
Abstract
Abstract
Sport and Exercise Medicine is one of the important subspecialties of 21st century healthcare contributing to improving the physical function, health, and vitality of populations while reducing the prevalence of lifestyle-related diseases. Moreover, sport and exercise are associated with injuries such as Achilles tendinopathy, which is a common tendon injury. The angiogenesis-associated signaling pathway plays a key role in extracellular matrix remodeling, with increased levels of angiogenic cytokines reported after cyclic stretching of tendon fibroblasts. We investigated the variants in angiogenesis genes in relation to the risk of Achilles tendinopathy in two population samples drawn independently from South Africa (SA) and the United Kingdom (UK). The study sample comprised 120 SA and 130 UK healthy controls, and 108 SA and 87 UK participants with Achilles tendinopathy. All participants were genotyped for five functional polymorphisms in the vascular endothelial growth factor, A isoform (VEGFA) (rs699947, rs1570360, rs2010963) and kinase insert-domain receptor (KDR) genes (rs1870377, rs2071559). The VEGFA A-G-G inferred haplotype was associated with an increased risk of Achilles tendinopathy in the SA group (15% in controls vs. 20% in cases, p = 0.048) and the combined SA+UK group (14% in controls vs. 20% in cases, p = 0.009). These new findings implicate the VEGFA gene with Achilles tendinopathy risk, while highlighting the potential biological significance of the angiogenesis signaling pathway in the etiology of Achilles tendinopathy. The evidence suggesting a genetic contribution to the susceptibility of sustaining a tendon injury is growing. We anticipate that high-throughput and multi-omics approaches, building on genomics, proteomics, and metabolomics, may soon uncover the pathophysiology of many diseases in the field of Sports and Exercise Medicine, as a new frontier of global precision medicine.
Introduction
S
The underlying biological pathways and molecular mechanisms culminating in acute and overuse injuries are currently under investigation in an effort to improve both preventive and treatment modalities. To date, several intrinsic factors, including genetics, have been identified as risk factors predisposing an individual to musculoskeletal soft tissue injuries (Collins and Raleigh, 2009; Meeuwisse, 1994; Posthumus et al., 2011). The results of genetic association studies, along with histopathological examination and expression studies, have assisted in elucidating some of the key biological pathways that may potentially be involved in the etiology of musculoskeletal soft tissue injuries (Collins et al., 2015; Riley, 2008; Xu and Murrell, 2008).
Tendons are metabolically active tissues and are able to respond to stimuli, but the rate of metabolism is slow (O'Brien, 1997). It is possible that the metabolic rate may increase in response to mechanical loading to allow for remodeling of the extracellular matrix (ECM) (Wang, 2006). Evidence shows that mechanical stimuli cause an increase in the expression of several cell signaling molecules (Jiang et al., 2012; Legerlotz et al., 2012; Skutek et al., 2001). In particular, a number of studies have reported a pro-angiogenic expression profile after cyclic strain of tenocytes (Mousavizadeh et al., 2014; Nakama et al., 2006; Petersen et al., 2004) and during remodeling (Petersen, Unterhauser, Pufe, et al., 2003). Moreover, an increase in vascularity was observed after mechanical loading and in symptomatic Achilles tendons (Lewis et al., 2009; Molloy et al., 2003; Pufe et al., 2001; Scott et al., 2008); whereas normal healthy tendons are relatively avascular structures (Pufe et al., 2001). A number of cell signaling molecules function in the angiogenesis pathway, such as transforming growth factor-β and fibroblast growth factor, with vascular endothelial growth factor (VEGF) considered a critical regulator of angiogenesis.
The VEGFA gene codes for the A isoform of the protein. This potent angiogenic factor is able to function in a diverse range of signaling pathways (Evans, 2015), and expression levels may be induced by mechanical loading, hypoxia, and a number of biochemical factors/molecules (Neufeld et al., 1999; Petersen, Pufe, et al., 2003). Although healthy tendons display negligible amounts of VEGF, this increases significantly during remodeling (Mousavizadeh et al., 2014; Petersen et al., 2004; Pufe et al., 2001). Furthermore, expression levels of VEGF in degenerative tendons are comparable to fetal tendons (Legerlotz et al., 2012; Pufe et al., 2001). VEGF binds to one of two high-affinity receptors, Flt-1 (fms-related tyrosine kinase 1) or KDR (kinase insert-domain receptor), which are localized on the cell surfaces of vascular endothelial cells. KDR, encoded by the KDR gene, is considered the major mediator of the angiogenic, mitogenic, and permeability-enhancing effects of VEGF (Bao et al., 2009; Ferrara et al., 2003) in both normal physiology and pathological processes (Ferrara et al., 2003).
Recently, genetic variants within the VEGFA and KDR genes were associated with risk of anterior cruciate ligament (ACL) ruptures (Rahim et al., 2014). Specifically, independent associations as well as haplotype combinations were implicated, thereby suggesting genomic intervals harboring potential DNA motifs within these two genes (Rahim et al., 2014). The aim of this study was to investigate the previously associated functional VEGFA rs699947, rs1570360, rs2010963 and KDR rs1870377 and rs2071559 variants with the risk of Achilles tendinopathy in a South African and a British population.
Materials and Methods
Participant recruitment
A total of 120 physically active South African Caucasian asymptomatic control (SA CON group) participants and 87 participants with chronic Achilles tendinopathy (SA TEN) were included in this study. These samples were previously recruited (2004–2013) according to the inclusion and exclusion criteria described by Mokone et al. (2005, 2006). In addition, a previously recruited (2011–2014) British Caucasian study group (Rickaby et al., 2015) consisting of 130 asymptomatic controls (UK CON group) and 108 participants with chronic Achilles tendinopathy (UK TEN) was included. The participants from each population were matched for country of birth (self-reported) and provided written informed consent. Furthermore, each participant completed questionnaires specifying personal details, injury details, and medical and sporting history. For the cases, age was recorded at time of injury and for the controls, at time of recruitment.
This study was approved by the Faculty of Health Sciences Research Ethics Committee within the University of Cape Town, South Africa and the Research Ethics Committee of the University of Northampton. This case–control genetic association study is in accordance with the recommendations of the STREGA Statement for reporting the results of genetic association studies (Little et al., 2009).
DNA extraction
For the South African participants, ∼5 mL of venous blood was collected from each patient by venipuncture of the forearm vein, into an ethylenediaminetetraacetic acid (EDTA) Vacutainer® tube. Samples were stored at −20°C before total DNA extraction using the standard protocol as described by Lahiri and Nurnberger (1991) with slight modifications (Mokone et al., 2006). For the British participants, 2 mL of saliva was obtained from each participant for DNA extraction as previously described (Rickaby et al., 2015). DNA extractions were performed at the University of Northampton, and the samples were made available for this study.
Single nucleotide polymorphism selection and genotyping
The five single nucleotide polymorphisms (SNPs) selected for analysis within the VEGFA (rs699947, rs1570360, rs2010963) and KDR (rs1870377 and rs2071559) genes were previously implicated with risk of ACL ruptures (Rahim et al., 2014). All samples (SA group n = 207 and UK group n = 238) were genotyped for the five polymorphisms using standard genotyping protocols. Polymerase chain reaction (PCR) and restriction fragment length polymorphism (RFLP) analysis was used for the VEGFA rs699947 (BglII) (Garza-Veloz et al., 2011) and KDR rs1870377 (AluI) SNPs. Details of the primers sequences and PCR conditions are available on request. Restriction fragments were resolved, together with a 100 bp size standard, by 2% agarose gel electrophoresis. Catalogued TaqMan™ Genotyping Assays (Applied Biosystems, Foster City, CA, USA) were used for the VEGFA rs1570360, VEGFA rs2010963, and KDR rs2071559 SNPs. The PCR reactions were conducted using the Applied Biosystems StepOnePlus™ Real-Time PCR system and the Applied Biosystems StepOnePlus Real-Time PCR software v2.2.2 (Applied Biosystems), following the manufacturer's recommendations.
Every PCR plate (RFLP and TaqMan analyses) contained negative controls (no DNA) and five repeat samples (known genotypes) as a quality control measure for reliable genotyping and detection of contamination. Genotypes were confirmed by two independent investigators, with an average genotyping success rate of 95%. Samples that failed twice to amplify during PCR for a particular SNP were considered unsuccessfully genotyped (Rahim et al., 2014). All genotyping was conducted at the Division of Exercise Science and Sports Medicine within the University of Cape Town.
Statistical analysis
Power calculations were calculated using QUANTO v1.2.4 (http://hydra.usc.edu/gxe) to determine sample size for the study. Assuming minor allele frequencies between 0.2 and 0.5, a sample size of 85 cases would be adequate to detect an allelic odds ratio (OR) of 2.0 and greater at a power of 80% and a significance level of 5%.
The programming environment R (R Development Core Team, 2010) was used for all data analysis. Basic descriptive statistics were compared using one-way analysis of variance to determine any significant differences between the characteristics of the CON versus TEN groups. The R packages genetics (Warnes et al., 2011) and SNPassoc (González et al., 2007) were used to analyze any differences in genotype and allele frequencies between the diagnostic groups (CON/TEN) and to calculate Hardy–Weinberg equilibrium (HWE) probabilities and linkage disequilibrium (LD). Haplotypes were inferred using the R package haplo.stats (Schaid et al., 2002; Sinnwell and Schaid, 2011). Analysis of the participant characteristics revealed potential confounding variables; so for this reason, the genotype and haplotype analyses were adjusted accordingly.
In addition to the analyses within the SA and UK study groups, the genotype and allele frequency distributions were compared between the countries (SA/UK), adjusting for age, weight, and diagnostic group and in the combined (SA+UK) groups, adjusting for age, weight, and country. Statistical significance was accepted when p < 0.05. No adjustments were made for multiple testing, as currently no obvious appropriate method exists (Nyholt, 2004; Perneger, 1998). The Bonferroni correction was considered too conservative for this study, as the tests were conducted on the same group of participants and several of the polymorphisms were in tight LD (Nyholt, 2004). Furthermore, correction for multiple testing was considered inappropriate, as there was an a priori hypothesis (Perneger, 1998; Posthumus et al., 2012).
Results
Participant characteristics
The participant characteristics for the SA and UK groups were previously described in earlier studies investigating the same study participants (Mokone et al., 2005, 2006; Rickaby et al., 2015). Briefly, the SA CON and SA TEN participants were unmatched for age, weight, and body mass index (BMI; Table 1). In the UK study group, the CON and TEN participants were matched for all characteristics except age (Table 2). There were no significant genotype effects on age, sex, height, weight, or BMI for any of the polymorphisms (Supplementary Table S1).
Values are expressed as mean ± standard deviation; sex and country of birth are represented as a percentage. The number of participants (n) with available data for each variable is in parentheses.
Age shows self-reported values at the time of onset of symptoms for the TEN group and at time of recruitment for the CON group.
p-Value adjusted for age and sex.
CON versus TEN, p-values in bold typeset indicates significance (p < 0.05).
BMI, body mass index; CON, control; SA, South African; TEN, Achilles tendinopathy.
Values are expressed as mean ± standard deviation; sex and country of birth are represented as a percentage. The number of participants (n) with available data for each variable is in parentheses.
Age, weight, and BMI are self-reported values at the time of onset of symptoms for the TEN group and at time of recruitment for the CON group.
CON versus TEN, p-values in bold typeset indicates significance (p < 0.05).
Genotype and allele frequencies
The VEGFA rs699947 genotype frequency was significantly different (p = 0.003) between the SA CON and SA TEN groups (Table 3). The CC genotype was significantly over-represented in the SA CON group (32%, n = 35) compared with the SA TEN group (17%, n = 14) (p = 0.019, OR: 2.30, 95% confidence interval [CI]: 1.14–4.64). Additionally, the CA genotype was significantly under-represented in the SA CON group (40%, n = 43) compared with the SA TEN group (61%, n = 49) (p = 0.005, OR: 2.32, 95% CI: 1.28–4.17). No other significant differences in genotype or allele frequency distributions were observed between the CON and TEN groups (Table 3) for any of the five polymorphisms investigated in either the South African or the British participants. The genotype and minor allele frequency distributions were similar to those reported for other European populations (www.ncbi.nlm.nih.gov/snp/) and for the previous ACL study by our group (Rahim et al., 2014). All the groups were in HWE except for the UK TEN group for the rs1570360 SNP (p = 0.020) and the rs1870377 SNP (p = 0.015).
Genotype and allele frequencies are expressed as a percentage, with the number of participants (n) in parentheses. p-Values are for differences in genotype and minor allele frequencies between countries (SA/UK) and diagnostic groups (CON/TEN), adjusted for one another. Differences between the diagnostic groups in the South African, British, and combined (SA+UK) groups are shown in the table. aIndicates p-values are adjusted for age and weight.
Indicates p-values are adjusted for age, and bold typeset indicates significance (p < 0.05). p-Values for the exact test of HWE for each of the groups are included in the table.
HWE, Hardy–Weinberg equilibrium; VEGFA, vascular endothelial growth factor, A isoform; KDR, kinase insert-domain receptor.
Comparisons in genotype and allele frequency distributions between the countries showed that only the KDR rs2071559 genotype frequency distribution was significantly different between the groups (p = 0.026) (Table 3). When the SA+UK groups were combined, no significant differences were observed in genotype or allele frequency distribution (Table 3). The linkage disequilibrium structure of the investigated SNPs within the VEGFA gene is presented in the Supplementary Figure S1.
Inferred haplotypes
Of the eight possible haplotype combinations constructed from the VEGFA variants (rs699947 C/A–rs1570360 G/A–rs2010963 G/C), only four (C-G-G, C-G-C, A-G-G, A-A-G) were inferred at a frequency >2%. The A-G-G inferred haplotype combination was significantly under-represented (Fig. 1A) in the SA CON group (15%, n = 18) versus the SA TEN group (20%, n = 22) (dominant model, p = 0.048; haplo.score: 1.97). Similarly, when the SA and UK groups were combined, the A-G-G haplotype was significantly under-represented in the SA+UK CON group (14%, n = 35) versus the SA+UK TEN group (20%, n = 39) (dominant model, p = 0.009; haplo.score: 2.61) (Fig. 1C). However, no significant differences were observed between the UK CON and UK TEN groups (Fig. 1B).
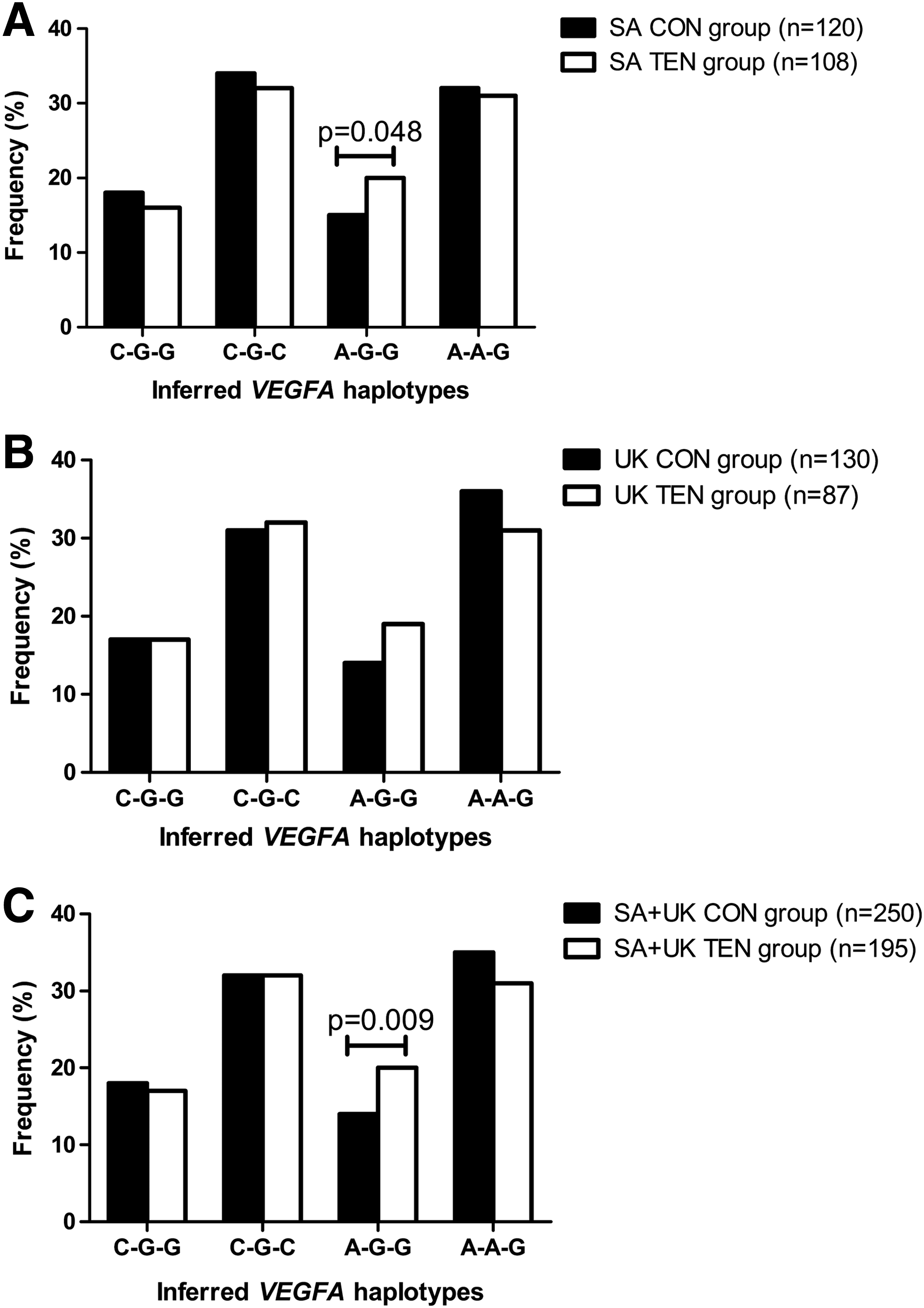
Inferred haplotype frequency distributions for the VEGFA rs699947, rs1570360, and rs2010963 polymorphisms in the control group (CON; black bars) and the Achilles tendinopathy group (TEN; white bars) for
Haplotypes were also inferred for the rs2071559 A/G and rs1870377 T/A polymorphisms within the KDR gene, but no significant differences in frequency distribution were observed between any of the groups after adjusting for the potential confounders (data not shown).
Discussion
The angiogenesis-associated signaling pathway is considered a key biological mediator to promote matrix remodeling. Therefore, this study investigated five polymorphisms within the VEGFA (rs699947, rs1570360, and rs2010963) and KDR (rs2071559 and rs1870377) genes, with risk of Achilles tendinopathy in two independent populations. The main finding of this article was the haplotype association of the three VEGFA SNPs with the risk of TEN in the SA group and the combined SA+UK group.
VEGF is a potent angiogenic factor, with a key role in the formation of new vasculature (Ferrara et al., 2003). The importance of VEGF in vasculogenesis and angiogenesis was demonstrated by Carmeliet et al. (1996) using a mouse gene knockout model, where loss of a single VEGF allele led to embryonic lethality. Consequently, an abundance of research has focused on the role of this growth factor in normal physiology, pathological conditions, and as a part of therapeutic strategies. To date, VEGF has been implicated in several pathological conditions, including cancer, macular degeneration, and rheumatoid arthritis (Evans, 2015). It is hypothesized that angiogenesis also occurs after mechanical loading in an effort to promote remodeling of the tendon ECM. This is further supported by studies identifying increased VEGF levels after mechanical loading of tenocytes (Mousavizadeh et al., 2014; Nakama et al., 2006; Petersen et al., 2004).
In the present study, the VEGFA A-G-G (rs699947 C/A–rs1570360 G/A–rs2010963 G/C) inferred haplotype was associated with increased risk of TEN. This association was noted in both the SA and the combined SA+UK groups, which provides further support implicating the VEGFA locus in modulating musculoskeletal injury susceptibility (Rahim et al., 2014). The VEGFA A-G-G haplotype combination is associated with decreased transcription of the VEGFA gene and reduced VEGF plasma levels (Lambrechts et al., 2003). Hence, it is possible that angiogenesis is limited in an already hypovascular structure, thus restricting the extent of matrix remodeling that is able to occur.
Tang et al. (2016) hypothesized that low VEGF activity may be partly responsible for the slow or weak healing capacity of injured tendons. Furthermore, the authors demonstrated that VEGF gene therapy via an adeno-associated viral type-2 (AAV2) vector resulted in marked improvements in tendon healing and remodeling in an animal model (Tang et al., 2016). Taken collectively, investigation into the role of VEGF in modulating both tendon capacity and injury susceptibility is warranted.
A recent study implicated the A-A-G inferred haplotype, with decreased risk of non-contact ACL ruptures (Rahim et al., 2014). The explanation for the contrasting haplotype associations observed between TEN and ACL ruptures is not completely understood. It is interesting to note that both these implicated haplotypes are associated with reduced VEGF plasma levels (Lambrechts et al., 2003). One could, therefore, hypothesize that because the vascularity of tendons and ligaments differs (Benjamin et al., 2006; Fenwick et al., 2002), the biological impact of low levels of VEGF on neovascularization related to remodeling and capacity would potentially be different between these functionally different tissues. Alternatively, the results may be indicating that these polymorphisms are potentially in linkage with a true risk variant. Thus, it is key that these genomic regions are further explored. Specifically, these associations need to be investigated at a functional level to determine the biological significance of these variants, and of angiogenesis in the repair model for TEN and ACL ruptures.
It is not surprising that the haplotype implicated in TEN risk modulation is reflected in the independent association of the rs699947 SNP genotype in the South African study group. Moreover, this promoter (-2578C/A) polymorphism is known to be functional, with the C allele correlating to increased VEGF protein production, and the AA genotype linked with lower transcriptional activity (Shahbazi et al., 2002) and increased risk in several phenotypes (Andraweera et al., 2012; Lambrechts et al., 2009; Seo et al., 2005). Therefore, there is evidence implicating the VEGFA gene locus and specific variants at a functional and genetic level with risk modulation.
Although no independent or haplotype associations were observed for the KDR gene, this remains a viable candidate for further investigation, particularly due to its importance in the VEGF signaling pathway. KDR is one of the two main receptors that VEGFA binds to and is responsible for the main downstream signaling effects of the ligand (Ferrara et al., 2003). Moreover, KDR null mutant mice were observed to be embryonically lethal (Shalaby et al., 1995). Therefore, it may be worth exploring additional genomic loci within this candidate gene, as the two polymorphisms (rs2071559 and rs1870377) investigated in the current study are situated in the promoter and exon 11, respectively.
A limitation of the study was that SA participants were not matched for age, weight, or BMI. We were unable to determine the weight at time of injury for the cases due to incomplete data but several participants reported an increase in weight due to injury-related inactivity. Additionally, the older age of the cases may have influenced the weight of the cases. In the UK participants, the CON and TEN groups were also unmatched for age and for this reason, all statistical analyses were adjusted for the confounders.
Although the UK TEN group deviated from HWE at the rs1570360 and rs1870377 loci, the genotype and minor allele frequency distributions were similar to those reported for other European populations. The main limitation of this study was the small sample size, and, therefore, the SA and UK study groups were also combined for the statistical analyses. A minimum sample size of 85 cases was adequate to detect only allelic ORs of 2.0 and greater, whereas the combined SA+UK group sample set was sufficient to detect allelic ORs of 1.6 and greater.
Conclusions
Our observations collectively suggest that a genomic interval within the VEGFA gene was associated with risk of TEN. This gene requires further exploration to understand the potential role of VEGFA and specifically angiogenesis in the etiology of TEN. Investigation into genes involved in the angiogenesis-associated signaling pathway may aid in the development of a risk profile for tendinopathy. In line with this, our future work is aiming toward employing next-generation technologies, specifically whole exome sequencing, to further explore these regions of interest in an effort to further characterize these genomic signatures.
The era of “precision medicine” is becoming a large focus underpinning research (Higdon et al., 2015; Poland et al., 2011). The technologies of the omics era has revolutionized our capacity to unravel these complex phenotypes. Through Big Data emerging from multi-omics research streams at genomics, proteomics, transcriptomics, epigenomics, and bioinformatics levels, we can advance the depth and breadth of research in the field of Sports and Exercise Medicine, with a view to improving global health, preventive medicine, and quality of life.
Footnotes
Acknowledgments
The authors would like to thank Mr. George Mokone and Dr. Martin Schwellnus for SA participant recruitment. M.R. was funded by the University of Cape Town and the National Research Foundation. M.P. was funded by the Thembakazi Trust. This research was funded in part by the University of Cape Town Research Council, the National Research Foundation, and the Department of Science and Technology/National Research Foundation Collaborative Grant (NRF92550). Authors M.C. and A.V.S. have filed patents on the application of specific sequence variations (not included in this article) related to risk assessment of ACL ruptures and Achilles tendinopathy.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.