
Abstract
Abstract
Shisha (water pipe) smoking is falsely believed to be a hazard-free habit and has become a major public health concern. Studies have reported shisha smoking to be associated with oral lesions, as well as carcinomas of the lung, esophagus, bladder, and pancreas. A deeper understanding of the underlying molecular mechanisms would contribute to identification of biomarkers for targeted public health screening, therapeutic innovation, and better prognosis of associated diseases. In this study, we have established an in vitro chronic cellular model of shisha-exposed oral keratinocytes to study the effect of shisha on oral cells. Normal nontransformed, immortalized oral keratinocytes were chronically exposed to shisha extract for 8 months. This resulted in significant increase in cellular proliferation and cell invasion in shisha-exposed cells compared to the parental cells. Quantitative proteomic analysis of OKF6/TERT1-Parental and OKF6/TERT1-Shisha cells resulted in the identification of 5515 proteins. Forty-three differentially expressed proteins were found to be common across all conditions. Bioinformatic analysis of the dysregulated proteins identified in the proteomic study revealed dysregulation of interferon pathway, upregulation of proteins involved in cell growth, and downregulation of immune processes. The present findings reveal that chronic exposure of normal oral keratinocytes to shisha leads to cellular transformation and dysregulation of immune response. To the best of our knowledge, this is the first report that has developed a model of oral keratinocytes chronically exposed to shisha and identified proteomic alterations associated with shisha exposure. However, further research is required to evaluate the health burden of shisha smoking.
Introduction
Oral cancer is a global health problem. The worldwide incidence of cancers of the oral cavity, lip, and pharynx is expected to rise from the current 529,500 cases to 856,000 cases by 2035 (Shield et al., 2017). The 5-year survival rate of oral cancer patients is less than 60%. Many risk factors have been associated with oral squamous cell carcinoma (OSCC). Some of the prime factors include tobacco and alcohol use, viral infections, and candidiasis (Ng et al., 2017). Tobacco-related deaths are projected to rise from 6.4 million in 2015 to 8.3 million in 2030.
Smoking is the most common form of tobacco use in the form of cigarettes, bidis, and hookah. Shisha, also known as hookah or water pipe, has been used for smoking tobacco in Africa and Asia for over four centuries. The prevalence of shisha use has increased in Middle East in the past few decades (Munshi et al., 2015). OSCC was known to primarily occur in males in their sixth to seventh decades; however, in the past few decades, there has been a shift in the OSCC incidence toward the younger demographic (Llewellyn et al., 2004). The availability of flavored tobacco and mushrooming of hookah bars have popularized the use of hookah among youth, which is likely to contribute to this shifting demographic (Maziak et al., 2015). Hookah smoking is less stigmatized compared to smoking cigarettes, and common among both men and women (Dar-Odeh and Abu-Hammad, 2009).
Notably, shisha use has been perceived to be less harmful than cigarette smoking since the tobacco smoke gets filtered through water before inhalation (Aljarrah et al., 2009; Kiter et al., 2000). However, studies indicate that shisha users are exposed to higher levels of carbon monoxide, nicotine, tobacco-specific nitrosamines, polycyclic aromatic hydrocarbons, and volatile aldehydes compared to cigarette smokers (Daher et al., 2010). In addition, the amount of formaldehyde and acetaldehyde released from a single session of shisha smoking equals that of 17 cigarettes and 5 cigarettes, respectively (Al Rashidi et al., 2008).
Shisha smoking has been reported to be associated with carcinomas of the lung, esophagus, bladder, and pancreas (Dar-Odeh and Abu-Hammad, 2009). Studies from Saudi Arabia, Yemen, and India have assessed the association of shisha use with oral lesions. However, these studies were not uniformly consistent (Al-Attas et al., 2014; Dangi et al., 2012; Schmidt-Westhausen et al., 2014). Centers for Disease Control and Prevention (CDC) reports that shisha smoking may increase the risk of developing oral cancers. However, the mechanisms of action of shisha on oral cells have not been elucidated. A deeper understanding of underlying mechanisms may facilitate identification of biomarkers for early detection, as well as therapeutic intervention of oral cancer associated with shisha use.
To achieve this, we developed a model of long-term cellular exposure, using normal, nontransformed oral keratinocytes that were chronically exposed to shisha to study shisha-associated changes in normal nontransformed immortalized oral cells. We observed that chronic exposure to shisha led to increased cellular proliferation and induced invasive ability in the noninvasive oral keratinocytes. In addition, we studied the proteomic profile of the OKF6/TERT1 cells chronically exposed to shisha compared to unexposed cells. To our knowledge, this is the first study that has developed a model of oral keratinocytes chronically exposed to shisha and identified molecular alterations associated with shisha exposure using proteomics.
Materials and Methods
Preparation of shisha extract
Shisha was obtained commercially from a local supplier. Five grams of shisha was crushed well using a mortar and pestle and suspended in 50 mL of 1 × phosphate-buffered saline (PBS), as described previously (Nanjappa et al., 2015; Rohatgi et al., 2005). The mixture was kept at 37°C overnight in a shaker incubator, followed by centrifugation at 12,000 rpm for 20 min. The supernatant was filtered using a Whatman filter paper and sterilized using a syringe-driven 0.22 μm cutoff filter. The shisha extract was made into aliquots and stored at −80°C until further use.
Cell culture
Nontransformed, immortalized, normal human oral keratinocytes, OKF6/TERT1, used in this study, was a generous gift from Dr. James Rheinwald (Brigham and Women's Hospital, Boston, MA). OKF6/TERT1 were cultured and maintained in keratinocyte serum-free medium supplemented with bovine pituitary extract (25 mg/mL), epidermal growth factor (0.2 ng/mL), 1% penicillin/streptomycin, and CaCl2 (0.4 mM). The cells were grown at 37°C in a humidified 5% CO2 incubator.
Exposure of OKF6/TERT1 cells with shisha extract
OKF6/TERT1 cells were chronically exposed with 0.5% shisha extract for 8 months. Henceforth, the parental cells will be referred to as “OKF6/TERT1-Parental or Parental cells,” and OKF6/TERT1 cells chronically exposed with shisha extract for 2, 4, 6, and 8 months will be termed as “OKF6/TERT1-Shisha-2 month, OKF6/TERT1-Shisha-4 month, OKF6/TERT1-Shisha-6 month, and OKF6/TERT1-Shisha-8 month,” respectively. All cells were grown at 37°C in a humidified 5% CO2 incubator.
Cell proliferation assays
OKF6/TERT1 cells and OKF6/TERT1-Shisha cells exposed for 2, 4, 6, and 8 months were seeded in a 96-well plate at a density of 5 × 103 cells/well. Cell viability was determined every 24 h using 3-(4, 5-dimethylthiazol-2yl)-2, 5-diphenyl tetrazolium bromide (MTT) assays as previously described (Chatterjee et al., 2006). All experiments were carried out in triplicate. Paired t-test was performed to evaluate the difference between the control and exposed groups. p Value <0.05 was considered to be significant.
Cell invasion assays
Invasion assays were carried out in a transwell system for 48 h. The transwell system consists of cell culture inserts with translucent polyethylene terephthalate membrane 8 μm pores (catalog no. 353097; BD Biosciences, San Jose, CA) placed in 24-well plate. The upper surface of the insert membrane was coated with Matrigel (catalog no. 354234; BD Biosciences). OKF6/TERT1 cells and OKF6/TERT1-Shisha cells exposed for 2, 4, 6, and 8 months were suspended at a density of 2 × 104 cells/well in supplement-free media in the upper compartment of the transwell. Complete media was added to the lower compartment. The cells were incubated at 37°C in 5% CO2 incubator for 48 h.
At the end of the incubation time, the upper surface of the insert membrane was wiped with a cotton-tip applicator to remove nonmigratory cells. Cells that have invaded the Matrigel to the lower surface of the membrane were fixed and stained with 4% methylene blue (Sigma, St. Louis, MO) in 50% methanol. All experiments were carried out in triplicates. The insert membrane with the invaded cells was imaged at 10 × magnification, and the number of cells invaded was counted using ImageJ. Paired t-test was performed to evaluate the difference between the control and shisha-exposed groups. p Value <0.05 was considered to be significant.
Protein extraction
OKF6/TERT1-Parental cells and OKF/TERT1-Shisha cells (2, 4, 6, and 8 months) were grown to 80% confluence. The cells were rinsed thrice with 1 × PBS and starved in supplement-free media containing 1% penicillin–streptomycin for 8 h. The cells were then rinsed thrice with 1 × PBS and harvested in 2% sodium dodecyl sulfate (SDS)-containing buffer (2% SDS, protease inhibitors [Roche, Indianapolis, IN] in 50 mM triethyl ammonium bicarbonate [TEABC]). The cell lysates were sonicated at 40% amplitude and centrifuged at 12,000 rpm for 10 min. Protein concentration was measured using bicinchoninic acid assay method (Thermo Scientific, Bremen, Germany) (Smith et al., 1985). Equal amounts of protein from each cell line were reduced using dithiothreitol (DTT) at 60°C for 20 min and alkylated with iodoacetamide (IAA) for 10 min at room temperature.
Protein was precipitated overnight at −80°C using the ice-cold acetone. The samples were centrifuged at 12,000 rpm for 15 min; the acetone was removed and air-dried. The pellet was then dissolved in 4 M urea. Proteins were then digested using Lysyl Endopeptidase®, Mass Spectrometry Grade (catalog no. 125-05061; Wako, Richmond, VA) at 1:100 enzyme to protein ratio for 4 h at 37°C. After 4 h, the urea concentration was reduced from 4 to 2 M using 50 mM TEABC. The samples were then digested using tosyl phenylalanyl chloromethyl ketone (TPCK)-treated trypsin (Worthington, NJ) at a 1:20 enzyme to protein ratio for 16 h at 37°C. The samples were cleaned using Sep-Pak Classic C18 columns (catalog no. WAT051910; Waters, Milford, MA).The samples were then completely dried and reconstituted in 50 mM TEABC buffer.
Tandem mass tag labeling and basic reversed-phase liquid chromatography
Peptides from each sample were differentially labeled using tandem mass tag (TMT) labeling reagents as per manufacturer's instructions (catalog no. 90110; Thermo Fisher Scientific, Bremen, Germany). Peptides were labeled with TMT channels as follows: OKF6/TERT1-Parental cells with 128N, OKF6/TERT1-Shisha 2 months with 128C, OKF6/TERT1-Shisha 4 months with 129N, OKF6/TERT1-Shisha 6 months with 129C, and OKF6/TERT1-Shisha 8 months with 130N. The TMT-labeled peptides were subjected to basic reversed-phase liquid chromatography fractionation as described previously (Nagarajha Selvan et al., 2014). The 96 fractions obtained were concatenated into 6 fractions. The fractions were vacuum dried, desalted using C18 Stage Tips, and analyzed using liquid chromatography tandem mass spectrometry (LC-MS/MS).
LC-MSn analysis
The peptide fractions were reconstituted in 0.1% formic acid and analyzed on Orbitrap Fusion™ Tribrid™ mass spectrometer (Thermo Electron, Bremen, Germany) interfaced with Easy-nLC 1000 nanoflow liquid chromatography system (Thermo Scientific). Peptides were loaded onto a trap column (75 μm × 2 cm, Magic C18AQ, 5 μm, 100 Å; Michrom Biosciences, Inc., Auburn, CA) using 0.1% formic acid at a flow rate of 3 μL/min. The peptides were then resolved on an analytical column (75 μm × 20 cm, Magic C18AQ, 3 μm, 100 Å; Michrom Biosciences, Inc.) at a flow rate of 300 nL/min using 8–25% solvent B (0.1% formic acid in 95% acetonitrile) for 3 to 73 min and 25–30% solvent B for 73 to 103 min.
The MS and MS/MS scans were acquired at a mass resolution of 120,000 and 50,000 at 200 m/z. Full MS scans were acquired in the m/z range of 400–1600. For MS/MS, top speed with a cycle time of 3 sec was selected for each duty cycle with charge ≥2 sequentially isolated in each cycle. Precursor ions with single charge or unassigned charge were rejected. Dynamic exclusion of fragmented precursor ions was set to 40 sec. Precursor ions were isolated using quadrupole mass filter using isolation width of 1.6 m/z. Higher energy collision dissociation was used as the fragmentation method for fragmentation with 34% normalized collision energy. The peptide fractions were analyzed thrice on the mass spectrometer (three technical replicates).
Data analysis
The raw data obtained were searched against human protein database NCBI RefSeq (Release 81) using Sequest search algorithm on a Proteome Discoverer (version 2.1) software suite (Thermo Fisher scientific). The search parameters included: Charge state 2–5, trypsin as the proteolytic enzyme with an allowance of up to two missed cleavages, oxidation of methionine as the dynamic modification, and carbamidomethyl at cysteine and TMT 6-plex modification at N-terminus of the peptide and lysine as the static modifications.
The acquired data were searched against decoy database to obtain the false discovery rate (FDR). Peptide spectrum matches at 1% FDR were considered for peptide identification. TMT quantitation was done using reporter ion intensities. The ratio was calculated as follows: 128C (OKF6/TERT1-Shisha-2 month)/128N (OKF6/TERT1-Parental), 129N (OKF6/TERT1-Shisha-4 month)/128N (OKF6/TERT1-Parental), 129C (OKF6/TERT1-Shisha-6 month)/128N (OKF6/TERT1-Parental), and 130N (OKF6/TERT1-Shisha-8 month)/128N (OKF6/TERT1-Parental).
Accessibility of proteomic data
The mass spectrometry-based raw data have been submitted to ProteomeXchange Consortium through the PRIDE public repository (Vizcaino et al., 2016) and can be accessed using the data identifier—PXD011106.
Bioinformatic analysis
Gene ontology (GO) enrichment was carried out using FunRich tool (Version 3.1.3) (https://www.FunRich.org) (Pathan et al., 2015).
Principal component analysis (PCA) of all quantified proteins in OKF6/TERT1-Parental, OKF6/TERT1-Shisha-2 month, OKF6/TERT1-Shisha-4 month, OKF6/TERT1-Shisha-6 month, and OKF6/TERT1-Shisha-8 month and Welch's t-test statistic of all differentially regulated proteins were calculated using Perseus (Version 1.6) (Tyanova et al., 2016). Heat map was generated using Perseus (Version 1.6) (Tyanova et al., 2016). Volcano plot was generated by carrying out the pair-wise comparison between OKF6/TERT1-Parental and OKF6/TERT1-Shisha-8 month cells using R 3.4.0 (Team, 2008).
Results
Chronic exposure to shisha extract induces phenotypic changes in oral keratinocytes
Nontransformed, immortalized, normal human oral keratinocytes, OKF6/TERT1, were exposed with varying concentrations of shisha extract ranging from 0% to 2% to ascertain the optimum concentration for chronic exposure to shisha (data not shown). The highest concentration of shisha extract with which the cells could be chronically exposed without notable cytotoxicity was 0.5%. OKF6/TERT1 cells were chronically exposed with 0.5% shisha extract for 8 months. Chronic exposure to shisha extract resulted in changes in cellular morphology of OKF6/TERT1 cells. OKF6/TERT1-Shisha cells were found to be more elongated and spindle shaped than the OKF6/TERT1-Parental cells (Fig. 1A). This change in cell morphology was observed after 2 months of shisha exposure and remained consistent till 8 months of shisha exposure.

Chronic exposure to shisha alters cell morphology and increases cellular proliferation and invasive ability in oral keratinocytes.
We further assessed the phenotypic changes observed during oncogenic transformation of cells, including cell proliferation and invasive ability of cells. OKF6/TERT1-Shisha cells showed increased cell proliferation compared with OKF6/TERT1-Parental cells (Fig. 1B). A progressive increase in rate of cellular proliferation was observed among OKF6/TERT1-Shisha cells with a steady increase from OKF6/TERT1-Shisha-2 month to OKF6/TERT1-Shisha-8 month.
In vitro invasion assays using Matrigel were carried out to investigate the metastatic potential of shisha-exposed cells. OKF/TERT1-Shisha cells showed an increased invasive ability compared to the OKF6/TERT1-Parental cells (Fig. 1C, D). This increase in invasion steadily increased from 2 months of shisha exposure to 8 months of shisha exposure.
These phenotypic changes indicate an oncogenic transformation in nontransformed, immortalized, normal human oral keratinocytes upon chronic exposure to shisha extract. To better understand the underlying molecular events associated with chronic exposure of oral keratinocytes with shisha extract, we studied proteomic alterations in OKF6/TERT1-Shisha cells compared to OKF6/TERT1-Parental cells in a temporal setting.
Chronic exposure to shisha extract results in global proteomic changes
We studied the altered proteome of OKF6/TERT1-Shisha cells compared to OKF6/TERT1-Parental cells in a temporal manner using TMT-based quantitative proteomics. The experimental workflow is depicted in Figure 2A. Temporal proteomic analysis led to the identification of a total of 5515 proteins in all conditions across all three replicates. A complete list of identified proteins and peptides is provided in Supplementary Table S1. A heat map depicting all proteins identified in this study has been provided in Figure 2B. Of these 5515 proteins, 82, 77, 106, and 110 proteins were found to be differentially expressed (1.5-fold) in OKF6/TERT1 cells chronically exposed to shisha extract for 2, 4, 6, and 8 months, respectively, compared with OKF6/TERT1-Parental.
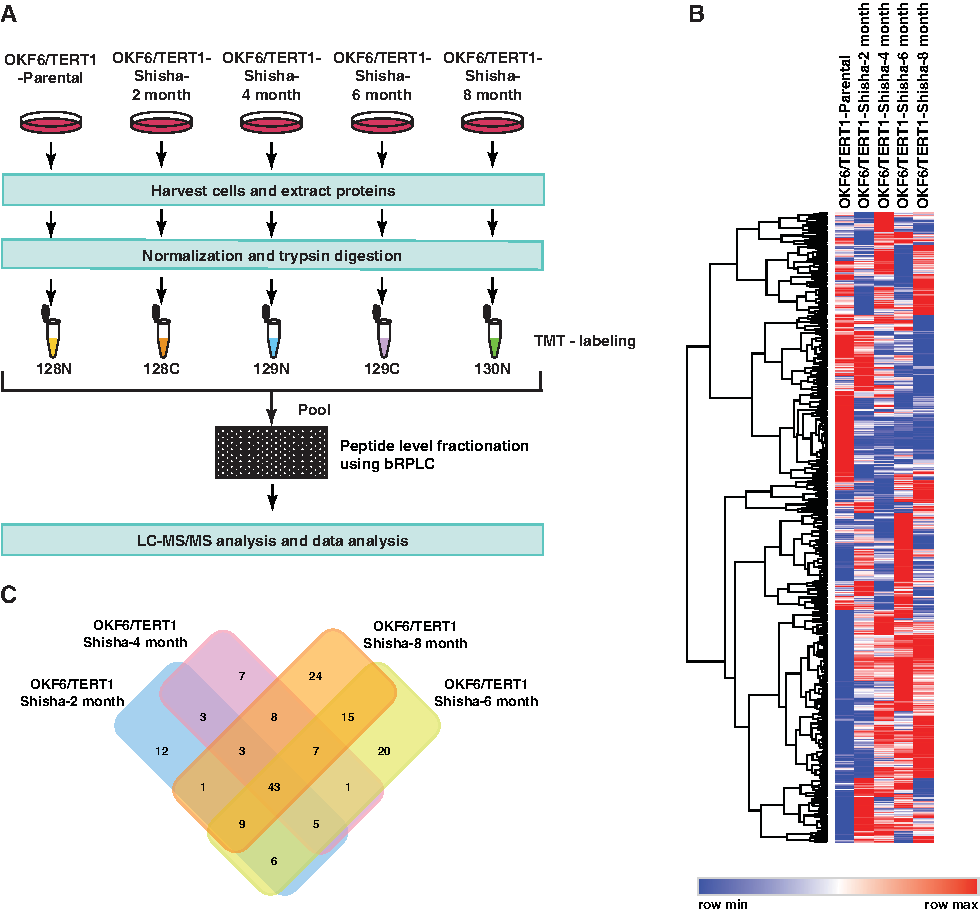
Experimental design and proteomic results.
Of these, 43 proteins were found to be differentially expressed across all conditions in all three replicates (Fig. 2C and Supplementary Table S2). A partial list of differentially expressed proteins has been provided in Table 1. Our data revealed progressive overexpression of 10 proteins and progressive downregulation of 27 proteins across the different time points of shisha exposure compared to the OKF6/TERT1-Parental cells (Fig. 3A). A Volcano plot representing the fold change distribution of the differentially expressed proteins in OKF6/TERT1-Shisha-8 months is provided in Figure 3B. PCA using Perseus software (Tyanova et al., 2016) revealed distinct variation between OKF/TERT1-Parental and OKF6/TERT1-Shisha cells (Fig. 3C).

Bioinformatic analysis of differentially regulated proteins on chronic exposure to shisha in a temporal manner.
A Partial List of Differentially Expressed Proteins Identified in OKF6/TERT1-Shisha Cells Across Time Points Compared to OKF/TERT1-Parental Cells
Chronic exposure to shisha extract increases pathways associated with cell growth and invasion and decreases immune response
FunRich was used to carry out GO-based analysis of proteins dysregulated in OKF6/TERT1-Shisha-8 month cells to gain insights into the altered biological pathways, biological processes, and cellular component. Biological process characterization of the proteins dysregulated in OKF6/TERT1-Shisha-8 month revealed upregulated proteins to be involved in cell growth and maintenance (17.9%, p < 0.05) and the downregulated proteins to be engaged in immune response (16.7%, p < 0.05) (Fig. 4A, B). Biological pathway analysis was performed for the proteins dysregulated in OKF6/TERT1-Shisha-8M. Data analysis revealed enrichment of immune response related signaling cascades in OKF6/TERT1-Shisha-8 month.
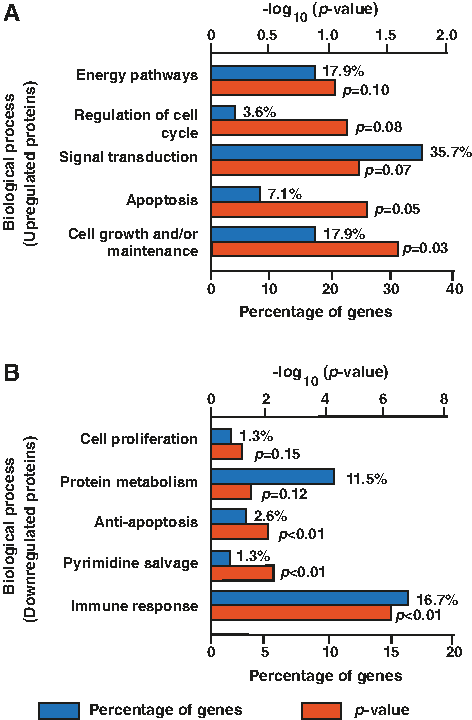
Gene-ontology based functional annotation of proteins dysregulated in OKF6/TERT1-Shisha-8M cells using FunRich.
We identified proteins that comprised 40% of the genes involved in immune response related signaling, including growth factor receptor bound protein 2 (GRB2), signal transducer and activator of transcription 1 (STAT1), signal transducer and activator of transcription 2 (STAT2), interferon-induced protein with tetratricopeptide repeats 2 (IFIT2), intercellular adhesion molecule 1 (ICAM), and HLA class I histocompatibility antigen, A-1 alpha chain (HLA-A) among others.
Carcinogens in tobacco smoke have been known to form DNA adducts and silence tumor suppressor genes resulting in loss of regulation of cell growth, formation of precancerous lesions, and metastatic disease (Forum 2013). We observed upregulation of proteins involved in cellular growth, including profilin 2 (PFN2, 1.6-fold) and tumor protein translationally controlled 1 (TPT1, 1.5-fold). Earlier studies have reported these proteins to be overexpressed in esophageal squamous cell carcinoma (ESCC) and head and neck squamous cell carcinoma (HNSCC), which in turn is a predictor of poor overall survival in these cancer patients (Bommer, 2017; Cui et al., 2016; Liu et al., 2018). Representative spectra of PFN2 and TPT1 are provided in Figure 5A and B.

Representative MS/MS spectra of overexpressed proteins
Tobacco use is known to increase oxidative stress through production of reactive oxygen species (ROS). It is also known to impair the defensive mechanisms of antioxidant systems. We found the downregulation of one such antioxidant enzyme, superoxide dismutase 2 (SOD2, 0.6-fold), in shisha-exposed cells.
Immune dysfunction has been observed to be an early event associated with carcinogenesis and tumor progression (Staveley-O'Carroll et al., 1998). We observed downregulation of proteins engaged in immune response, including HLA-A (0.6-fold), deoxynucleoside triphosphate triphosphohydrolase (SAMHD1, 0.4-fold), and 2′-5′-oligoadenylate synthase 3 (OAS3, 0.5-fold). Representative spectra of HLA-A and OAS3 are provided in Supplementary Figure S1A and B.
Cellular component analysis of dysregulated proteins observed in OKF6/TERT1-Shisha-8 month cells using FunRich revealed upregulated proteins to be localized in the lysosomes (51.9%, p < 0.01) and cytoplasm (77.8%, p < 0.01), while downregulated proteins were mainly localized to the cytoplasm (69%, p < 0.01) (Supplementary Fig. S2A, B).
Discussion
Tobacco use is one of the leading causes of morbidity worldwide. Cigarette smoking and chewing tobacco are the commonly used forms of tobacco. However, another form of tobacco use through shisha smoking has seen an upward global trend in the past decade (Munshi et al., 2015). Several investigations have confirmed various adverse effects associated with shisha smoking, including inflammation, Candida infection, low birth weight, periodontitis, oral premalignant lesions, and cancer (Munshi et al., 2015).
The precise molecular alterations in response to shisha smoke can be best appreciated by chronic exposure to carcinogenic insult, >rather than acute. We and others have elucidated the effect of chronic exposure to tobacco in the form of cigarette smoke or chewing tobacco, on multiple cell types using in vitro models (Bhat et al., 2018; Chang et al., 2010, 2011; Nanjappa et al., 2015; Rajagopalan et al., 2018a, 2018b). Even though the adverse effects of shisha smoking have been well explored, the molecular level changes induced by chronic exposure of normal oral keratinocytes to shisha have not been elucidated.
In this study, we have developed a model of oral keratinocytes chronically exposed to shisha extract and systematically studied the proteomic alterations in response to chronic exposure to shisha. Using this cellular model, we demonstrate early phenotypic alterations associated with oncogenic transformation in normal oral keratinocytes chronically exposed to shisha extract. These changes were evidenced by the temporal increase in cell proliferation and invasive ability of the oral keratinocytes on chronic exposure to shisha.
Furthermore, we used quantitative proteomics to identify dysregulated proteins that could be associated with the hallmarks of tumor progression in oral keratinocytes upon chronic exposure to shisha. Tobacco use through smoking has been associated with tissue injury and changes at the molecular level which, in turn, lead to cellular proliferation and transformation into cancer cells (Services, 2010). Bioinformatic analysis of the dysregulated proteins identified in this study revealed an enrichment of the biological processes associated with cell growth and maintenance. Among these proteins, we identified PFN2 and TPT1 to be upregulated in the OKF6/TERT1-Shisha-8 month compared to the OKF6/TERT1-Parental cells. PFN2 is an actin-binding protein that is involved in cell motility and regulation of actin polymerization.
Recent studies have shown a role of dysregulated PFN2 in progression of cancer. Upregulation of PFN2 has been associated with epithelial-to-mesenchymal transition followed by tumor cellular proliferation, invasion, migration, and metastasis in several cancers, including ESCC (Cui et al., 2016), lung cancer (Tang et al., 2015), colorectal cancer (Kim et al., 2015), and HNSCC (Liu et al., 2018). High expression of PFN2 has also been reported to predict poor overall patient survival in primary HNSCC and unfavorable prognosis in ESCC (Cui et al., 2016; Liu et al., 2018).
TPT1 is a p53 target gene that promotes cell survival (Chen et al., 2013). TPT1 has also been termed to be a cytoprotective protein that is involved in response to cellular stress such as oxidative stress, involved in DNA damage repair, promotes cell division and cell growth, and modulates immune response (Bommer, 2017). TPT1 has been reported to be associated with cancer progression, invasion, and metastasis (Bommer, 2017). A study reported higher transcriptional levels and overexpression of TPT1 in tumor tissues of oral cancer patients, and miR-27b-regulated TPT1 was put forward as a novel biomarker for oral cancer (Lo et al., 2012).
Overexpression of TPT1 was also found to be associated with colon cancer (Bommer et al., 2015) and skin squamous cell carcinoma (Wu et al., 2012). These results indicate that chronic exposure to shisha induces overexpression of key proteins involved in cell growth and maintenance, as well as invasion. This, in turn, potentially accounts for an increased cell proliferation and invasion observed in oral keratinocytes on chronic exposure to shisha observed in our study.
TPT1 has also been identified to be associated with reducing oxidative stress and promoting cell survival on generation of free radicals (Bommer, 2017; Chen et al., 2013). Tobacco use is known to induce oxidative stress through generating free radicals. These ROS are scavenged by antioxidant enzymes, including SOD1, SOD2, and glutathione peroxidase among others (Patel et al., 2008). However, SOD2 has been studied to have a dichotomous role in cancer, in which it can function either as a tumor suppressor or a tumor initiator. A loss of SOD2 expression is frequently observed in early lesions of tumor during tumor initiation. However, the expression of SOD2 elevates during tumor progression as the tumor becomes metastatic (Dhar et al., 2011; Kim et al., 2017; Robbins and Zhao, 2014). In our study, we found a downregulation of SOD2 in the shisha-exposed cells indicating its probable role in increased oxidative stress and tumor initiation.
Immune dysfunction has been observed as an early event associated with carcinogenesis and tumor progression (Staveley-O'Carroll et al., 1998). Cigarette smoke has been revealed to have immunosuppressive effects in smokers (Stampfli and Anderson, 2009). However, no studies indicate a similar effect in shisha users. In this study, bioinformatic analysis of the proteins dysregulated in OKF6/TERT1-Shisha-8 month revealed a similar decrease in immune process. Among the proteins involved in immune response, HLA-A, HLA-B, HLA-C, HLA-F, B2M, OAS1, OAS2, OAS3, BST2, SAMHD1, MX2, ISG15, and IFIT3 were found to be downregulated in the shisha-exposed cells.
Loss or downregulation of major histocompatibility class I molecules, including HLA-A, HLA-B, HLA-C, HLA-F, and B2M, has been identified as one of the chief mechanisms of tumor immune escape from T lymphocytes in several cancers (Garrido et al., 2016). HLA-A has been put forward as a prognostic marker in tonsillar and the base of the tongue cancers (Tertipis et al., 2014). SAMHD1 is a deoxynucleotide triphosphate (dNTP) hydrolase, which is involved in the regulation of immune response and homeostatic balance of cellular dNTPs (Mauney and Hollis, 2018).
Decreased expression or mutation of SAMHD1 increases cell proliferation in lung cancer (Wang et al., 2014), T cell lymphomas (Kodigepalli et al., 2017), colon cancer (Rentoft et al., 2016), and chronic lymphocytic leukemia (Clifford et al., 2014). OAS3 is an interferon stimulated gene (ISG) that is involved in innate immunity and has been reported to be downregulated in bladder carcinoma (Xu et al., 2015) and hepatocellular carcinoma (Narkwa et al., 2017).
Interferon signaling is critical to lymphocyte function. Impaired interferon signaling is an early event responsible for immune dysfunction in patients with cancers, including breast cancer, gastrointestinal cancers, and melanoma (Critchley-Thorne et al., 2009). Biological pathway analysis of the dysregulated proteins in this study also revealed a decrease in interferon pathway in chronic exposure to shisha. Our data indicate a decrease in immune response in oral keratinocytes on chronic exposure to shisha, which in turn may be as a result of decreased interferon signaling. Additional experiments for deciphering the exact mechanism of immune disorder in response to shisha exposure are beyond the scope of this study. Taken together, our study indicates the distinct dysregulation of pathways associated with cell growth and maintenance, invasion, immune response, and interferon signaling.
The use of shisha is no longer endemic to the Middle Eastern countries. The shifting epidemiological trend from the older to the younger generation water pipe users, social acceptability, as well as low perceived risk of shisha smoking, prompt the need for appropriate health promotion response (Ali and Jawad, 2017). Currently, there is a lack of evidence-based literature on the ill effects of shisha use to promote a public health response. Further studies are required to determine the full health burden of shisha smoking. Our study highlights of the deleterious effect of shisha on oral keratinocytes. This study and those of others may help researchers and public health promotion regulatory bodies to take much needed actions toward well-framed public health awareness measures and policies.
Conclusions
This study offers new molecular insights on the ways in which chronic shisha exposure impacts on normal oral keratinocytes, leading to cellular transformation through the dysregulation of the proteome. The temporal data obtained in this study may serve as a resource for identifying potential diagnostic and prognostic markers, as well as therapeutic targets among shisha smokers who might be susceptible to oral cancer. However, the data obtained in this study are based on a chronic cell line model and will require further validation in animal models and clinical settings to establish the role of these dysregulated molecules as potential biomarkers in shisha smoking-related oral cancer. Our study and those of others contribute to a growing evidence base on the ill effects of shisha smoking and may contribute toward public health awareness on negative effects of shisha smoking.
Footnotes
Acknowledgments
The authors thank the Department of Biotechnology (DBT), Government of India for research support to the Institute of Bioinformatics. Institute of Bioinformatics (IOB) is supported by DBT Program Support on Neuroproteomics and infrastructure for proteomic data analysis (BT/01/COE/08/05). The authors thank the “Infosys Foundation” for the research support to the Institute of Bioinformatics. N.B., J.A., and K.P. are the recipients of Senior Research Fellowship from the Council for Scientific and Industrial Research (CSIR), Government of India. The authors thank Dr. Anita Mahadevan of National Institute of Mental Health and Neuro Sciences for providing the use of the microscope facility.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.