig. 2).
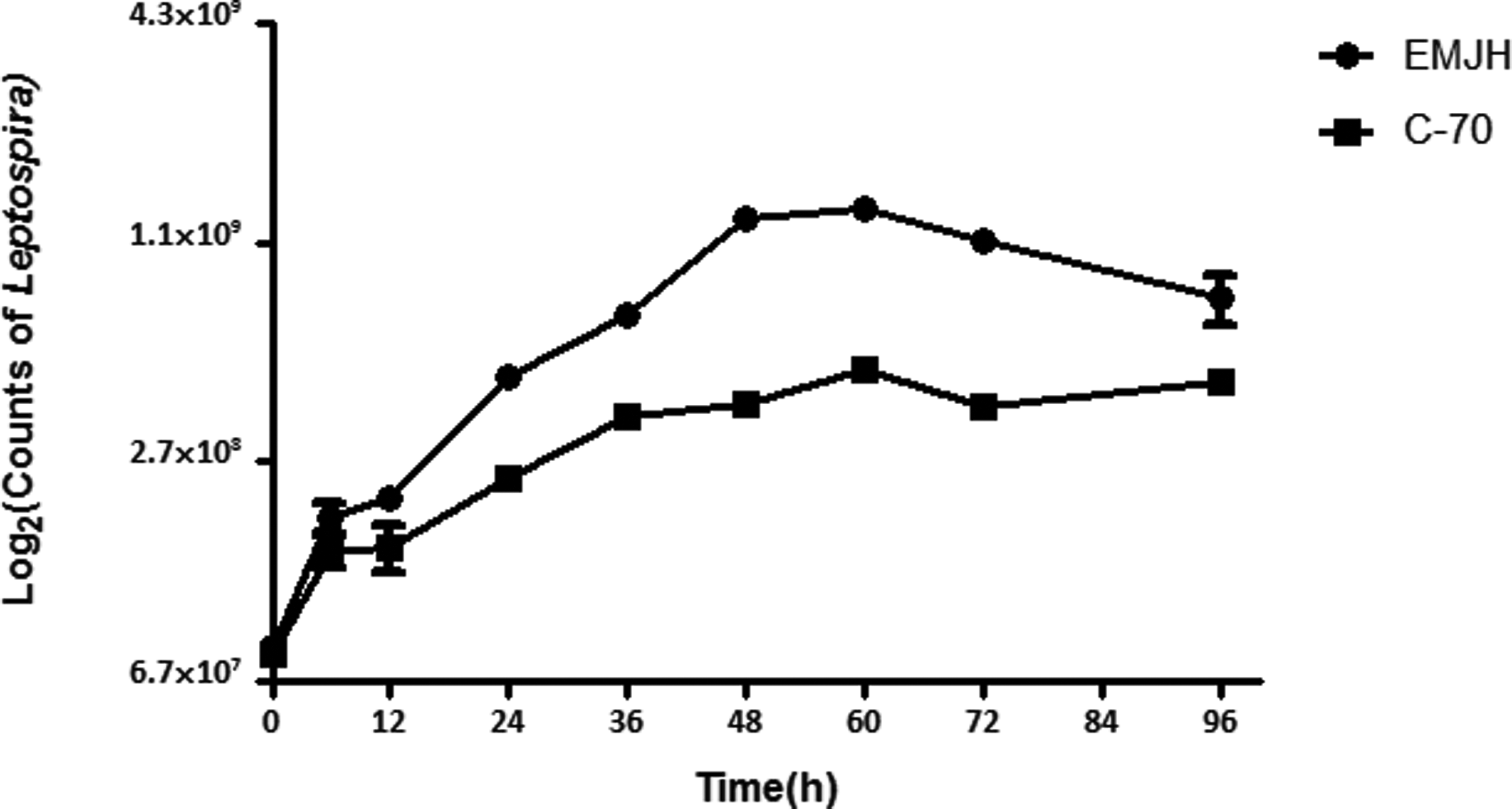
The growth curve of L. interrogans serovar Lai cultivated in protein-free medium C-70 and EMJH. Leptospira were diluted to a density of 5×107cells/mL and grown at 28°C in medium C-70 and EMJH. Triplicate samples were counted under a dark-field microscope with a Petroff-Hausser cell counter. Differences among the three groups were nonsignificant at all time points (p>0.05).

The profile of L. interrogans lysates and extracellular proteins.
LTQ MS/MS spectra obtained from the supernatant were searched against the NCBI database of strain 56601 annotations (Download from the GenBank: http://www.ncbi.nlm.nih.gov/). We selected CDSs matched by at least two unique peptides, for a total of 66 detected proteins. Compared with other gram-negative bacteria, L. interrogans secretes few extracellular protein (Viratyosin et al., 2008). Its incomplete secretion core compartment and slow growth characteristics may account for the lower number of exported protein. Nine novel CDSs that were absent from our previously published whole-cell proteomics dataset (total 2158 CDSs) from sevorar Lai were identified in this study (Supplementary Table S2) (Cao et al., 2010).
Cellular localization of the identified supernatant protein based on combined bioinformatics analysis
It has been challenging to confirm the identities of extracellular proteins. At present, Psortb, Cello, and SoSuiGramN are used to predict protein localization, while SignalP, Phobius, TMHMM, and several other programs are used to predict specific protein structures. Taken together, the results of Psortb 3.0 (Yu et al., 2010), Signal P (Bendtsen et al., 2004; Nielsen et al., 1997), Cello (Yu et al., 2004), secretome P (Juncker et al., 2003), lipoP (Juncker et al., 2003), SoSuiGramN (Imai et al., 2008), TMHMM and SpLip (Setubal et al., 2006), predicted 33 proteins to be exported to the extracellular space (Table 1).
Functional categorization of the extracellular proteome
Virulence factors
Loa22 has been confirmed as a pathogenic factor (Adler et al., 2011; Ristow et al., 2007) in L. interrogans. In our study, Loa22 was detected in the supernatant, indicating that this protein might interact with host in its extracellular form. Other potential virulence factors, such as LA_0505 and LenC, were also detected in the secretion proteome.
Host interaction proteins
One of the important roles of extracellular proteins is in host interaction during infection (Stathopoulos et al., 2000). Adhesion is one of the major processes of pathogen infection of a host. Five adhesion candidates, such as LipL32 (Hoke et al., 2008), Loa22 (Ristow et al., 2007), LenC, LenE (Stevenson et al., 2007), and LA0505 (LIC13050) (Pinne et al., 2010) were identified in our extracellular proteomics data. These proteins have all been reported as outer membrane proteins in Leptospira (Chaemchuen et al., 2011; Haake and Matsunaga, 2002; Pinne et al., 2010; Ristow et al., 2007; Stevenson et al., 2007). The detection of these proteins in the extracellular proteome further supports the hypothesis that these proteins interact directly with the host during infection.
Cellular process proteins
Three of the proteins detected in the secretion proteome have been reported to be related to Leptospira survival. The putative lactoylglutathione lyase encoded by LA_1417 (Ozyamak et al., 2010) is a component of the glutathione-dependent glyoxalase system. This system is related to bacterial survival during glycation stress. LA1953 encodes an ATP-dependent Clp protease proteolytic subunit (Marchler-Bauer et al., 2009; Marchler-Bauer and Bryant, 2004; Marchler-Bauer et al., 2011). ClpP is a potential target for modulating the presentation of protective antigens such as LLO and thereby the immune response against L. monocytogenes (Gaillot et al., 2001). It is also important for growth under stress conditions in S. typhimurium (Thomsen et al., 2002). LA_2809 encodes peroxiredoxin, which takes part in the bacterial antioxidant defense (Dubbs and Mongkolsuk, 2007; Tripathi et al., 2009). It was reported that the secreted proteins could function to help bacteria adapt to their environment (Haake and Matsunaga, 2002). These secreted cellular process-related proteins might help L. interrogans adapt to different environments.
Lipoproteins of Leptospira
Lipoproteins play important roles in the physiological and pathogenic processes in gram-negative bacteria (Kovacs-Simon et al., 2011). Lipoproteins can trigger the host inflammatory response and are important during the infection process (Schroder et al., 2008). A total of 167 lipoproteins were predicted by SpLipV1 analysis of the L. interrogans serovar Lai genome (Setubal et al., 2006). Among the exported proteins detected in our analysis, eight were lipoproteins (Supplementary Table S3). Although LipL32 is one of the most abundant outer membrane proteins in L. interrogans (Haake et al., 2000), its role in pathogenesis is still not clear. lipl32− mutant isolate showed the same virulence as the wild type, which indicates that LipL32 does not play a key role in pathogenesis (Murray et al., 2009). Expression of LipL36 is temperature dependent and is downregulated when L. interrogans is cultured at a temperature over 30°C or during the infection (Nally et al., 2001). The downregulation of LipL36 might be important for Leptospira to survive the infection process. It has been reported that LipL48 could be downregulated in response to oxygenic pressure, which might be a potential virulence factor (Xue et al., 2010). Although it is difficult to distinguish whether these three abundant proteins are contaminants or true exported proteins. The function of their extracellular forms should be studied further to determine their importance. The function of the other five lipoproteins remains unknown.
Differential transcriptional analysis of the extracellular proteins produced in vitro and in vivo
Extracellular and surface-exposed proteins are the molecules that make direct contact with host elements when the pathogens invade the host. It has thus been proposed that these extracellular proteins are the molecules that initiate many host responses (Rolando and Buchrieser, 2012; Shames and Finlay, 2012; Ustun et al., 2012). To analyze gene expression during Leptospira infection, real time PCR was used to detect the transcription levels of the 33 bioinformatically identified exported proteins (Fig. 3). The results showed that 15 of these 33 genes were upregulated, and two were downregulated in vivo. Among the 15 upregulated genes, virulence factors, including Loa22 and LipL32, were found. LA_3242 is another upregulated gene that encoded TonB-dependent receptor related to Fe2+ absorption. Since iron availability is low in the host, it presumed that upregulation of this gene might help Leptospira acquire iron and establish infection. LipL48 is one of the most abundant lipoprotein in L. interrogans. It was reported the transcriptional level of lipL48 was downregulated when Leptospira were co-cultured with phagocytes (Xue et al., 2010). However, it showed that the lipL48 was upregulated when Leptospira infected with the host. The function of LipL48 needs further study.
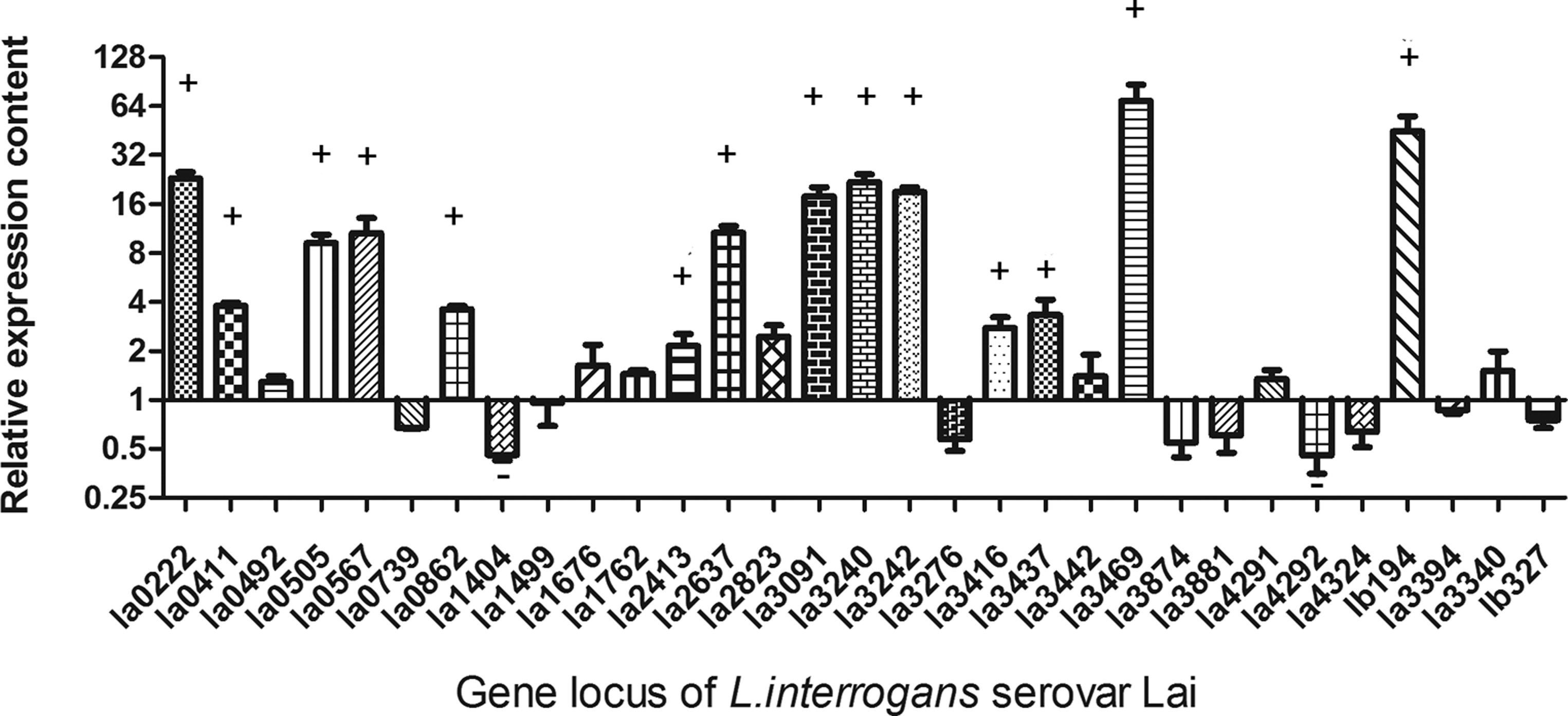
The transcriptional differences of L. interrogans serovar Lai extracellular proteins encoding genes between in vivo and in vitro. Hamsters inoculated with L. interrogans serovar Lai were used as in vivo models. Real-time PCR was used for comparing the transcriptional level between in vivo and in vitro. + means these genes were upregulated twice in vivo compared to in vitro. – means these genes were downregulated twice in vivo compared to in vitro.
Analysis of L. interrogans serovar Lai secretion systems
To the best of our knowledge, no previous study has focused on exported proteins and export-related machinery in Leptospira. Leptospira is a gram-negative bacteria that possesses an inner membrane and outer membrane. Gram-negative bacteria have evolved different methods of protein export (Tseng et al., 2009). The genome sequence of L. interrogans serovar Lai was completed in 2003, and the corresponding 4727 protein-coding sequences (CDSs) are available in GenBank (Ren et al., 2003). It was reannotated based on whole proteomics data, reducing the number of protein-coding sequences to 3718 CDSs (Zhong et al., 2011). Of these 3718 CDSs, 2158 can be detected in the high-accuracy tandem mass spectrometry (MS/MS) spectra obtained by the Yin-yang multidimensional liquid chromatography (MDLC) system coupled to an LTQ-Orbitrap mass spectrometer (Cao et al., 2010). Based on the genomic and proteomic data available for L. interrogans (Cao et al., 2010; Zhong et al., 2011), we performed an in silico analysis of the secretion machinery components of L. interrogans serovar Lai. BLASTP search for homologs of components of the secretion systems of type I, type II, type III, type IV, type V, and type VI (Cao et al., 2010; Hayes et al., 2010; Ren et al., 2003) in the NCBI database revealed presence of many secretion-related genes in the L. interrogans genome. Combined with whole cell proteomics data (Cao et al., 2010), it showed that L. interrogans possesses a relatively complete type I secretion system and type II secretion system and incomplete type III, type IV, type V, and type VI secretion systems. (Supplementary Tables S4 and S5). Schematic figures of the secretion systems in L. interrogans serovar Lai are shown in Figure 4. Furthermore, complete Sec translocase and Tat translocase systems were also found in its genome.
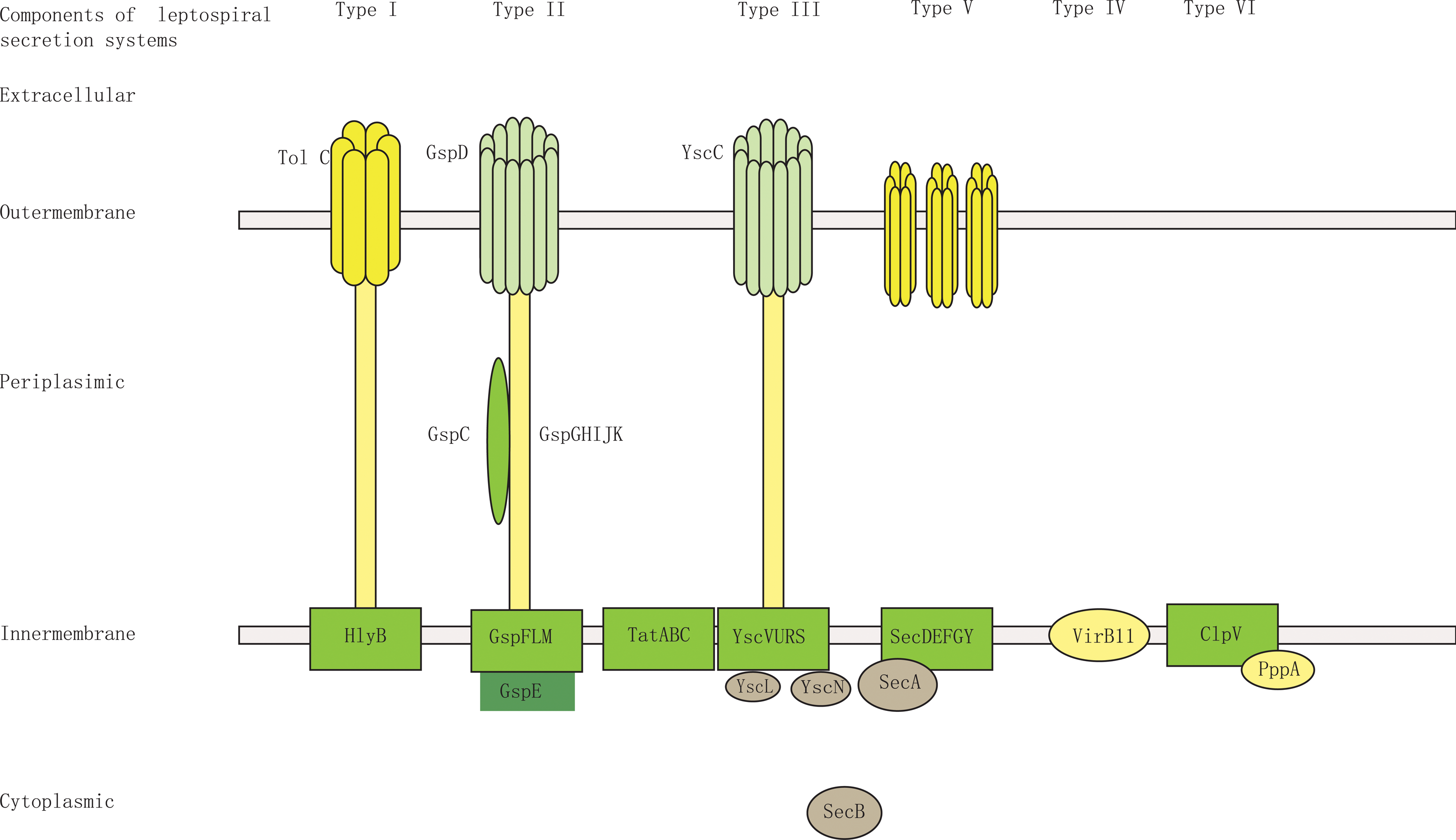
Schematic of the predicted secretion systems of L. interrogans serovar Lai. L. interrogans serovar Lai is a gram-negative bacteria that possesses an inner membrane and outer membrane. Based on the genomic and proteomic data available for L. interrogans (Cao et al., 2010; Zhong et al., 2011), BLASTP was used to search for homologs of components of the secretion system of T1SS, T2SS, T3SS, T4SS, T5SS, and T6SS in the NCBI in the L. interrogans genome and further to confirm the expression of the components by whole cell proteomics data search. This figure shows the schematic of a relatively complete T1SS and T2SS and incomplete T3SS, T4SS, T5SS, and T6SS in L. interrogans serovar Lai.
Type I secretion system
TolC genes can function with several types of transporters or alone to create trans-periplasmic channels (Saier, 2006), and the relationship between TolC channels and the type I secretion system is well defined. It has been reported that the type I secretion system can export hemolysins to the extracellular environment. Although no hemolysins were detected in our extracellular proteomic assay, there are at least 11 hemolysins in the L. interrogans genome. As L. interrogans can cause pulmonary hemorrhage, which is typically caused by hemolysins during infection, it is presumed that type I secretion system is active and functions to export the hemolysins during this process.
Type II secretion system
The general secretion system is a major pathway for the translocation of unfolded proteins across the inner membrane. Among them, the type II secretion system is broadly conserved in gram-negative bacteria that secrete enzymes and toxins across the outer membrane. Proteins secreted following this pathway contain an N-signal peptide (Rahimi and Kheirabadi, 2012). The precise assembly of type II secretion system requires a set of Gsp proteins. Nine Gsp proteins (GspD, GspC, GspF, GspG, GspJ, GspK, GspL, GspM, and GspE) have been identified in the L. interrogans genome and proteome, suggesting that this strain has a relatively complete type II secretion system. Among the 66 secreted proteins identified in our studies, 14 appeared to have signal peptides (Sec-dependent pathways) (Table 2) and thus might be exported through the type II secretion system pathway.
Twin arginine translocation systems
The twin arginine translocation (Tat) system is composed of TatA, TatB, and TatC. The Tat pathway mainly exports fully folded and assembled enzyme complexes from the cytoplasm to the periplasm in bacteria, distinct from the case in general secretion systems. The target proteins exported by the Tat pathway require a specific amino-terminal signal sequence, R-R-X-F-L-K, which is cleaved after exportation. In silico analysis revealed the presence of TatA, TatB, and TatC in the L. interrogans genome and proteome.
Nonclassical secretion pathway
The Sec- and Tat-dependent pathways are referred to as the classical secretion pathways (Bendtsen et al., 2005). In addition, there is a nonclassical secretion pathway in bacteria that is independent of the Sec and Tat pathways. Secretome P is a tool that can be used to predict whether a protein is secreted by the nonclassical secretion pathway (Bendtsen et al., 2005). In our study, Secretome P analysis indicated that 18 out of the 66 secreted proteins are predicted to be exported through the nonclassical secretion pathway (Table 3).
Conclusion
In this study, we grew L. interrogans serovar Lai in protein-free medium to obtain its set of extracellular proteins for analysis by LC/MS. Many virulence factors were detected in the supernatant of L. interrogans serovar Lai, providing new insights into the pathogenesis of leptospirosis. Furthermore, we identified homologs of several secretion system components in L. interrogans by BLAST search of the whole genome and proteome data (Cao et al., 2010). The results revealed the presence of active secretion systems and thus protein secretion capability. Many proteins have been reported to function remotely in their extracellular forms, and this might explain why Leptospira can cause severe lung hemorrhage, despite the fact that few Leptospira are detected in the host lung. Extracellular proteins are also components of the bacterial proteome. Although there might be additional secretory protein(s) not identified by the C-70 culture medium, which may only be relevant to pathogen growth in protein-rich media or during infection of the hosts. No previous global proteomic research of Leptospira has included the extracellular proteins, and these studies therefore cannot actually represent the entire Leptospira proteome. Our study might be considered as a supplement to these other proteomics analyses, especially whole proteome studies. Furthermore, it reminded us that we should also pay attention to the function of secretory proteins that are not easily detected both in vivo and in vitro.
Footnotes
Acknowledgments
This research was supported by the National Natural Science Foundation of China (81101264, 81171587, 81271793, 81201334 and 81261160321).
Author Disclosure Statement
The authors declare that there are no conflicting financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.