
Abstract
Abstract
Bacterial exoproteomes vary in composition and quantity among species and within each species, depending on the environmental conditions to which the cells are exposed. This article critically reviews the literature available on exoproteins synthesized by the foodborne pathogenic bacterium Listeria monocytogenes grown at different temperatures. The main challenges posed for exoproteome analyses and the strategies that are being used to overcome these constraints are discussed. Over thirty exoproteins from L. monocytogenes are considered, and the multifunctionality of some of them is discussed. Thus, at the host temperature of 37°C, good examples are provided by Lmo0443, a potential marker for low virulence, and by the virulence factors internalin C (InlC) and listeriolysin O (LLO). Based on the reported LLO-induced mucin exocytosis, a model is proposed for the involvement of extracellular LLO in optimizing the conditions for InlC intervention in the invasion of intestinal epithelial cells. At lower growth temperatures, exoproteins such as flagellin (FlaA) and oligopeptide permease (OppA) may explain the persistence of particular strains in the food industry environment, eventually allowing the development of new tools to eradicate L. monocytogenes, a major concern for public health.
Introduction
T
Based on the presence of somatic and flagellar antigens, L. monocytogenes strains can be classified into 13 serovars: 1/2a; 1/2b; 1/2c; 3a; 3b; 3c; 4a; 4ab; 4b; 4c; 4d; 4e; and 7 (Farber and Peterkin, 1991). Although strains from all the serovars can cause human infection (Seeliger and Jones, 1986), epidemiological data show that the majority of human cases are associated with serovars 1/2a, 1/2b and 4b (Chenal-Francisque et al., 2011; Goulet et al., 2008; Schuppler and Loessner, 2010). DNA-based subtyping methods have established in L. monocytogenes at least four genetic evolutionary lineages correlated with the serovars (Datta et al., 2012; Orsi et al., 2011; Ward et al., 2008). Lineage I (flagellar antigen types b and d) contains strains that are more likely to cause human disease than the isolates classified in lineages II (flagellar antigen types a or c), III and IV (some strains of 4b and the rarely detected serovars 4a and 4c). The strains within lineage II are usually present in food and food environments more frequently than lineage I isolates (Nucera et al., 2010; Orsi et al., 2011; Parisi et al., 2013; Wang et al., 2012). However, some reports indicate the prevalence of serovar 4b (lineage I) in food and food-related isolates (Leite et al., 2006; Pagadala et al., 2012).
Listeria monocytogenes is a well-adapted saprophytic microorganism. The survival of this bacterium under diverse environmental stresses strongly contributes to the niche persistence of some strains. The persistence of L. monocytogenes in food-processing facilities has been described in great detail (D'Amico and Donnelly, 2008; Ferreira et al., 2011; Fox et al., 2011; Keto-Timonen et al., 2007; Leite et al., 2006; Orsi et al., 2008). These strains become established for months or years as part of the resident microbiota, suggesting that they are capable of withstanding and overcoming the significant challenges imposed, such as cold, unfavorable pH, and salt stress. The increased diversity in consumer preferences for food products has led to the widespread implementation of cold chains to meet food safety demands (Kuo and Chen, 2010; Rediers et al., 2009). The adaptability of Listeria to low temperature combined with food storage at improper conditions may lead to significant growth of the pathogen within food matrices (Rosset et al., 2004). To such low temperatures, different strains of L. monocytogenes respond differently (Kagkli et al., 2009; Moorhead and Dykes, 2004; Pal et al., 2008).
The survival of a bacterial pathogen depends on its ability to maneuver and react actively with host cells. This host–pathogen interaction during listeriosis has been extensively studied (Cossart et al., 2003; Pizarro-Cerdá et al., 2012; Posfay-Barbe and Wald, 2009; Stavru et al., 2011; Vazquez-Boland et al., 2001). Over several years, these studies have established that L. monocytogenes promotes its internalization into the host epithelial cells via an interaction between the bacterial surface molecules internalin A (InlA) and internalin B (InlB), and the host cellular receptors epithelial cadherin (Ecad) and mesenchymal epithelial transition factor (Met), triggering several complex host cell pathways that lead to bacterial engulfment and escape from recognition by the innate immune system. In the subsequent stages of infection, the escape from the phagocytic vacuole is listeriolysin O (LLO)-dependent, and intracellular actin-based motility and cell-to-cell spread are actin A (ActA)-dependent (Fig. 1A). Cellular infection studies performed in vivo revealed that transcytosis could be responsible for L. monocytogenes early, rapid and efficient intestinal barrier cross. In fact, L. monocytogenes enters intestinal epithelial cells after InlA-Ecad interactions and, in a LLO and ActA-independent manner, crosses the epithelial cells by a furtive intra-vacuolar path (Fig. 1B) (Nikitas et al., 2011).
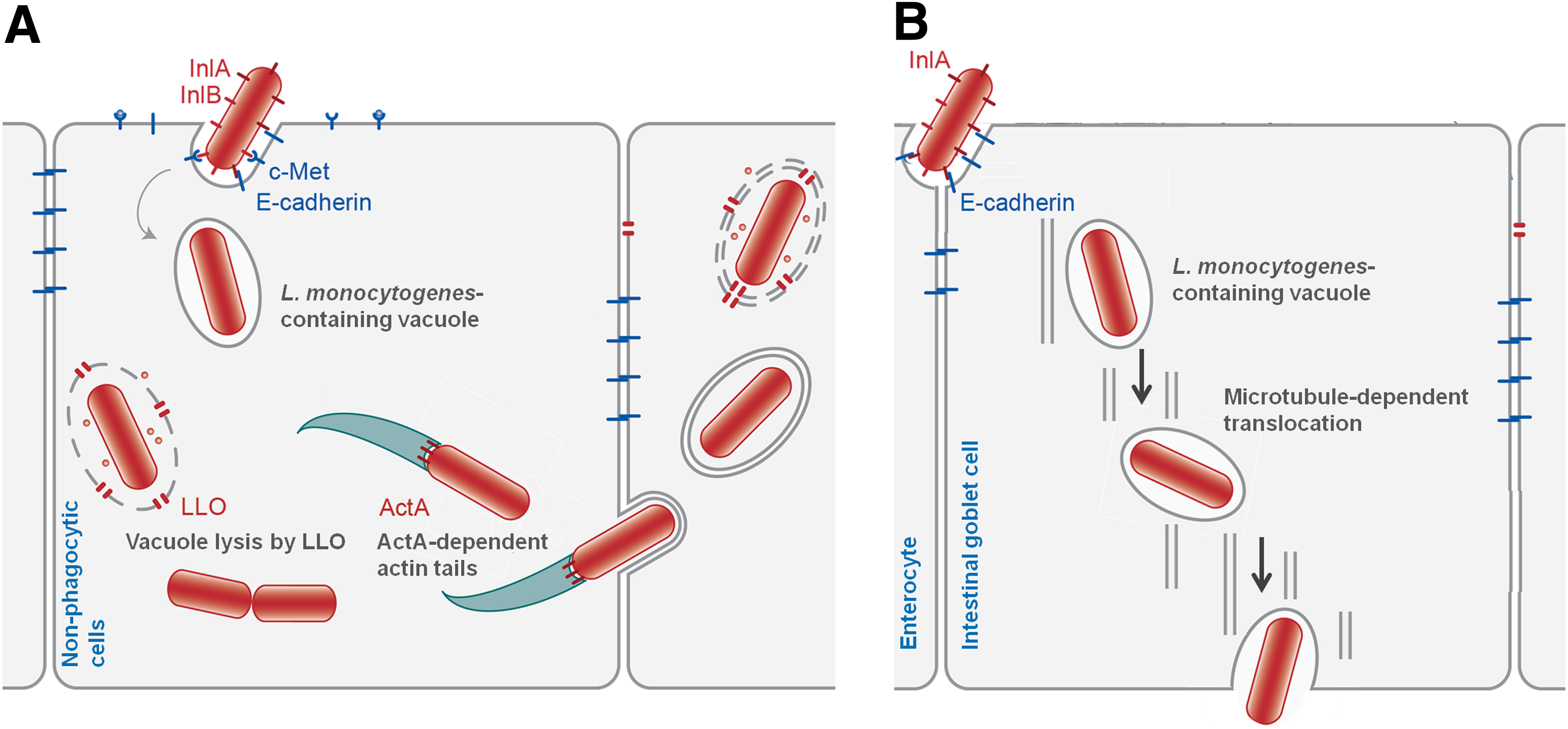
Listeria monocytogenes host-cell invasion.
L. monocytogenes can switch between life in soil and in the mammalian host through complex regulatory pathways that modulate the expression of factors involved in pathogenicity and/or in the adaptation to different abiotic life styles (Freitag et al., 2009; Gray et al., 2006; Toledo-Arana et al., 2009). Positive regulatory factor A (PrfA), class three stress gene repressor protein (CtsR), heat-inducible transcription repressor (HrcA), and the alternative sigma factors σB, σC, σH and σL are among the regulators that play determinant roles in the bacterial stress response and adaptation to the environment, including gastrointestinal survival and the systemic stages of infection (Chaturongakul et al., 2008; 2011).
In the context of host–pathogen interactions, a number of proteins that are secreted are important in evading and availing host cell defenses. The bacterial extracellular proteome or exoproteome undergoes multiple adaptations according to the bacterial environmental conditions (e.g., degradative enzymes during saprophytic conditions, as opposed to virulence factors when inside the host) (Conte et al., 2000; Lee and Schneewind, 2001; Midelet-Bourdin et al., 2006). Furthermore, the expression patterns of the exoproteome can undergo more pronounced alterations than the cytoplasmic proteins (Dumas et al., 2008, 2009b). Schliep et al. (2012) proposed that it is unnecessary to analyze all the proteins in a proteome and that a smaller subset of outer membrane proteins is sufficient for the elucidation of environmental patterns.
In this review, recent studies on the L. monocytogenes exoproteome are summarized and discussed. The bacteria are repeatedly exposed to the host temperature or the low temperatures used for food preservation. Therefore, the interstrain exoprotein diversity may explain the particularly high virulence of some strains and the persistence of some others in the food environment.
The Exoproteome
Successful infection by L. monocytogenes requires diverse adaptation strategies to efficiently avoid (by abolishing, counterattacking, and/or circumventing) the host defense mechanisms; this process is dependent on specific virulence factors such as extracellular proteins, which play an important role in pathogenicity (Desvaux and Hebraud, 2006; Fuchs et al., 2012; Trost et al., 2005; Vazquez-Boland et al., 2001). Emergence of powerful analytical equipments for mass spectrometry has enabled the complete analysis of these proteins, which can provide insights into pathogenesis and therapeutics (Windle et al., 2010), in a global effort to understand and eradicate listeriosis. An important feature of such studies is the comparison between virulent and less virulent strains, and the identification of new key virulence factors. In this respect, the comparison with the closely related species L. innocua, motivated several researchers to screen for new potential virulence factors.
Based on the analysis of the genome for sequences that encode proteins, Glaser et al. (2001) predicted that 86 proteins were secreted into the external milieu by L. monocytogenes. Trost et al. (2005) identified 45 putative secretory proteins from L. monocytogenes grown at 37°C. This study experimentally demonstrated that almost half of the extracellular proteins identified had no recognizable signal sequence for secretion, which meant that their extracellular location could not be predicted from their genomic sequences. Furthermore, the discrepancy between the expected and observed proteins in the extracellular milieu led to the proposal that this set of proteins should be called the “exoproteome” or “extracellular proteome”, instead of “secretome” (Desvaux et al., 2009). In fact, an actively secreted protein could be entirely outside of the cell and can exist as a soluble (free) protein, a surface-associated protein, or a subunit part of a surface appendage. In the extracellular milieu, there are actively secreted proteins, but also proteins released after cell lysis or burst membrane vesicles budding, which cannot be considered as secretion systems sensu strict (Desvaux et al., 2009). The exoproteome of a bacterium is actually composed by all of the proteins found in its culture supernatant.
Main challenges in exoproteome analysis
The typical low concentration of proteins in bacterial culture supernatants constitutes the main challenge in the analysis of cell exoproteomes and requires the use of more sensitive techniques. Table 1 includes 34 of the exoproteins present in the supernatants of 37°C-cultures of L. monocytogenes strains EGD-e and 10403S, grouped by their main functional categories. These proteins were identified by Trost et al. (2005) in the first comprehensive post-genomic study of the L. monocytogenes exoproteome. Subsequently, other groups have confirmed the presence of the same proteins. Trost et al. (2005) combined predictive bioinformatics and two proteomic approaches: 1) two-dimensional gel electrophoresis (2D-GE), followed by matrix assisted laser desorption ionization–time of flight–mass spectrometry (MALDI-TOF-MS) peptide mass fingerprint and post source decay (PSD) fragmentation; 2) high performance liquid chromatography/electrospray ionization-mass spectrometry (LC-MS). Based on these two approaches, respectively 58 and 47 exoproteins were identified, resulting in a total of 105 exoproteins. Dumas et al. (2009a) used 2D-GE and MALDI-TOF-MS to compare the exoproteomes of 12 L. monocytogenes strains, including EGD-e, but only identified 60 exoproteins. The virulence factor Mpl was one of the exoproteins identified by Trost et al. (2005) (Table 1), but not by Dumas et al. (2009a).
Member of the variant exoproteome of L. monocytogenes (Dumas et al., 2009a).
Protein underexpressed in the strains from serovar 1/2a, relative to strains from serovars 1/2b and 4b (Dumas et al., 2008).
Protein overexpressed in the strains from serovar 1/2a, relative to strains from serovars 1/2b and 4b (Dumas et al., 2008).
Protein absent or downregulated in a L. monocytogenes mutant with constitutive expression of prfA gene (Port and Freitag, 2007).
Combination of different experimental approaches for extracellular protein separation and identification has therefore resulted in a larger number of proteins identified. However, differences in strain response to environmental conditions have not yet been fully elucidated. The simultaneous use of different techniques increases the time and cost of the study and does not always reflect better discrimination of strain behavior. Further studies focusing on a larger number of L. monocytogenes strains with different degrees of virulence and distinct abilities to persist as in-house strains will allow a better understanding of how this species interacts with its milieu.
Strategies to overcome the major constraints of exoproteome analyses
One successful strategy that may be used to increase the relative amount of each protein, and therefore the probability of exoprotein detection and identification, is reducing sample complexity. In fact, the L. monocytogenes exoproteome can be divided into different subsets of exoproteins. The most important and well-studied subset contains virulence factors (Chaturongakul et al., 2008; 2011), which may be located in the bacterial cell surface to promote host–cell interactions. Schaumburg et al. (2004) established a methodology for the isolation of a low complexity surface proteome and identified 55 proteins by N-terminal sequencing and MS, after serial extraction and 2D-GE. These authors reported the difficulty in detecting and identifying cell wall proteins covalently linked by the LPXTG motif, most likely because of their tight linkage to peptidoglycans and/or their low levels of expression. Calvo et al. (2005) analyzed the complex peptide mixture obtained from a preparation of peptidoglycan-enriched material by a nonelectrophoretic approach such as 2D nano-LC-MS/MS. Nineteen different proteins were identified for the L. monocytogenes EGD-e strain (some of which are indicated in Table 1), showing that this highly sensitive method was effective for studying protein–peptidoglycan associations. This same approach was successfully used by other authors to study surface proteomes of L. monocytogenes (Calvo et al., 2005; Huang and Hussain, 2012; Nilsson et al., 2013; Pucciarelli et al., 2005; Zhang et al., 2013) and of other microorganisms (Armengaud, 2013). Gel-free approaches were also used, with prior separation of the proteins based on their isoelectric point (Habicht et al., 2011).
Recently, nonlabeling quantitative approaches have gained prominence over 2D-GE methods in most proteomic studies, although the 2D-GE methods are still commonly used (Armengaud, 2013; Porteus et al., 2011). The high-throughput identification of proteins by LC-MS/MS following trypsin proteolytic cleavage has been widely used to study the protein complement expressed under certain conditions by a specific microorganism. Typically, the protein sample under study is subjected to SDS-PAGE and the fractionated polypeptides are subsequently prepared for high-throughput identification (Clair et al., 2010; Wijte et al., 2010).
Another approach that has been used to overcome difficulties in exoproteome analysis is based on the use of mutant strains. Port and Freitag (2007) found that the use of mutationally activated prfA alleles was a useful approach to study the role of proteins Lmo0135 and TcsA, which had not been previously associated with bacterial virulence (Table 1). This approach, which can be extended to other regulators of virulence factors as shown by Abram et al. (2008) and Mujahid et al. (2013), will certainly aid in the discovery of new exoproteins associated with virulence.
The importance of “multi-omics” studies in integrative biology is also very well documented in the work described by Bowler et al. (2013) that identified an association between the levels of the multifunctional protein cytokine IL-16 and emphysema. These integrative “omics” approaches, will help in identifying key proteins to be used as relevant biomarkers in public health.
Comparison of the exoproteins of distinct Listeria strains
Trost et al. (2005) identified 16 proteins in the culture supernatant of L. monocytogenes strain EGD-e that have no orthologs in the nonpathogenic bacterium L. innocua. Nine of these proteins (PlcA, LLO, Mpl, ActA, PlcB, InlA, InlB, InlC, and InlH) were virulence factors (Table 1). These authors also found 43 proteins that were specifically expressed by L. monocytogenes but not by L. innocua. Similarly, a comparative proteomic analysis of the cell wall subproteomes in pathogenic and nonpathogenic Listeria identified the molecular components associated with virulence in L. monocytogenes (Calvo et al., 2005). The mutational activation of the central virulence regulator PrfA allowed the identification of novel potential extracellular virulence factors (Port and Freitag, 2007). These important exoproteomic approaches were based on the study of a single L. monocytogenes strain, generating conclusions that are insufficient to be generally applied to the species. Dumas et al. (2008) compared the extracellular proteins of 12 strains belonging to the serovars 1/2a, 1/2b, and 4b, and concluded that the abundance or scarceness of specific proteins was related to the genetic lineage of the strains. These authors also found proteins that were not considered potential virulence factors by Trost et al. (2005). These results suggest that the environmental conditions and stimuli required for the expression of virulence factors may vary by strain.
The importance of cell wall proteins in strain discrimination
Trost et al. (2005) showed that the “cell surface proteins and metabolism of the cell wall” was the functional category that contributed most to the differences between L. monocytogenes and L. innocua, followed by proteins in the category of “specific virulence factors.” It is well known that the degree of pathogenicity of L. monocytogenes under the experimental conditions tested (i.e., its virulence potential) differs among isolates. Differences in the virulence potential of the strains within each serovar of the species have been reported, despite the association of most human cases with serovars 1/2a, 1/2b, and 4b (Cabrita et al., 2004; Dumas et al., 2009b; Neves et al., 2008; Olier et al., 2003). Dumas et al. (2008) performed a proteomic analysis of L. monocytogenes strains from these three serovars and found that the proteins responsible for different levels of virulence were mainly from the “cell surface proteins and metabolism of the cell wall” functional category. These results revealed the importance of this protein set in strain discrimination. A more detailed analysis indicated that more than half of the non-orthologous proteins identified in L. monocytogenes were commonly found in the extracellular milieu of all strains (core exoproteome), whereas 17 proteins (variant exoproteome) appeared to be expressed only in some L. monocytogenes strains with different levels of virulence, as PlcA in strain EGDe (Table 1). This suggests the existence of partially strain-specific post-translational modifications, as reported for some virulence factors (Dumas et al., 2009a). However, none of these proteins was considered by these authors as a virulence marker (Dumas et al., 2008; 2009a).
One of the proteins of the “cell surface proteins and metabolism of the cell wall” functional category is Lmo0443, a putative cell-wall protein (Table 2). This protein belongs to the core exoproteome of L. monocytogenes and is differential expressed, as is described below.
Studies on low-virulence strains
It is generally assumed that all L. monocytogenes strains are strictly pathogenic, but some questions about the virulence in this species remain unanswered. For instance, nonvirulent or weakly virulent strains of L. monocytogenes have been reported in the intestine of 1%–15% healthy individuals among the population (Grif et al., 2003; Olier et al., 2003; Rocourt et al., 2000; Velge and Roche, 2010). According to some authors, the prevalence of low-virulence strains isolated from different environmental sources could be greater than 60% (Roche et al., 2009; Velge and Roche, 2010). Mutations in key virulence genes, including inlA, inlB, plcA, plcB, hly, or actA, have been detected in these strains (Holch et al., 2010; Nightingale et al., 2008; Roberts et al., 2005; Roche et al., 2012; Temoin et al., 2008; Van Stelten et al., 2010). The transcriptional regulators of L. monocytogenes virulence genes, such as PrfA, can also be affected by mutations (Roche et al., 2012; Velge et al., 2007) influencing gene expression and most likely compromising extracellular protein expression. Comprehensive studies showed that the diversity and population structure of L. monocytogenes according to the virulence level is complex and based on different mechanisms, which seem to differ according to the genetic lineages of the strains (Roche et al., 2012). There are few studies on the differential extracellular proteomic expression and/or differential gene expression between virulent and low-virulence strains of L. monocytogenes (Duodu et al., 2010; Olier et al., 2002; Olier et al., 2003). Cabrita et al. (2010) performed comparisons on the exoproteins from virulent and low-virulence strains, belonging to genetic lineages I, II, and III, grown in minimal medium at 37°C and found significantly more InlC and LLO proteins in the culture supernatants of the virulent strains (Table 2). The results obtained indicated the absence of correlation between these exoproteins and the genetic lineage of the strains and emphasized the need for the inclusion of low-virulence strains in proteomic studies, to gain new insights into the expression of virulence factors.
Key exoproteins produced at 37°C
Lmo0443, a protein with a putative role in cell-wall structural maintenance
Protein Lmo0443 (Tables 1 and 2) is similar to the Bacillus subtilis transcription regulator LytR, which belongs to the LytR/CpsA/Psr family and has a putative role in cell-wall structural maintenance, possibly through autolysin regulation (Chatfield et al., 2005). The members of this family consist of membrane-anchored proteins with a short cytoplasmic N-terminal sequence, a transmembrane helix, and a large extracellular region containing the LytR/CpsA/Psr domain. Although the main function of these proteins remains unclear, they are known to influence the clinically relevant attributes of various gram-positive pathogens, such as cell division and septum formation (Chatfield et al., 2005; Hübscher et al., 2008; Johnsborg and Havarstein, 2009; Over et al., 2011). Recently, Minami et al. (2012) found that the lytR mutant strain of the gram-positive bacterium Streptococcus pyogenes had a higher activity of the cysteine protease SpeB and was more virulent than the wild-type strain. Consistent with the results of Dumas et al. (2008), the LytR protein of S. pyogenes (25% similar to the Lmo0443 protein of L. monocytogenes) seems to play an important role in the virulence of S. pyogenes.
The transcription of lmo0443 is controlled by CesRK, an antibiotic-sensing two-component system in L. monocytogenes, and is upregulated by cell wall-targeting antibiotics such as ampicillin, vancomycin, cefuroxime (Gottschalk et al., 2008; Kallipolitis et al., 2003; Nielsen et al., 2012), amoxicillin, penicillin, and teicoplanin (Knudsen et al., 2012). Knudsen et al. (2012) also tested protein- and DNA/RNA-synthesis-targeting antibiotics and found that lmo0443 expression is downregulated by tetracycline and upregulated by co-trimoxazole. The gene encoding Lmo0443 has an ortholog in L. innocua; however, it is only expressed in L. monocytogenes (Schaumburg et al., 2004; Trost et al., 2005). The expression of lmo0443 is co-regulated by PrfA and σB; in the absence of σB, PrfA suppresses the transcription of lmo0443, whereas in the presence of σB, PrfA promotes the transcription of lmo0443 (Chaturongakul et al., 2011; Ollinger et al., 2009). Lmo0443 is generally overexpressed in less virulent strains and underexpressed in more virulent ones (Dumas et al., 2008) (Table 2), and can be seen as a potential marker for low virulence.
Pleiotropic effects of LLO
LLO, a member of the cholesterol-dependent cytolysin (CDC) family of bacterial toxins, was first described as essential for the intracellular escape of L. monocytogenes from the internalization vacuole and subsequent growth within infected cells (Gaillard et al., 1987; Sun et al., 1990) (Table 2). LLO is positively regulated by PrfA (Leimeister-Wachter et al., 1990). LLO is also required for the escape of L. monocytogenes from the double-membrane vacuole that is formed during cell-to-cell spreading (Dancz et al., 2002; Gedde et al., 2000) (Fig. 1). LLO is not only a major virulence and immunogenic determinant in L. monocytogenes infection but also a highly cytotoxic protein (Carrero et al., 2012). LLO is the only known CDC toxin whose activity is regulated by pH (Hamon et al., 2012). Inside the vacuole, the low pH required for LLO activity, which was estimated at 5.5, restrains the toxin activity to acidic vesicles (Glomski et al., 2002; 2003). Under cytosolic conditions of neutral pH, LLO spontaneously forms aggregates, preserving host cell components from damage; this constitutes the major mechanism of inactivation of this toxin (Bavdek et al., 2012). In addition to its role as a pore-forming toxin, LLO has several other functions inside host cells, which may be equally important for infection, such as the control of autophagy upon escape from the double-membrane vacuoles and the suppression of the key host mechanisms that reduce pathogen replication (Hamon et al., 2012).
Several studies revealed that extracellular LLO plays an important role before the bacteria enter the host cell, which is consistent with the observation that this important virulence factor can be detected in the supernatants of 37°C-cultures of this pathogen (Cabrita et al., 2010; Moors et al., 1999; Trost et al., 2005). LLO acts as a signaling molecule in different host cell pathways such as the induction of bacterial internalization and early induction of autophagy, modulation of host immune response, and actions on host cell organelles (Hamon et al., 2012; Kayal and Charbit, 2006; Stavru et al., 2011). It was recently reported that LLO secreted by L. monocytogenes activates the unfolded protein response of the transmembrane and secretory proteins by the host endoplasmic reticulum, which is another example of the bacterial manipulation of the host defense strategies (Pillich et al., 2012). However, despite these extensive pleiotropic effects of LLO, the recently discovered process of transcytosis of L. monocytogenes across the intestinal barrier was described as LLO-independent (Fig. 1B) (Nikitas et al., 2011).
The intestinal mucosal epithelial cells are covered by a protective mucus layer, mainly composed of mucins (Fig. 2A). The mucins of this constantly renewed layer act as receptors for pathogenic bacteria and may represent an important factor in host–pathogen interactions (Linden et al., 2004; 2008a). During the initial stages of infection before bacterial cell invasion, extracellular LLO induces mucin exocytosis from the intestinal epithelial cells (Fig. 2B) and upregulation of the MUC genes encoding membrane-bound mucins, which is considered a host cell defense response against pathogen invasion (Lievin-Le Moal et al., 2005). In Campylobacter jejuni and Salmonella enterica, the presence of the MUC2 appears to enhance virulence (Alemka et al., 2012; Gagnon et al., 2013). MUC2 is the main secreted mucin in the protective layer of mucosal epithelial cells. In C. jejuni, the presence of MUC2 causes the upregulation of virulence genes, among which are those involved in motility, adhesion, invasion, and toxin formation (Alemka et al., 2012). However, regardless of the method of invasion (classical or transcytosis) used by L. monocytogenes, the LLO-induced mucin exocytosis can be a host defense mechanism exploited by the pathogen to enter epithelial cells, optimizing the conditions in the intestinal lumen for InlC intervention (Fig. 2B).
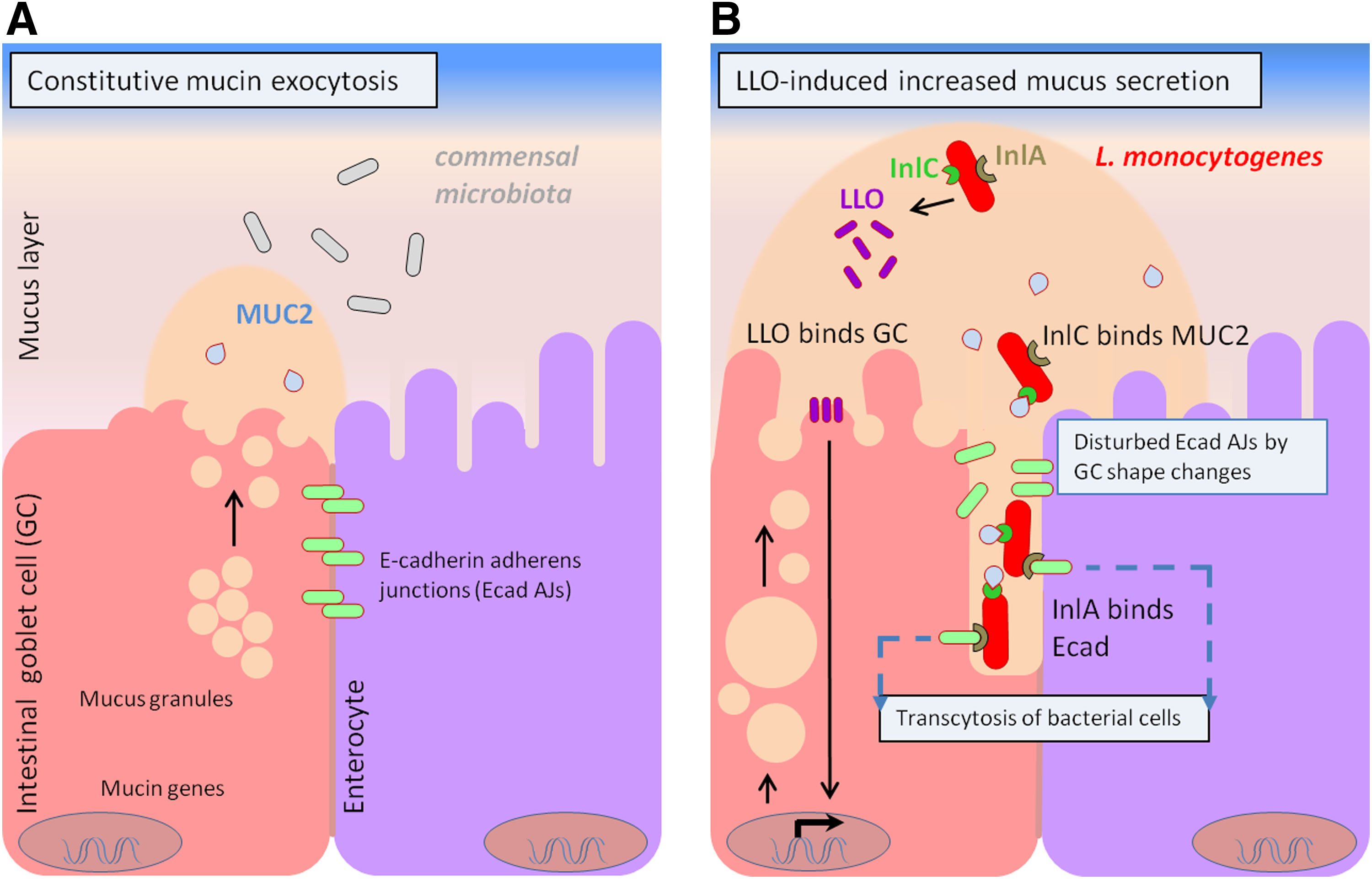
The role of LLO and InlC in Listeria monocytogenes infection by transcytosis.
InlC, a multifunctional virulence factor
InlC is a major protein target of the human humoral immune response against L. monocytogenes (Table 2) (Grenningloh et al., 1997). The inlC gene is not present in all strains of L. monocytogenes (Tsai et al., 2006; 2011). Previous studies have found it to be present in lineages I and II, but absent in some strains from lineage III (Doumith et al., 2004; Tsai et al., 2006). Doumith et al. (2004) even proposed that the differentiation of strains belonging to the serovars 4a and 4c was based on the lack of inlC gene in the 4a strains. However, Liu et al. (2007) analyzed two serovar 4c strains for the presence of inlC and reported that the gene was absent in one strain (ATCC 19116). This result was confirmed by Cabrita et al. (2010), who reported the absence of InlC in the supernatant of this strain grown in minimal medium at 37°C.
InlC belongs to a large family of proteins known as internalins, which contain an “internalin domain” comprised of a leucine-rich repeat (LRR) domain flanked by an immunoglobulin-like (IR) domain (Ooi et al., 2006). In some internalins, the LRR domain is involved in binding to a receptor during host–pathogen interactions (Dramsi et al., 1997; Schubert et al., 2002), whereas the IR domain might constitute a potential receptor-binding site (Ooi et al., 2006). Several observations support the importance of InlC in host infection, including some structural features of the protein (Ooi et al., 2006) and the fact that the transcription of inlC is positively regulated by PrfA at 37°C but is not regulated by σB (McGann et al., 2007a; 2007b; 2008; Port and Freitag, 2007). The inlC gene is present in pathogenic but not in nonpathogenic Listeria species (Domann et al., 1997; Doumith et al., 2004; Trost et al., 2005). Deletion mutations of inlC cause reduced virulence in a mouse infection model (Domann et al., 1997; Engelbrecht et al., 1996). The intracellular overexpression of the inlC gene at the late stages of infection suggests that InlC might play a role when bacteria are in the host cell cytoplasm and during intercellular bacterial spread (Chatterjee et al., 2006; Joseph et al., 2006). Recent studies showed that InlC is overexpressed and secreted into the host cytosol after bacterial escape from the internalization vacuole and attenuates the effects of the host innate response induced during the infection process (Gouin et al., 2010). InlC also promotes efficient actin tail motility to facilitate the well-known cell-to-cell spread during infection (Rajabian et al., 2009; Romero and Van Nhieu, 2009).
However, the potential of L. monocytogenes to cause disease most likely correlates initially with its capacity to survive and remain within the host intestinal lumen, with InlC operating as an important factor during the first stages of host–pathogen interaction. Some results suggest that during the later stages of host–cell infection, InlC might play a supportive role in the InlA-mediated internalization of L. monocytogenes by nonphagocytic cells (Bergmann et al., 2002). Moreover, some studies showed that InlC, along with InlJ and InlB, binds specifically to surface mucin MUC2 (Fig. 2B) through its LRR-IR domain (Linden et al., 2008a; 2008b). These studies showed that the interactions between InlC and MUC2 can contribute to one of two effects. On one hand, MUC2 might inhibit the interactions between L. monocytogenes and the intestinal epithelial cells, reinforcing the previously proposed hypothesis that the LLO-induced mucin exocytosis constitutes a host cell defense response that inhibits the cell entry of L. monocytogenes (Lievin-Le Moal et al., 2005). On the other hand, the interaction between MUC2 and L. monocytogenes might fix the bacterial cells within the intestinal mucus in a step preceding the crossing of the mucosal barrier and attachment to the intestinal epithelial cells. This might be a way of exploiting the host defense mechanisms. The results obtained by Nikitas et al. (2011) on transcytosis are consistent with the second hypothesis. During the transcytosis mode of infection, mucin production by the goblet cells enables the L. monocytogenes invasion of the host cells, which demonstrates the relationship between mucus secretion and luminal access to the Ecad expressed by human intestinal epithelial cells. These observations suggest that the LLO-induced mucin exocytosis reported by Lievin-Le Moal et al. (2005) might indeed represent an exploitation of the host defense mechanisms by L. monocytogenes to enter epithelial cells. Therefore, the L. monocytogenes strains that express higher levels of the LLO and InlC virulence factors potentially have a competitive advantage over strains that express lower levels of these proteins, either during the classical infection cycle (Fig. 1A) or the recently described transcytosis pathway (Fig. 1B and Fig. 2B). Therefore, current evidence suggests that InlC, such as LLO, is a multifunctional virulence factor, playing a role in host infection from the earliest stages of bacterial colonization of the intestinal lumen to the later intracellular and cell-to-cell spreading stages.
Dumas et al. (2008) found that L. monocytogenes strains from serovars 1/2a, 1/2b, and 4b grown at 37°C produced InlC, but at different levels. The InlC protein was considered a component of the core exoproteome of 12 L. monocytogenes strains from those three serovars (Dumas et al., 2009a). However, other studies using virulent and low-virulence strains suggested that the higher levels of InlC found in the exoproteomes of the virulent strains are directly related to their virulence potential (Cabrita et al., 2010).
Key exoproteins produced at low temperatures
At low temperatures, microbes must acclimatize to the cold stress environment by reducing their growth rate and altering, in a tightly regulated manner, gene expression. Several cold stress adaptation mechanisms have been investigated in L. monocytogenes, some of which are related to cell membrane functions. Some of these mechanisms seem to involve the uptake of short peptides from the growth medium and specific modifications in cell-surface protein expression, suggesting the potential contribution of extracellular proteins in the response of bacteria to low temperatures; this contribution is not completely understood (Tasara and Stephan, 2006). Changes in the expression of several cytoplasmic proteins were detected in a strain of L. monocytogenes grown at low temperature (Cacace et al., 2010). However, there are few studies comparing the exoproteomes of more than one L. monocytogenes strain grown at low temperatures. Recently, Cabrita et al. (2013) showed that the exoproteomes of four genetically different L. monocytogenes strains grown at 11°C and 20°C are significantly different from the exoproteomes obtained at the host temperature of 37°C (Cabrita et al., 2010). These authors also found major differences in exoproteins detected among the strains grown at low temperature. The main findings were the lack of FlaA and OppA in the exoproteome of a persistent strain, along with differences in the levels of these proteins among the other strains. Furthermore, the levels of similarity of the polypeptide profiles of the strains decreased with growth temperature, suggesting the increased differentiation of the strain response at low temperatures (Cabrita et al., 2013). These results further highlight the importance of studying the bacterial exoproteomes.
Flagellin (FlaA)
Tasara and Stephan (2006) reported that changes in the expression of cell membrane/surface proteins induced by temperature shifts are involved in adaptation to cold stress. L. monocytogenes strains are motile and flagellated at low temperatures (30°C and below), with up to four peritrichous flagella per cell (Griffin and Robbins, 1944), but typically not motile at the host temperature (37°C) (Way et al., 2004), at which production of flagellin, encoded by the flaA gene, is markedly reduced (Cabrita et al., 2010; Dons et al., 1992; Peel et al., 1988) (Table 3). Because bacterial flagellin triggers the activation of the innate immune response (Gewirtz et al., 2001; Hayashi et al., 2001), it was proposed that the downregulation of flaA expression during in vivo infection by L. monocytogenes might serve as an adaptive mechanism to avoid host recognition and mobilization of the host innate immune responses (Dons et al., 2004; Way et al., 2004).
Earlier studies showed that flagellin expression and flagellar assembly vary broadly among strains (Grundling et al., 2004; Peel et al., 1988; Schirm et al., 2004). Cabrita et al. (2013) showed that the detectable differences in flagellin amounts in the exoproteomes of different L. monocytogenes strains corresponded to differences in the level of flagellation of the strains (Cabrita et al., 2013).
Several studies demonstrated that biofilm formation is responsible for the persistence of some strains in food-processing environments (Keto-Timonen et al., 2007; Lomonaco et al., 2009; Wulff et al., 2006) and flagella are considered mediators in bacterial attachment to different synthetic surfaces (Caly et al., 2009; Lemon et al., 2007; Vatanyoopaisarn et al., 2000). Recently, flaA mutants were found to be defective in biofilm formation (Chang et al., 2012; Ouyang et al., 2012). de la Fuente-Nunez et al. (2012) reported that a specific small cationic peptide induced the underexpression of some bacterial flagellar genes in P. aeruginosa, suggesting that flagella have an important role in biofilm formation. However, mutant strains of L. monocytogenes lacking flagella or flagellar mobility displayed a hyperbiofilm phenotype (Todhanakasem and Young, 2008). Other results (Cabrita et al., 2010; 2013) showed that the planktonic cells of a persistent strain of L. monocytogenes (Leite et al., 2006) possessed a few flagella at 11°C and failed to present extracellular flagellin after growth at 11°C, 20°C, or 37°C.
Oligopeptide permease A (OppA)
The uptake of short peptides by L. monocytogenes is mediated by two cell membrane systems: a proton motive force-dependent system that mediates the transport of di- and tripeptides, and the oligopeptide permease (Opp) transport system, which mediates oligopeptide transport to the bacterial cell (Verheul et al., 1995; 1998) (Table 3).
The gene oppA that encodes the protein OppA belongs to the opp operon, which contains four other genes, oppB, oppC, oppD, and oppF. However, due to the operon organization, oppA is predominantly transcribed as a monocystronic mRNA (Borezee et al., 2000). During growth at low temperature, the transcription of oppA was significantly reduced in a L. monocytogenes ΔsigL mutant compared to the wild-type strain (Raimann et al., 2009), suggesting the involvement of the alternative sigma factor L (σL) in the synthesis of OppA.
OppA, which is most likely a lipoprotein attached to the surface of the cell, is important for cold stress adaptation in L. monocytogenes and other bacteria (Borezee et al., 2000; Schliep et al., 2012), and participates in a number of bacterial processes: bacterial nutrient uptake, recycling of cell wall peptides, peptidoglycan turnover, and sensing the external medium via specific or nonspecific peptides (Goodell and Higgins, 1987; Monnet, 2003). Borezee et al. (2000) reported that an oppA mutant strain of L. monocytogenes was unable to use peptides longer than three residues, demonstrating that OppA mediates the transport of oligopeptides into the cell.
The extent of peptidoglycan turnover remained unchanged in mutants lacking oppA, indicating that the peptide fragments recycled by OppA are responsible for only a minor part of the recycling pathways in Escherichia coli and other gram-negative bacteria (Park, 1993; Reith and Mayer, 2011). However, several studies reported that some strains of L. monocytogenes showed higher levels of oppA transcripts in stationary phase cells grown at 4°C (Cacace et al., 2010; Chan et al., 2007). Recent results indicated that not all L. monocytogenes strains overexpress OppA when grown at low temperatures (Cabrita et al., 2013).
OppA also might be involved in biofilm formation. Studies in Vibrio fluvialis showed that in both rich and minimal media containing peptone or tryptone, an oppA mutant formed biofilm at higher levels than the wild type strain (Lee et al., 2004). Recent work reported that a putative ABC-transporter permease, which exhibits a 99% similarity to OppA, is involved in the negative regulation of L. monocytogenes biofilm formation (Zhu et al., 2008; 2011).
The opp system in L. monocytogenes might account for the contamination of cheese and other dairy fermented foods by this pathogen. In fact, these types of foods contain a wide variety of peptides ranging from 4 to at least 18 residues generated by the lactococcal cell envelope-associated proteinases (Borezee et al., 2000; Detmers et al., 1998). However, Cabrita et al. (2013) could not detect this protein in the supernatant of an 11°C-culture of a strain persistently present in a cheese dairy environment. The fact that this strain does not depend on this protein for the uptake of oligopeptides might have contributed to its persistence in the cold environment of the dairy industry.
Studies in E. coli showed that nonsense mutations or transcriptional reductions in OppA could provide bacterial resistance to aminoglycoside antibiotics such as kanamycin, streptomycin, isepamycin, and neomycin. The majority of these resistant strains displayed reduced or undetectable levels of OppA (Acosta et al., 2000; Kashiwagi et al., 1998). OppA binds to toxic peptide analogues and transports them to the interior of the cell (Monnet, 2003). Similar to other bacteria such as B. subtilis (Solomon et al., 2003), OppA mediates the transport of the toxic peptide derivate bialaphos in L. monocytogenes. Other antibiotics such as bacilysin, phosphinothricyl, and vancomycin enter the cells via a peptide transporter, which is significant for the development of antimicrobial drugs targeting ABC import systems (Eirich et al., 2011; Garmory and Titball, 2004).
Conclusions
The proteins secreted by L. monocytogenes play important roles in overcoming host cell defenses and adaptation to the saprophytic lifestyle. Exoproteome studies, promoted by high-resolution mass spectrometry methods, have gained importance, with several authors reporting that the stimuli required for the secreted protein response differed among strains. The exposure to stressful environmental conditions may cause an increasing differentiation of the response, and the extracellular proteins seem to be the major contributors to this differentiation. At the host temperature, Lmo0443, LLO and InlC can be good predictors (key exoproteins) of the virulence potential of the strains. At low temperatures, studies on FlaA and OppA using larger groups of L. monocytogenes strains, including sporadic and persistent strains collected from the food industry environment, may provide insight into the behavioral diversity within the species.
Footnotes
Acknowledgments
The authors thank Fundação Marquês de Pombal for financial support, and INIAV, IP for allowing Paula Cabrita's stay at Laboratório de Microbiologia (CBAA/DRAT), ISA.
Disclosure Statement
The authors declare that there are no conflicting financial interests.