
Abstract
Abstract
Exercise and physical activity have long been recognized for health promotion and to delay the onset of many pathological situations such as diabetes and cancers. Still, there appears to be an upper limit on the beneficial health effects regarding intensity and frequency of exercise training. In humans, the effect of exercise training in the male reproductive system has been studied mainly through the analysis of semen quality parameters, with inconsistent results. Less is known on molecular biomarkers of exercise-related changes in testis at the protein/proteome level. This review offers an in-depth analysis on the small scale protein studies available primarily from the preclinical studies and interprets their functional impact on the reproductive health with a view to humans. In all, exercise training in preclinical models seems to negatively modulate, in the course of health, critical functions that directly affect spermatogenesis, such as testosterone biosynthesis, energy supply, and antioxidant system components. Exercise training induces apoptosis, leading to the impairment of spermatogenesis and, consequently, to male infertility. In pathological conditions, an improvement in the testicular functions is observed by increases in steroidogenic enzymes and antioxidant defenses, and reductions in activity of inflammatory pathways. Importantly, the mechanisms by which exercise training modulates the reproductive function are far from being fully understood. The analyses of the testis proteome in varying exercise conditions would inform the molecular mechanisms involved and identify putative theranostics opportunities. Such future research is a cornerstone for health promotion in the pursuit of reproductive health informed by omics systems sciences.
Introduction
M
Exercise training alters several physiological processes, including energy metabolism (Barrero et al., 2015), bone density (Barry et al., 2008; Maimoun et al., 2005; Nichols et al., 2003), muscle metabolism (Denvir et al., 1999; Peake et al., 2005; Suzuki et al., 2006), liver metabolism (Chevion et al., 2003; Pettersson et al., 2008), and myocardial remodeling (Laslett et al., 1996; Neilan et al., 2006), and also modifies electrocardiogram profiles (Brosnan et al., 2014; Tulloh et al., 2006).
In what concerns the male reproductive system, for instance, regular moderate exercise training prevents overweight and underweight (common factors of subfertility) and regulates hormone levels (Barazani et al., 2014). In fact, men who perform regular moderate exercise training scored higher in almost all sperm parameters (Sharma et al., 2013). However, it should be noticed that the physiological effects are largely dependent on the intensity and frequency of the exercise training (O'Keefe et al., 2012). Endurance exercise frequently leads to semen deterioration, with men presenting decreased spermatozoa concentration, altered motility, and abnormal morphology (Safarinejad et al., 2009; Wise et al., 2011). Moreover, endurance exercise disturbs the hypothalamic–pituitary–gonadal (HPG) axis (Lucia et al., 1996), causing a decrease in the circulating testosterone levels (Hackney et al., 2003).
The effect of exercise training in the overall male reproductive system has been the focus of several studies (Cooper et al., 1998; Fry et al., 1998; Gorostiaga et al., 2004; Hall et al., 1999; Vaamonde et al., 2012). Omics sciences, which include large-scale data-rich biology fields (e.g., genomics, transcriptomics, proteomics, and lipidomics) have been particularly useful to identify and characterize, by high-throughput technologies, biological molecules, and build cell-, tissue- and organism-specific molecular profiles (Longuespee et al., 2014; Rabara et al., 2014).
In spite of the recent advances, the knowledge on exercise-related changes in testis, particularly at the protein/proteome level, is still scarce. Drawing from the field of proteomics and integrative biology, this review offers an in-depth analysis on the small-scale protein studies currently available using rodent models since human studies on testis are not available. The panel of proteins reported in such studies is here discussed in what concerns protein expression, activity, and functional impact on male reproductive health. This study may constitute the basis for large-scale omics studies, so that novel biomarkers that reflect the impact of exercise training on the reproductive health could be discovered.
Data Retrieval and Analysis
A comprehensive Pubmed search was conducted to select studies that reported the effect of exercise training on testis physiology (search terms: exercise and testis). All results were analyzed and only the studies that assessed the effect of exercise training, specifically on testis protein profile, were considered for further analysis. The search revealed 19 studies that applied nonproteomic techniques to study the impact of different exercise training protocols (swimming, free wheel running, and treadmill training) in testis. All studies were performed in rodents (Mus musculus and Rattus norvegicus) and used testis extracts for analysis. For this reason, we were not able to distinguish cell type-specific alterations.
From the 19 studies, 16 were performed in healthy rodents (two groups practiced acute exercise and 14 chronic exercise). The remaining studies used nonhealthy rodents and for those only one study per condition was analyzed. All the proteins reported to suffer alterations either in terms of expression or activity in consequence of the exercise training protocols performed are listed in Table 1. It is noteworthy to mention that, as far as we know, there is no study on the effect of exercise training on human testis proteome.
Although these proteins have different isoforms, in the original paper, total protein expression/activity was measured. Therefore, the Uniprot ID presented is only representative and corresponds to isoform 1.
Impact of Exercise Training on Testis Molecular Pathways
According to the literature reviewed, exercise affects several testis functions, from spermatogenesis to hormone production. In this section, the molecular pathways and cellular alterations due to exercise (chronic and acute) will be discussed.
Exercise training and spermatogenesis
There is evidence that exercise training produces divergent effects on spermatogenesis. On one hand, long-term endurance exercise training is associated with decreased production of testosterone, and consequently spermatogenesis is impaired. On the other hand, moderate volunteer exercise training, attenuates the age-associated impairment in spermatogenesis (Torma et al., 2014).
Some studies reported negative effects of exercise training on spermatogenesis (Jana et al., 2014; Joseph et al., 2014). After chronic exercise, a decrease in the activity of sorbitol dehydrogenase (SDH) enzyme occurs. Since SDH converts sorbitol into fructose (Jana et al., 2008) and spermatogonia and spermatozoa use glucose and fructose as main energy sources (White et al., 2013), it is not surprising that the lack of SDH can affect spermatogenesis.
Another protein affected by exercise training is the outer dense fiber 1 (ODF1). ODF1 is involved in spermatozoa morphogenesis, more specifically in the process of head and tail junction (Yang et al., 2012). With exercise, ODF1 expression decreases, leading to incomplete morphogenesis (tails separated from heads) and, consequently, to impaired spermatogenesis (Torma et al., 2014; Yang et al., 2012).
Besides this apparently harmful effect on spermatogenesis, exercise training seems to benefit DNA damage regulation. Histone H2AX phosphorylation on serine 139 is a well-established marker of DNA damage: it increases after DNA damage to induce DNA repair systems (Kuo et al., 2008). A number of studies reported a decrease in H2AX phosphorylation upon free wheel running and treadmill training (Chigurupati et al., 2008; Joseph et al., 2014). Since H2AX phosphorylation is a biomarker for DNA double-strand breaks, the decreased phosphorylation levels observed upon exercise training suggests that it is beneficial to DNA preservation (Sharma et al., 2012).
Exercise training and hormones
Previous studies demonstrate that exercise training has a strong influence in testosterone biosynthesis. Acute and intense exercise training leads to an increase in testosterone levels, while chronic and intense exercise training decreases testosterone levels due to subclinical inhibition of normal reproductive function (Constantini et al., 2013). The effect of exercise training on testosterone levels is mainly a result of the HPG axis deregulation. Upon exercise training, gonadotropin-releasing hormone (GnRH) secretion is compromised due to either resistance to luteinizing hormone (LH) or depletion of LH secretion from the pituitary. The decrease of the GnRH levels leads to a decrease in the testosterone biosynthesis that occurs in the Leydig cells in the testis (Di Luigi et al., 2012; Hackney, 2008; Safarinejad et al., 2009). Interestingly, this negative impact of exercise training on testosterone levels has also been described in humans (Hackney, 1996; Di Luigi et al., 2012).
Additionally, the studies hitherto published point through a decreased activity of the proteins involved in the testosterone biosynthesis. After chronic exercise training, 3β-hydroxysteroid dehydrogenase (3β-HSD), 17β-hydroxysteroid dehydrogenase (17β-HSD), and testicular steroidogenic acute regulatory (StAR) expression (Jana et al., 2008; Manna et al., 2003; 2004b; Samanta et al., 2006) and activity levels are reduced. By contrast, the expression of steroid 17-alpha-monooxygenase (CYP17A1) is higher in exercise trained-mice when compared with the sedentary group (Barone et al., 2013). This protein is responsible for catalyzing the transformation of pregnenolone and progesterone into dehydroepiandrosterone and 4-androstenedione, respectively, both crucial steps in the pathway of active steroids formation (Luu-The, 2013).
These contradictory results further confirm the ambiguous effect of exercise training on testis physiology. Figure 1 illustrates the effect of exercise training on spermatogenesis and testosterone biosynthesis. The data analyzed suggest that exercise training influences both the HPG axis and the Leydig cells themselves, leading to a testicular dysfunction characterized by a decrease on testosterone production. Since testosterone controls spermatogenesis, this process is also affected, resulting in spermatozoa defects and impaired male fertility (Kobayashi et al., 2002). Considering the literature, we strongly believe that discrepancies among the exercise training protocols used in the studies are the main contributing factor to the controversial results.
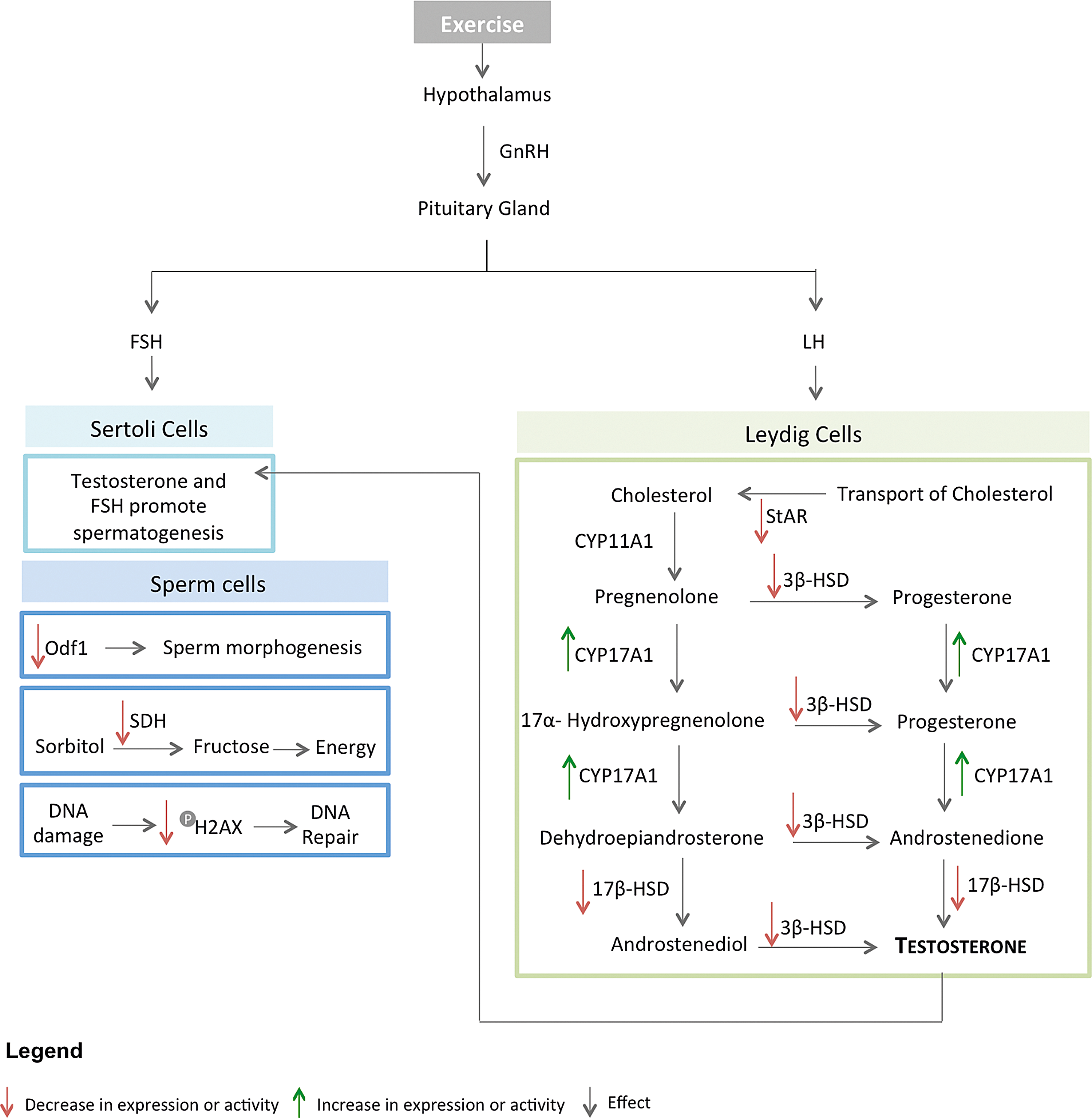
Schematic representation of the relation between exercise training, testosterone biosynthesis, and spermatogenesis in testis. Exercise reduces testosterone levels through a decrease on StAR, 3β-HSD, and 17β-HSD levels or activity. By contrast, CYP17A1 levels increase. Diminished levels of testosterone lead to spermatogenesis abnormalities. Energy metabolism, DNA integrity, and spermatozoa morphogenesis are affected due to a decrease on the activity/levels of SDH, phosphorylated H2AX and Odf1, respectively. Red arrows represent decreased expression or activity; Green arrows represent increased expression or activity. 3β-HSD, 3 beta-hydroxysteroid dehydrogenase; 17β-HSD, estradiol 17-beta-dehydrogenase 1; CYP11A1, cholesterol side-chain cleavage enzyme, mitochondrial; CYP17A1, steroid 17-alpha-hydroxylase; FSH, follicle-stimulating hormone; GnRH, gonodotropin-releasing hormone; H2AX, histone; LH, luteinizing hormone; Odf1, outer dense fiber protein 1; SDH, sorbitol dehydrogenase; StAR, steroidogenic acute regulatory protein, mitochondrial.
Exercise training and reactive oxygen species
One of the consequences of increasing oxygen consumption during exercise training is the generalized increase in reactive oxygen species (ROS) production (Lanfranco et al., 2013; Manna et al., 2004a). In healthy subjects, who regularly practice moderate exercise training, testis antioxidant capacity increases and is able to antagonize the rise in ROS production. However, when exercise intensity increases, the organism is no longer capable of eliminating ROS efficiently, thereby leading to increased ROS concentration and a decreased antioxidant activity (du Plessis et al., 2011; Olive, 2010).
Several enzymes in the antioxidant system are profoundly affected by exercise training. Total activity of superoxide dismutase (SOD) and glutathione peroxidase (GPx) decreased in animals submitted to chronic exercise training (Aksoy et al., 2006; Manna et al., 2003; 2004a; Jana et al., 2008; Samanta et al., 2006) Curiously, Chigurupati and colleagues (2008) described both an increase and a decrease in GPx1 and GPx4, respectively.
Other antioxidant components commonly altered are the glutathione-S-transferase (GST) and the glutathione reductase (GR). The activity of both GST and GR was decreased in animals that practiced chronic swimming (Chigrinskiy et al., 2011; Manna et al., 2003; 2004a; 2004b; Jana et al., 2008; 2014; Samanta et al., 2006). Moreover, glutathione (GSH) levels were shown to be reduced after chronic exercise (Chigrinskiy and Conway, 2011; Jana et al., 2014; Manna et al., 2004b; Samanta et al., 2006) and controversial results were obtained in what concerns catalase (CAT) activity (Aksoy et al., 2006; Chigrinskiy et al., 2011; Manna et al., 2003; 2004a; 2004b; Jana et al., 2008; Samanta et al., 2006).
In general, chronic exercise training, either swimming or running, decrease the antioxidant capacity in testis. However, GPx and CAT are still a major discussion. Curiously, both proteins catalyze the conversion of H2O2 into H2O, following SOD activity that converts O2.- into H2O2. Since SOD expression and activity decrease with exercise, a reduction in H2O2 may occur, probably leading to unchanged or reduced GPx and CAT activities.
Exercise training and apoptosis
Currently, there is evidence that exercise training modulates several factors, which may alter apoptosis in a variety of tissues. ROS produced during exercise training interact with intracellular and extracellular proteins and trigger apoptosis (Phaneuf et al., 2001), which in turn is a vital regulatory process of male germ cell development (Aitken et al., 2013). Jana et al. (2014) observed an increase in caspase-3 activity on testis after exercise training. However, in other studies, the activity of caspases-3, −8, and −9 remained unchanged (Faraone-Mennella et al., 2010; Ferreira, 2012; Shokri et al., 2010). Furthermore, it was shown that the pro-apoptotic protein BAX and the anti-apoptotic protein BCL-2 are decreased and increased, respectively, while apoptosis inducing factor (AIF), which is involved in the initiation of caspase-independent pathway, is increased (Joseph et al., 2014).
Alterations on Poly ADP ribose polymerases (PARP) were also observed. PARP-1 is an enzyme that becomes activated upon cleavage, leading to the formation of an 89 kDa fragment, during the apoptotic process. Poly ADP ribose polymers, the major products of PARP-1 activation, and PARP-1 89 kDa apoptotic fragment are increased in trained rats (D'Amours et al., 2001; Faraone-Mennella et al., 2010; Ko et al., 2012).
Exercise training also modulates the expression of heat shock proteins (HSPs), more precisely by decreasing the activity of HSP32 and HSP70 (Chigurupati et al., 2008; Koeva, 2005). These proteins can be activated by a variety of endogenous and exogenous factors (Middendorff et al., 2000; Piotrkowski et al., 2009). HSPs enhance cell survival in response to stress, such as oxidative stress, since they are capable of attenuating the toxicity of denatured and misfolded proteins that are accumulated during stress (Murphy, 2013). Sirtuin 1 (SIRT1) is a NAD-dependent deacetylase that inhibits p53, thereby preventing apoptotic events. However, when ROS increase, SIRT1 is inhibited and, consequently, p53 is activated (Hori et al., 2013).
Figure 2 represents the interplay between the apoptotic pathway and ROS production in testis after exercise training. We suggest that exercise training affects the balance between ROS production and antioxidant systems, increasing the presence of ROS. In turn, ROS modulate apoptosis in testis. Moreover, exercise training decreases the activity of antioxidant system components, which also contributes to alter the apoptotic events. Curiously, the extrinsic pathway seems to be more affected than the intrinsic pathway. After exercise, BCL-2, BAX, and caspase-9, three key players of the intrinsic pathway, do not suffer pro-apoptotic alterations, neither on their expression nor in the activity. However, caspase-3 was shown to be increased or unaffected. Caspase-3 is the main effector caspase, which reinforces that ROS influence apoptosis in testis after exercise training (Circu et al., 2010; Hori et al., 2013; Powers et al., 2008; Simon et al., 2000).

Schematic representation of the relation between exercise training, antioxidant systems and apoptosis in testis. Increased levels of O2−. induce the antioxidant system components (SOD; GPx and CAT). However, exercise has contrasting effects on the levels/activity of antioxidant system components, suggesting that a limit for the antioxidant system is reached. Moreover, GR and GST, enzymes indirectly linked to the ROS elimination, are decreased further strengthen that the antioxidant system was bypassed. High ROS levels activate Casp8 due to NF-kB inhibition. In turn, Casp8 activates Casp3 and cleaves Bid (tBid). tBid activates mitochondrial apoptotic signaling pathway, leading to the activation of apoptogenic factors such as Bax and Casp9 (through the apoptosome) and AIF. In physiological conditions, AIF is inhibited by HSP. Casp9, together with Casp8 activates Casp3, which in turn cleaves PARP1 and promotes apoptosis. Additionally, ROS inhibit Bcl-2, an anti-apoptotic protein and Sirt1, a protein that keeps p53 inhibited. After exercise, it appears that there is an increase in Casp3 and 89kDa PARP1, and a decrease in Sirt1, suggesting a pro-apoptotic stimulus. By contrast, decreased Bax, increased Bcl-2 and unchanged Casp8 and Casp9 suggest an anti-apoptotic stimulus. Red arrows represent decreased expression or activity; Green arrows represent increased expression or activity; Orange equal sign represent unchanged expression or activity. AIF, apoptosis inducing factor; Bax, Bcl-2-associated X protein; Bcl-2, B-cell lymphoma 2; Bid, BH3 interacting-domain death agonist; Casp3, caspase 3; Casp8, caspase 8; Casp9, caspase 9; CAT, catalase; GPx, glutathione peroxidades; GR, glutathione reductase; GST; glutathione S-transferase; HSP; heat shock protein; NF-kB, nuclear factor kappa beta; p53, cellular tumor antigen p53; PARP1, poly [ADP-ribose] polymerase 1; ROS, reactive oxygen species; Sirt1, NAD-dependent protein deacetylase sirtuin-1; SOD, superoxide dismutase.
Functional Benefits of Exercise Training on Aging
Aging is associated with an impairment on the testicular function, which has been attributed to oxidative stress and chronic inflammation with the subsequent damage of DNA, proteins, and lipids (Veldhuis et al., 2009; Woods et al., 2012). Lipid damage leads to the loss of membrane permeability and fluidity, thus impairing the molecular transport and the intracellular signaling. Due to the production of ROS, aberrant proteins are also accumulated within aged cells, but the main features observed are alterations in the mitochondria function (Amaral et al., 2013a).
Oxidative stress and chronic inflammation have been associated with the increased testicular failure observed in aged mice. The aging process is related to a decrease in serum testosterone concentration, which is associated with a reduction in the LH receptors number and the StAR protein levels, and a downregulation of the downstream steroidogenic enzymes involved in testosterone synthesis (Chen et al., 2005). Several studies proved that life-long moderate exercise training reduces the age-related physiological processes and improves the antioxidant activity and response to stress (Chigurupati et al., 2008; Zhao et al., 2013).
In testis protein profile of SAMP8 (senescence-accelerated prone mouse) mice, a mouse model for aging studies that presents age-associated pathologies and increased rate of senescence (Takeda, 1999), exercise training increased the levels of serum and testicular testosterone, together with an increase in CYP11A1 enzyme and StAR levels. To assess the antioxidant defense system in mice after chronic swimming, testicular physiology and mRNA levels of antioxidant enzymes SOD, CAT, and GPx were evaluated, demonstrating a significant increase in the expression and total activity of these enzymes (Zhao et al., 2013).
Nuclear factor-erythroid 2 related factor (NRF2), a transcription factor that activates genes in response to oxidative stress (Shih et al., 2007), was significantly increased (Zhao et al., 2013). Moreover, inflammation is one of the conditions that emerges with age and leads to an increase in pro-inflammatory molecules, such as interleukins and tumor necrosis factor alpha (TNF-α), which activates nuclear factor kappa B (NF-κB) (Lawrence, 2009; Woods et al., 2012). In aged animals submitted to swimming, TNF-α is decreased and interleukin 1 beta (IL-1β) and NF-κB also suffers a reduction. The levels of cyclooxygenase 2 (COX2), involved in the induction of inflammation (Ricciotti et al., 2011), also decrease in response to chronic exercise training. In contrast, the levels of interleukin IL-10 and TGF-α, which are anti-inflammatory molecules, are increased (Zhao et al., 2013).
Along with aging, mitochondrial proteins and genome (mtDNA) suffer alterations and mutations. The electron transport chain (ETC) is one of the main sources of ROS production. Since ETC is located near the mtDNA, the ROS produced damage the mtDNA altering the mitochondrial biogenesis (Amaral et al., 2013b). The mtDNA mouse model results from the expression of a proofreading-deficient mitochondrial DNA polymerase. Consequently, there is accumulation of mtDNA mutations that are associated with increased markers of oxidative stress and apoptosis (Kujoth et al., 2005). Safdar et al. (2011) tried to understand if exercise training could reverse the effects of mtDNA mutations on testis. They showed a decrease in the levels of mitochondrial ETC subunits, as well as, in the activity of COX in sedentary rats, which was prevented in rats submitted to chronic exercise training.
Functional Benefits of Exercise Training on Several Pathological Conditions
The beneficial effect of exercise training in individuals with different diseases, such as type 2 diabetes, cardiac arrhythmias, and cancer, is well-established (Hagberg et al., 1989; Kavanagh et al., 1988;) The effect of exercise training on testis of individuals with such diseases was indirectly assessed using the low capacity runners (LCR) rat models (Torma et al., 2014). These rats present an inefficient oxygen metabolism, which is a common feature in a variety of diseases (inflammation, cardiac and neurodegenerative dysfunction, among others) (Koch et al., 2008).
Since there is only one study regarding the testis protein profile of LCR rats, the results obtained are preliminary and should be carefully considered. In the referred study, an increase in ODF1 protein and lactate dehydrogenase C (LDHC) was observed. These two proteins are crucial to sperm tail function: ODF1 is involved in motility and LDHC in ATP production (Odet et al., 2008; Torma et al., 2014). However, exercise training seems to increase the acetylation of p53 in LCR rats. Curiously, proteins responsible for p53 acetylation, such as BAX and BCL-2, remained unaffected (Tang et al., 2008; Torma et al., 2014).
Are Omics Biomarkers the Future?
Omics science allows profiling of cells, tissues, and even full systems at several organizational levels, from genes to proteins, lipids, and protein–protein interactions. Further, it is performed in a dynamic way, in different time frames and, for instance, comparing different lifestyle conditions: exercise versus sedentary groups. The amount of data generated is enormous but, more important, is reliable due to the use of sophisticated high-throughput advanced Omics technologies (e.g., MudPIT and protein microarray-based technology) (Chandramouli et al., 2009). Together with bioinformatics tools, it makes evaluation possible of the impact of a diversity of external stimulus on a system biology level.
Since exercise-induced alterations in testis proteome are mostly unknown, we reviewed the publications concerning specific protein modifications in response to distinct types of exercise to serve as the basis for further proteomic studies within this field. In our opinion, high-throughput proteomics studies are of utmost importance to enhance the current knowledge on the effect of exercise training on testicular physiology and to move this topic forward. Hence, the authors have combined all the existing data to give their own integrated omics perspective, addressing the relevance of particular molecules on crucial testis pathways and functions. The future is to perform integrated omics studies in testis of different mice models after different exercise protocols in order to identify specifically-modulated proteins that may aid monitoring of exercise impact.
Conclusions
This review provides a new perception into how exercise training influences testis physiopathology. Despite the limited number of studies, exercise seems to negatively modulate critical functions that directly affect the production of sperm cells, such as testosterone biosynthesis, energy supply, and antioxidant pathways. Furthermore, exercise induces apoptosis, which leads to the impairment of spermatogenesis and, consequently, to male infertility. However, it should be noticed that the conclusions obtained are highly influenced by the exercise training protocols.
Several studies showed that exercise training jeopardizes male infertility, by affecting spermatozoa concentration, motility, and morphology (Arce et al., 1993; du Plessis et al., 2011; 2014). This phenotype is frequently observed in athletes. However, other studies demonstrated that exercise training is beneficial for male fertility, particularly when an underlying pathological condition is present.
In age-related pathological conditions, exercise training seems to improve testicular functions by increasing steroidogenic enzymes and antioxidant defenses and by decreasing molecules involved in inflammatory pathways, such as NF-κB, TNF-α, and interleukin IL-1β. The restoration of mitochondrial function after exercise in rats with mtDNA mutations was also verified in previous studies, indicating that exercise can ameliorate the biological mitochondrial alterations that occur with aging. Despite strong evidence for the health benefits of exercise training on pathological situations, such as hypertension and diabetes, the impact of exercise in male reproductive function remains controversial and, therefore, more in-depth omics scale studies are urgently needed. Future research on testis proteomes would also serve as a cornerstone of health promotion informed by omics and systems sciences.
Footnotes
Acknowledgments
The study was supported by “FCT–Fundação para a Ciência e Tecnologia (PTDC/DTP-PIC/0460/2012) and co-financed by FEDER through “Eixo I do Programa Operacional Fatores de Competitividade (POFC) do QREN” (COMPETE: FCOMP-01-0124-FEDER-028692).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.