
Abstract
Abstract
Rabies is one of the oldest diseases known to mankind. The pathogenic mechanisms by which rabies virus infection leads to development of neurological disease and death are still poorly understood. Analysis of rabies-infected proteomes may help identify novel biomarkers for antemortem diagnosis of the disease and target molecules for therapeutic intervention. This article offers a literature synthesis and critique of the differentially expressed proteins that have been previously reported from various in vitro/in vivo model systems and naturally infected clinical specimens. The emerging data collectively indicate that, in addition to the obvious alterations in proteins involved in synapse and neurotransmission, a majority of cytoskeletal proteins are relevant as well, providing evidence of neuronal degeneration. An interesting observation is that certain molecules, such as KPNA4, could be potential diagnostic markers for rabies. Importantly, proteomic studies with body fluids such as cerebrospinal fluid provide newer insights into antemortem diagnosis. In order to develop a complete integrative biology picture, it is essential to analyze the entire CNS (region-wise) and in particular, the brain. We suggest the use of laboratory animal models over cell culture systems using a combinatorial proteomics approach, as the former is a closer match to the actual host response. While most studies have focused on the terminal stages of the disease in mice, a time-series analysis could provide deeper insights for therapy. Postgenomics technologies such as proteomics warrant more extensive applications in rabies and similar diseases impacting public health around the world.
Introduction
R
Rabies is one of the oldest diseases known to mankind. Despite this, the pathogenic mechanisms by which rabies virus infection leads to development of neurological disease and death are not well understood (Dhingra et al.,2007; Wang et al., 2005). Electroencephalographic (EEG) abnormalities have also been recorded in animal models infected with different strains of RV. These alterations in neurophysiology could be attributed to the malfunction in neurotransmission. Functional genomics and proteomics have previously been used to determine host responses in cells/animals infected with different viruses (Dhingra et al., 2007). While there may be some understanding of the role of viral proteins in rabies pathogenesis, neither the contribution of host factors to rabies virus transcription/replication and axonal/trans-synaptic spread is understood nor are the mechanisms by which these factors determine the outcome of the disease precisely known (Dietzschold et al., 2008; Faber et al., 2004; Prosniak et al., 2001).
The current focus is therefore on identifying potential protein biomarkers in the host's CNS during the disease. A proteomics-based approach would not only provide us with the potential diagnostic and therapeutic targets but also give us a better insight into the pathological mechanisms and the associated host response. It would also help to identify differentially regulated proteins at the transcriptional, translational, as well as post-translational stages. It is a formidable task to identify and isolate a specific biochemical pathological course of the disease as the host would vary, depending on the virus strain (street/attenuated). In the current review, we have attempted to provide a comprehensive overview of the significant contributions in the field of rabies proteomics. The article particularly emphasizes the host proteins found to be differentially expressed when compared with uninfected controls.
Comparative Proteomics Analyses with Naturally Infected Specimens
Proteomic studies on rabies infection conducted to date are based on varied host systems such as mouse and human brain tissue samples, neuronal, and fibroblastic cell lines, which also contribute to the diversity in the type of responses observed. Table 1 provides a summary of the proteome analysis in RV infected different in vitro/in vivo host model systems and clinical samples.
B2C, attenuated strain derived from CVS-24; BD06, street virus isolated from dog; CVS, Challenge Virus Standard; 2-DE, two-dimensional gel electrophoresis; DIGE, Difference in Gel Electrophoresis; ESI, Electrospray Ionization; iTRAQ, isobaric tags for relative and absolute quantification; MALDI-TOF-MS/MS, Matrix Assisted Laser Desorption Ionization-Time of Flight-Mass Spectrometer; PV, Pasteur Virus; q-TOF, Quadrupole Time of Flight Analyzer; RV, Rabies Virus; SHBRV, Silver Haired Bat Rabies Virus; SRV9, Street Rabies Virus.
Proteome profiles of in vitro/in vivo systems vs. clinical specimens
Rabies virus propagation in animal models is preferred over tissue culture-based systems because unlike most viruses, rabies infection/replication in routine cell culture is not accompanied by any clear definable cytopathic effect, though the virus adapts easily on several cell lines such as neuroblastoma (N2a), Vero, and baby hamster kidney (BHK-21) cells (Meslin et al. 1996). Further, it is simpler to track the course of the disease in laboratory animals based on development of clinical signs. Proteomic analysis of the most common rabies infected in vitro models/in vivo host systems and street virus infected clinical samples revealed a list of differentially expressed proteins when compared to uninfected controls. Based on their functions, all the proteins (listed in Table 2 and Table 3) have been classified into eight broad categories as depicted in Figure 1.
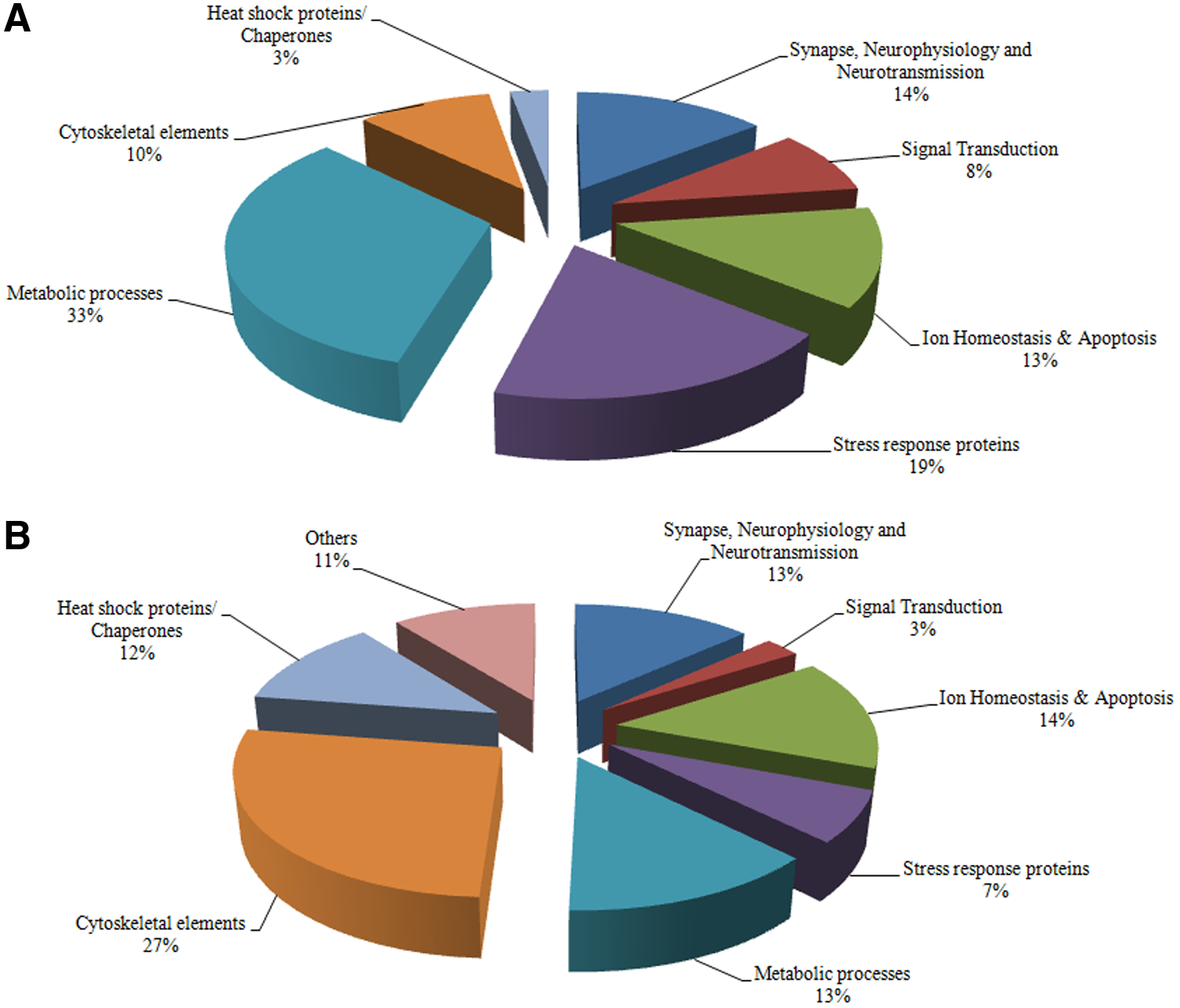
Percentage distribution of differentially expressed proteins identified in rabies infected in vitro/in vivo model systems
B2C, attenuated strain derived from CVS-24; BD06, street virus isolated from dog; CVS, Challenge Virus Standard; DRV, Dog Rabies Virus; i.c., intracerebral route of inoculation; i.m., intramuscular route of inoculation; ND, not done; NK, not known; PV, Pasteur Virus; SHBRV, Silver Haired Bat Rabies Virus; SRV9, Street Rabies Virus; ↑, upregulation; ↓, downregulation; P, present.
F, Furious Rabies, NS, not significant; P, Paralytic Rabies, PM, postmortem; PV, partially vaccinated; ↑, upregulation; ↓, downregulation.
It was observed that the expression of a vast majority of the metabolic proteins (33%) were altered in the cell culture and animal model systems (Dhingra et al., 2007; Kluge et al., 2013; Vaziri et al., 2012; Wang et al., 2011; Zandi et al., 2009; 2013) while analysis of rabid human and dog brain samples revealed changes in the expression of a large number of cytoskeletal associated proteins (27%) (Faber et al., 2004; Thanomsridetchai et al., 2011; Venugopal et al., 2013). This difference may be attributed to the diversity in virus cultivation, cell line, media component, and culture conditions. It has been demonstrated earlier that adaptation and propagation of viruses in different animal cells with different cultivation conditions involve changes in virus replication dynamics and protein expression. Changes in media components and differences in process conditions also have shown to alter expression of proteins, particularly those involved in metabolism and stress response (Kluge et al., 2013). This indicates that the hypotheses put forward based on laboratory model systems may sometimes deviate largely from the actual findings.
We also observed that, of all the differentially expressed proteins reported, only four proteins—CRMP2 (collapsin response mediator protein), anti-oxidative stress protein- PRDX1 (peroxiredoxins), signal transducing protein- α-enolase, and chaperone Hsp70—are common in the two groups [i.e., laboratory host model systems (in vitro/in vivo) and clinical specimens]. Further, the similarity in expression pattern is also noteworthy (Table 4). It is possible that this limited overlap in the protein profile may be due to the variability of different model systems or virus strains and limitations in the individual techniques employed.
↑, Upregulation;↓, Downregulation.
Techniques employed to study differential expression
Several gel-based proteomic studies have been carried out either using 2-DE (two- dimensional gel electrophoresis) or DIGE (difference in gel electrophoresis) to study RV host cell interactions. Different mass spectrometric (MS) based approaches following all gel/non-gel based platforms have made it possible to analyze such virus–host interactions successfully. A recent study has employed in comparison an enhanced technique of iTRAQ (isobaric tags for relative and absolute quantification) labeling to quantitatively analyze the proteome of human frontal cortex tissue in paralytic, furious, and partially vaccinated cases. It is obvious from Table 1 that, with high throughput techniques such as iTRAQ coupled with LC-MS/MS analysis, greater number of proteins can be identified. Venugopal and colleagues (2013) have identified 402 proteins, of which 94 were found to be differentially expressed with respect to controls. Nevertheless, 2-DE followed by q-TOF MS & MS/MS has helped identify 71 altered proteins in the hippocampus, brainstem, and spinal cord of street virus-infected CNS of dogs expressing paralytic and furious forms (Thanomsridetchai et al., 2011). Thus, tandem mass spectrometry increases the likelihood of finding proteins that may have not been considered previously. Pre-fractionation to 2-DE using reverse phase high performance liquid chromatography (RP-HPLC) can improve the chances of resolving and thus identifying low abundance proteins (Vaziri et al., 2006). It therefore appears that with each type of technique altogether different sets of proteins are being identified and hence an extremely narrow overlap is observed on comparing the proteins reported from laboratory model systems to those from clinical specimen. A combination of multiple proteomic methods may be utilized to develop an effective output. Complementary information may be obtained on using combinatorial strategies such as 2-DE/DIGE followed by iTRAQ and LC-MS/MS analysis to generate a better picture of the host protein profiles.
Proteomics insights: Neuronal dysfunction versus neuronal degeneration
It has been well established that the virus propagates at the synapse and modulates the synaptic physiology to disseminate across the trans-neuronal network. Alterations in the rabies-infected CNS are largely attributed to neuronal dysfunction. Proteomic experiments by Dhingra et al. (2007) have demonstrated that proteins involved in the formation of SNARE complexes are downregulated. SNARE-associated proteins such as Trim 9, SNAP α, annexin A7, synataxin 13, and syntaxin 18, which are also involved in docking of synaptic vesicle and exocytosis, are also downregulated. This may explain the decreased docking ability and accumulation of the synaptic vesicles at the presynaptic membranes in rabies-infected neurons. Impaired release and binding of serotonin as well as gamma-aminobutyric acid (GABA) in CVS infected primary neuronal cultures has also been demonstrated earlier. In fact, neurotransmitters such as norepinephrine (NE), dopamine (DA), dihydroxyphenylacetic acid (DOPAC), and serotonin were found to be stimulated in the hippocampus of rats infected with CVS in the early stages of infection, but the expression declined on development of clinical signs (Fu et al., 2005). This is further supported by the NMR findings of a rabid human brain that exhibited reduced levels of N-acetyl-aspartate (NAA) and GABA w.r.t. controls (Reinke et al., 2013) indicating that with progression of the disease, neurons are rendered incapable of releasing neurotransmitters at the synaptic junctions (Fu et al., 2005).
In addition to altered neurotransmission, in clinical specimens, a majority of the cytoskeletal proteins such as microtubule-associated proteins such as TUBA1C, β centractin, dynein light chain, dynamin, and tubulin α-2 were observed to be downregulated except GFAP, tubulin α-1 isoform 9, vinculin, and XIRP1. Defects in dynein/dynactin function due to altered expression are thought to affect the retrograde neuronal transport (Farahtaj et al., 2013), while suppression of actin associated proteins such as destrin, nebulin related anchoring protein, and myosin are further suggestive of abnormalities in the actin dynamics regulatory mechanism. The neurofilament heavy polypeptide was reported to be underexpressed in the brainstem of furious and paralytic rabies cases, whereas the medium polypeptide was downregulated in the spinal cord of paralytic rabies but upregulated in furious cases (Venugopal et al., 2013).
However, recent in vitro studies show an overall upregulation in the expression of a large number of cytoskeletal elements such as vimentin, tubulin subunits, and β-tropomyosin during RV infection. This overexpression may result from either an increased mRNA translation or an augmented cleavage of the cytoskeletal elements or both. Transcriptomic studies are required to validate the former, though several reports indicate the possibility of the latter also. RV-induced collapse in vimentin network and therefore cytoskeletal disruption (Zandi et al., 2013) and downregulation of total vimentin in CVS-infected neuroblastoma N2a cells (Wang et al., 2011) has also been demonstrated. While vimentin is thought to modify the intermediate filament network, CP–-a new form of heterodimeric capping protein (CP) involved in myofibrillogenesis actin organization has also been reported (Zandi et al., 2009). These findings are interesting as glycolysis in primary cells is also found to be cytoskeleton dependent (Kluge et al., 2013).
Thus, rabies, though often associated with absence of morphological changes supported by few or no neuronal degeneration (Jackson 2006), data from proteomic analysis indicate otherwise. Structural changes observed in the cerebral cortex of CVS infected mice such as swollen mitochondria, presence of vacuoles in the axons and pre-synaptic nerve endings, and beading of dendrites and axons, only corroborate the proteomic findings further (Scott et al., 2008). Even electron microscopic investigations reveal complete disappearance or reduction in a number of intracellular organelles such as rough endoplasmic reticulum, free ribosomes, and mitochondria in the pathogenic RV infected neurons (Li et al., 2005). Furthermore, demyelination and axonal injury in peripheral nerves, as well as axonal degeneration, have also been observed in paralytic rabies (Sheikh et al., 2005). With years of debate over neuronal dysfunction or degeneration responsible for pathogenesis in RV infection, it is clear that while neuronal dysfunction may be a major reason contributing to it, there is ample evidence supporting existence of the latter as well.
Potential Biomarkers
Rabies expresses itself in two distinct forms, paralytic or the dumb/numb form in which peripheral nerve demyelination or axonal degeneration have been observed, and the classical encephalitic/furious form characterized typically by hydrophobia wherein anterior horn dysfunction has been reported (Gadre et al., 2010). The paralytic form is often difficult to distinguish from and confused with the Guillain-Barre (GB) syndrome (Madhusudana et al., 2008). Discovery of protein biomarker appears to have the potential to answer the challenging question of clinical diagnosis of the disease antemortem. Mass spectrometry in its various guises has played an important role in discovery of biomarkers for many diseases. Venugopal et al. (2013) have reported several novel proteins found to be differentially expressed on analyses of rabid human proteomes, as listed in Table 5.
PLP1 was found to be downregulated (<two-fold) in both furious and paralytic rabies. It has also been implicated in axonal degeneration in patients suffering from chronic multiple sclerosis (MS) (Garbern et al., 2002). Another molecule, glutamate ammonia ligase (GLUL) is one of the important enzymes involved in ROS associated programmed necrosis (Moquin et al., 2010; Yu et al., 2011). Interestingly, while its overexpression (four-fold) has been reported in the later stages of Alzheimer disease (Venugopal et al., 2013), it is reported to be downregulated in the occipital lobe (Farahtaj et al., 2013), emphasizing the need to study the region-wise expression of such molecules in the entire CNS. GLUL is also reported to undergo oxidation in the later stages of the neurodegenerative disease Alzheimer's (Butterfield et al., 2006). CAMK2A, ubiquitously known to be involved in downstream signaling for building of neuromuscular junctions (Wu et al., 2010) and in synaptic plasticity in the hippocampus was found to be upregulated in paralytic (two-fold) and PV (four-fold) groups. CAMK2A aggregates are found to co-localize within the intracytoplasmic Negri bodies within the neurons. Immunohistochemical studies indicate higher expression in vaccinated cases of paralytic rabies in comparison to both nonvaccinated paralytic and furious cases (Venugopal et al., 2013). CAMK2A is also elevated in individuals with bipolar disorders (Beraki et al., 2005).
Other novel upregulated proteins such as OPA 1 were reported (though <two-fold) in paralytic, furious, and partially vaccinated cases, and HPCAL4 only in paralytic rabies (2.4-fold). Interestingly, PLP1, GLUL, CAMK2A, and OPA1 have all been implicated in schizophrenia too (Martins-de-Souza et al., 2010; Qin et al., 2005; Venugopal et al., 2013). AP3B2 gene expression is also downregulated by the human T-lymphotropic virus type-1 (HTLV) (Nair, 2004). Interestingly, while PDCD6IP is downregulated (<three-fold) in paralytic or furious cases, it is found to decrease five-fold in partially vaccinated cases. While its depletion is known to cause accumulation of unusual actin molecules (Akgul et al., 2009), its increased expression has been associated with survival of injured neurons (Rojo et al., 2011). The roles of TAGLN3, PDCD6IP, and LASMP in neuroviral/neurodegenerative diseases are still not explored and further investigation is needed to determine if either/all hold any potential as therapeutic or diagnostic marker.
Another such molecule that may be investigated for its role as a diagnostic marker to identify paralytic rabies is KPNA4. It was found to increase seven-fold in partially vaccinated cases and three-fold in paralytic cases. Karyopherin alpha 4 has been previously reported to be downregulated in tuberculosis meningitis and hepatocellular carcinoma (Kumar et al. 2012; Yu et al. 2007).
Neurons being the primary host for the rabies virus makes it imperative to study differential protein expression in the brain tissue, however from the diagnosis point of view, rabies-infected CSF proteome may provide a better insight. Metabolomic studies reveal that rabies-infected brain and CSF exhibit vast differences in their metabolite profiles. Upregulation was observed for CSF metabolites such as lactate, quinolinate, glutamate, malate, ornithine, and many other lipid metabolites with progress in disease, while others such as acetate, myo-inositol, pyroglutamate, and succinate were nearly absent. Increased lactate levels were also reported in the frontal cortex and white matter, suggesting anaerobic metabolism in rabies. In addition, the CSF metabolite profiles were found to differ in rabies survivors on comparison with those who died of rabies (O'Sullivan et al., 2013; Reinke et al., 2013). The field of proteomics and transcriptomics has not yet been explored for rabies-infected CSF but hold greater prospect in determining potential antemortem diagnostic markers for rabies.
Conclusions and the Way Forward
Based on the type of protein profiles generated on analysis of infection with different virulent strains of RV, including the wt-street strains, we draw the following conclusions:
i. A vast majority of the proteins that exhibited altered expression in the in vitro/in vivo laboratory model systems belonged to the metabolic processes category (33%). This may be attributed to the differences in virus culture conditions including nature of the cell line, media component, etc. ii. Changes in the expression of a large number of cytoskeletal associated proteins (27%) have been observed in the clinical specimen of rabid human and dog brain samples, indicating that not only neuronal dysfunction but also neuronal degeneration contributes to the disruption of neurotransmission dynamics in RV infection. iii. The vast difference in the host protein profiles with only four common proteins—CRMP2, PRDX1, α-enolase, and Hsp70—between the two groups, in vitro/in vivo systems and clinical specimens may be due to the diversity of the host and the techniques employed to study them and their limitations. This may be resolved by adopting combinatorial strategies that can generate complementary information to have a better correlation while studying rabies proteomes in model host systems and actual clinical specimens. iv. The currently available information on rabies proteome from human samples is restricted to only the occipital lobe and frontal cortex. In order to generate comprehensive data, it is imperative to perform proteomic profiling of all the regions of brain tissue. A targeted region-wise analysis of certain proteins such as GLUL that exhibit different expression patterns in different areas of the brain, an amalgamation of 2-DE, iTRAQ, and LC-MS/MS may provide deeper insight into its patho-physiological role in rabies. v. Proteins such as KPNA4 may prove to be useful diagnostic biomarker that may help distinguish paralytic rabies from GB, but further studies are warranted. vi. Proteomic profiling of rabies-infected CSF specimen may provide a better insight into protein biomarkers for antemortem diagnosis.
We therefore conclude that currently only partial data from proteomic analysis of rabies-infected CNS or other varied host cell systems are available. In order to generate a holistic picture, it is essential to analyze the entire CNS (region-wise) and in particular, the brain. We further suggest the use of laboratory animal models over cell culture systems, using a combinatorial proteomics approach, as they are a closer match to the host response observed in reality. While most studies have focused on the terminal stages of the disease in mice, a time-series analysis may provide a better insight for therapy. In addition, the proteomics data may be complemented with more information from metabolomic and transcriptomic studies of the same samples. Using CSF as a clinical specimen, especially in antemortem laboratory investigation, is crucial, as potentially identifiable lead biomolecules may be used either as diagnostic markers or more importantly, for therapeutic intervention.
Footnotes
Author Disclosure Statement
The authors declare that no competing financial interests exist.