
Abstract

To the Editor:
Carcinogenesis can be defined as the acquisition of certain properties by cells that make them immortal and provide them with unlimited proliferation capacity. Polyploidy and deregulation of gene expression are two characteristics of carcinogenesis. These two processes are most often associated with a denaturation of chromatin and especially that of heterochromatin, which constitutes the signature of most solid tumors (Yuen et al., 2005).
Heterochromatin is a tightly packed form of DNA. It is comprised of DNA wrapped around octamers of histones called nucleosomes. These nucleosomes are tightened together and maintained by the action of several proteins and histone marks (Fig. 1A). Proteins such as methyl transferases, demethylases, histone acetyltransferases (HATs), and deacetylases allow the stability of different chromatin structures.
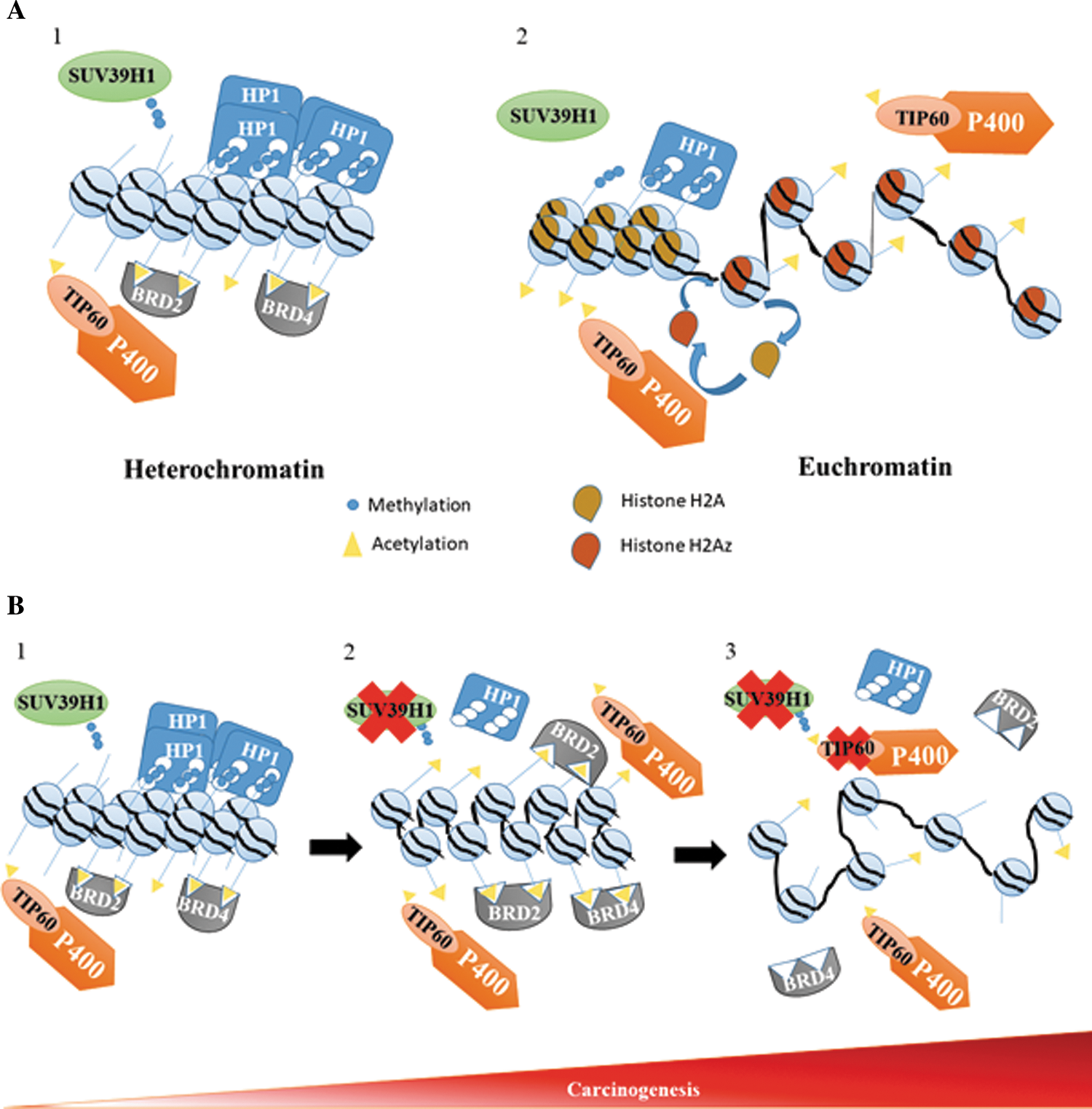
The methyl transferase suppressor of variegated 3–9 homolog 1 (SUV39H1), through the trimethylation of N-terminal tail of histone H3 on lysine 9 (H3K9me3), allows the binding of the heterochromatin protein 1 (HP1) that will preserve the structure of the heterochromatin (Fig. 1A).
In many cancers such as breast and colorectal cancers, it has been shown that there is a loss of SUV39H1 and the H3K9me3 biomarkers. However, it was shown that Tip60 intervened at the level of heterochromatin to compensate for the absence of SUV39H1. Tip60 would acetylate histone H4 on its lysine 12 (H4k12ac) at the heterochromatin, which would allow proteins with bromodomains such as bromodomain-containing protein 2 or 4 (BRD2/4) to settle and thus maintain this structure (Fig. 1A). This TIP60 action could involve the Tip60 complex, with the intervention of ATPase P400 and balance of the Tip60/p400 ratio (Mattera et al., 2009).
Acetyltransferase Tip60 is a protein capable of attaching an acetyl group to other proteins, such as transcription factors and histones. This acetylation activates proteins, and in the case of histones, it has always been associated with the opening of chromatin allowing gene expression (Fig. 1A).
Recent studies have shown the presence of histone acetylation H3 and H4 in the heterochromatin. Acetylation of histone H4 lysine 12 in heterochromatin would maintain a basal transcription level and a plasticity of the heterochromatin by binding bromodomain proteins BRD2 and BRD4 to this mark (Fig. 1B). Indeed, heterochromatin is the condensed part of the chromosomes, where genes are fewer or not expressed. It is also the genomic location where most of the oncogenes are nested. A skeleton composed of SUV39H1-H3K9me3-HP1 under normal conditions most often maintains this structure (Fig. 1B). Grezy et al. showed that when this normal compaction pathway is defective, HAT Tip60 is recruited on heterochromatin where it mediates histone acetylation H4K12 and BRD2/4 binding. This new pathway is the last step for the denaturation of heterochromatin (Fig. 1B).
In most cancers, there is variation in the transcriptional and/or translational expression of Tip60. For example, in colorectal and breast cancers, there are transcriptions and translations under expression of Tip60, followed by a loss of the Tip60-H4K12ac-BRD2/4 pathway. This leads the cells to a process of carcinogenesis with a loss of the chromosomal architecture and including polyploidy (Fig. 1B).
The presence of Tip60 at the heterochromatin is even more possible, given the presence of P400 in the heterochromatin to incorporate the histone variant H2Az in the G1 phase in the normal cell (Fig. 1A). P400 is an ATPase involved in chromatin remodeling through its SNF2 catalytic domain common to all chromatin remodeling ATPase (Fuchs et al., 2001). The enzymatic activity of this domain allows incorporation of the histone variant H2Az instead of the histone H2A (Fig. 1A).
Indeed, Grezy et al. observed a denaturation of heterochromatin in SUV39H1/2−/− cells after inhibition of P400 by small interfering RNA similar to the inhibition of TIP60, which suggests that Tip60 is being recruited within its complex by P400. However, other studies conducted on human fibroblasts have shown that the depletion of P400 or histone HA2z variant by short hairpin RNA could induce heterochromatin formation that would be associated with cellular senescence (Gévry et al., 2007). Under normal conditions, P400 may allow chromatin release (Fig. 1A). However, the loss of the SUV39H1-H3K9me3-HP1 skeleton allows the recruitment of Tip60 and the implementation of the new heterochromatin maintenance pathway Tip60-H4K12ac-BRD2/4 (Fig. 1B).
Mattera et al. (2009) have shown another link between Tip60, P400, and carcinogenesis. They demonstrated that the Tip60/P400 ratio is a key factor toward inducing carcinogenesis. It was shown that the variation in the Tip60/P400 ratio in favor of P400 resulted in a decrease in Tip60 expression and carcinogenesis progression, whereas a variation in this ratio in favor of Tip60 leads to apoptosis of tumor cells. Indeed, it has been shown that P400 is capable of binding its SANT domain to the Tip60 HAT domain, thus P400 inhibits the acetyltransferase activity of Tip60 (Park et al., 2010). This suggests that P400 would be a Tip60 regulator when the latter is underexpressed, as in the case of colorectal or breast cancer (Mattera et al., 2009).
Taken together, we shall also note that Tip60 is a pleiotropic protein involved in several physiological processes, such as transcriptional regulation, DNA repair, cell migration, invasion, alteration of chromatin structure, and genomic instability (Judes et al., 2015). Its involvement in these different processes and the small trip to the land of heterochromatin makes Tip60 an ideal target for a deeper understanding of carcinogenesis and a worthwhile therapeutic target for cancer therapeutics and diagnostic innovation.
Footnotes
Acknowledgment
K.R. is the recipient of a grant from the Hariri Foundation.
Author Disclosure Statement
The authors declare there are no competing financial interests.