
Abstract
Enterococcus faecium is a lactic acid bacterium with applications in food engineering and nutrigenomics, including as starter cultures in fermented foods. To differentiate the E. faecium probiotic from pathogenic bacteria, physiological analyses are often used but they do not guarantee that a bacterial strain is not pathogenic. We report here new findings and an approach based on comparison of the genetic mobility of (1) probiotic, (2) pathogenic, and (3) nonpathogenic and non-probiotic strains, so as to differentiate probiotics, and inform their safe use. The region of the 16S ribosomal DNA (rDNA) genes of different E. faecium strains native to Pernambuco-Brazil was used with the GenBank query sequence. Complete genomes were selected and divided into three groups as noted above to identify the mobile genetic elements (MGEs) (transposase, integrase, conjugative transposon protein and phage) and antibiotic resistance genes (ARGs), and to undertake pan-genome analysis and multiple genome alignment. Differences in the number of MGEs were found in ARGs, in the presence and absence of the genes that differentiate E. faecium probiotics and pathogenic bacteria genetically. Our data suggest that genetic mobility appears to be informative in differentiating between probiotic and pathogenic strains. While the present findings are not necessarily applicable to all probiotics, they offer novel molecular insights to guide future research in nutrigenomics, clinical medicine, and food engineering on new ways to differentiate pathogenic from probiotic bacteria.
Introduction
Enterococcus faecium is a lactic acid bacterium that is valued when used as starter cultures in fermented foods. These bacteria contribute to the development of organoleptic characteristics and the biopreservation of different foods such as cheese, since they are used to produce natural aromatic and antimicrobial compounds, the bacteriocins, which control the growth of pathogens (Giraffa, 2003). There are also strains of E. faecium with probiotic characteristics, which benefit human and animal health (Bybee et al., 2011).
Probiotics are described as: “live microorganisms that, when administered in adequate quantities, confer benefits on the health of the host” (FAO, 2002). These benefits concern physical health such as treating diarrhea, regulating intestinal microbiota, and using them as a therapeutic adjuvant in the treatment of common human disorders (Ejtahed et al., 2011; Villageliú and Lyte, 2018).
The probiotic potential of E. faecium was verified by Ghatargi et al. (2018) and Bagci et al. (2019). However, for this species, the classification as probiotic must be analyzed for each potential strain, as Enterococcus is deemed as not Generally Recognized as Safe (GRAS) and is not recommended in many countries because of the possibility of it causing nosocomial infections. In addition, there is a risk of horizontal transfer of antibiotic resistance genes (ARGs) to other bacteria, and therefore, the probiotic strain could be a reservoir of these genes (Araújo et al., 2013).
Caution still needs to be taken before using members of the genus Enterococcus as a probiotic, since pathogenic vancomycin-resistant E. faecium strains permeate hospital environments. So what would be the way to differentiate them? The answer may lie in their genetic and different evolutionary mechanisms.
Therefore, a study of genetic mobility is a good way to differentiate strains. Mobile genetic elements (MGEs) are a type of genetic material that can move around within a genome. They can also rearrange genes, cause duplication in the host genome, and lead to mutations that are the basis of the evolution of species (Singh et al., 2014). Specifically for E. faecium, the MGEs could be one of the factors responsible for the contrasting probiotic and pathogenic phenotypes, with the transfer of ARGs and exclusive genes to each group, in addition to rearranging and inserting deactivating genes. Enterococcus has different types of conjugation, which facilitates the spread of resistance genes to other species of microorganisms for plasmids that can replicate in several gram-positive bacteria, and the mechanism via transposon conjugation, which can be transferred intracellularly and integrate into the genome of the host bacteria (Zarilli et al., 2005).
This study set out to compare the genetic mobility of probiotic, pathogenic, and “nonpathogenic and nonprobiotic” strains. The starting point of the research was to use sequences of the 16S rDNA of E. faecium native to Pernambuco-Brazil to investigate how best to differentiate between strains with a view to making probiotics safer to use.
Materials and Methods
Characterization, DNA extraction, and amplification of wild “Coalho” cheese isolate
Native strains of lactic acid bacteria isolated from artisanal “Coalho” cheese from Pernambuco State, Brazil, were isolated and kept under refrigeration at −80°C. They were subsequently thawed and grown in Man, Rogosa, and Sharpe—MRS broth (Becton, Dickinson, and Company, Sparks, USA) for 18 h. They were characterized in vitro for probiotic potential. DNA extraction and polymerase chain reaction analysis were performed according to Doumith et al. (2004).
Sequencing of the 16S rDNA region
The isolates of the 16S rDNA region, previously named EF141V and EF137V from Brazil, were sequenced in the University of Minho—Portugal. DNA Sanger sequencing (sense and antisense) of the amplified isolates was used to generate the consensus sequence. The products sequenced used queries that were input into the Basic Local Alignment Search Tool for nucleotide (BLASTn)/National Center for Biotechnology Information (NCBI) database.
Complete genomes
The 16S rDNA sequences of the native strains EF141V (GenBank: MT002681) and EF137V (GenBank: MT002682) were submitted to Blast/NCBI, to select complete genomes. Details regarding the identification and source of the samples used are in Table 4. RAST ANOTATION SERVER with the genus Enterococcus was used to verify the annotations of the genome. Thus, the strains were divided into three groups: probiotic, pathogenic, and “nonpathogenic and nonprobiotic” based on modifications of the methodology of Ghattargi et al. (2018). The classification of the strains as probiotic, nonpathogenic, and pathogenic was based on experimental data in published articles and also on the isolation source of the strain deposited in the GenBank.
Mobile genetic elements
The number of phages was quantified and those in common in probiotic and pathogenic strains in the complete phages were listed. PHAGE Search Tool Enhanced Release (PHASTER) was used under the following references: intact score >90, questionable score 70–90, and score <70 incomplete. The number of MGEs (the protein of transposases, integrases, and conjugative transposons) for both chromosomes and plasmid was quantified in a complete genome of E. faecium obtained from the GenBank.
Antibiotic resistance genes
The Comprehensive Antibiotic Resistance Database was used to predict the resistome under BLAST and RGI (under the criteria of Perfect hit, rigorous hit alone, and perfect and strict hit criteria). CARD was used to verify the position of the gene in the genome (chromosome or plasmid). The ResFinder 3.0 server was used to identify acquired antimicrobial resistance genes in the probiotic bacteria with selected % ID threshold: 90% and selected minimum length: 60%.
Pan-genome
PGAweb-Pan Genomes Analyze Server was used to identify core, accessory, and unique protein families. panX, Pan-genome analysis & exploration, was also used to search for the presence or absence of genes, and phylogenetic inference in 53 strains of E. faecium in the database, using the same criteria as the analysis of complete genomes.
Multiple genome alignment
Mauve, under DNASTAR, was used to perform the synteny analyses and the multiple genome alignments (default setting). The rearrangements were also analyzed.
Ethics approval and consent to participate
No ethics committee approval was required as per study contents and institutional regulations. The research did not involve human subjects or animals, or their biological samples in the study, but only DNA sequenced from bacteria and sequences made available for free on the GenBank.
Results
Complete genomes
The return of the 16S rDNA nucleotide sequences (Table 4), which were input into BLASTn as query sequences showed 11 homologous complete genomes of the three groups under study.
Mobile genetic elements
The probiotic strains compared with other strains analyzed presented a smaller number of transposase and integrase (Fig. 1). Furthermore, two pathogenic strains (Aus0004 and DO) had genes for encoding conjugative transposon proteins in the genome. In addition, the pathogenic RBWHI and 2014-VREF-268 strains presented a higher number of MGEs (Fig. 1). All strains analyzed presented phage in the genome, and 10 of 11 presented intact phages. However, infection by phage is different in probiotic and pathogenic strains. The probiotic strain was infected by Lactobacillus, Lactococcus, and Enterococcus phages, and the pathogenic strains were infected by Listeria, Bacillus, and Staphylococcus phages (Fig. 2).
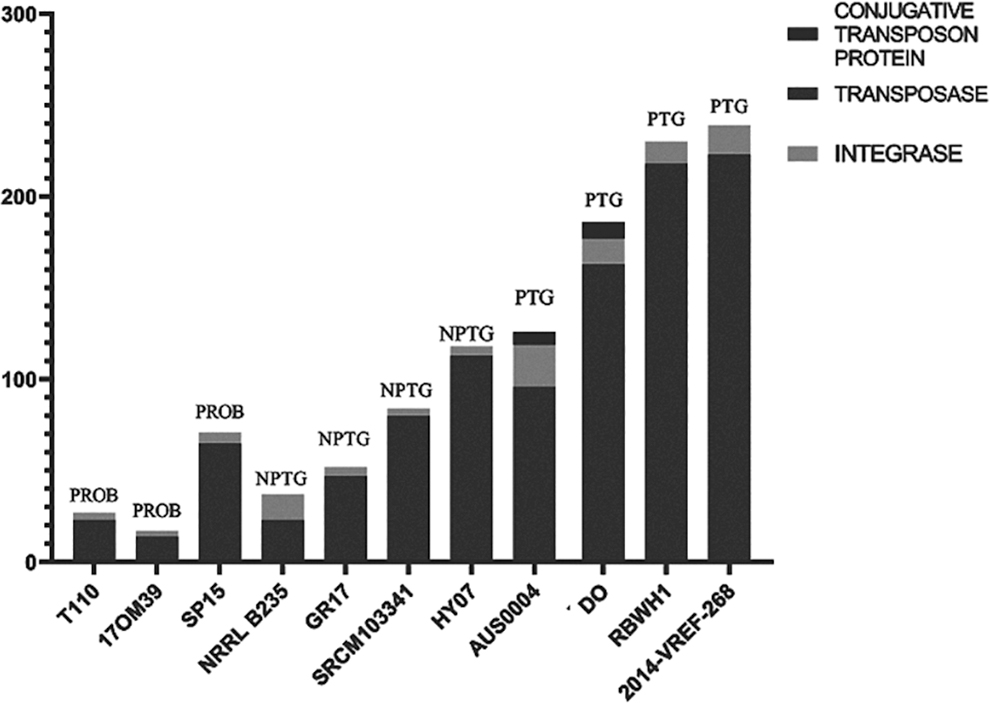
The presence of mobile genetic elements on chromosome and plasmids in strains of Enterococcus faecium. NPTG, nonpathogenic; PROB, probiotic; PTG, pathogenic.
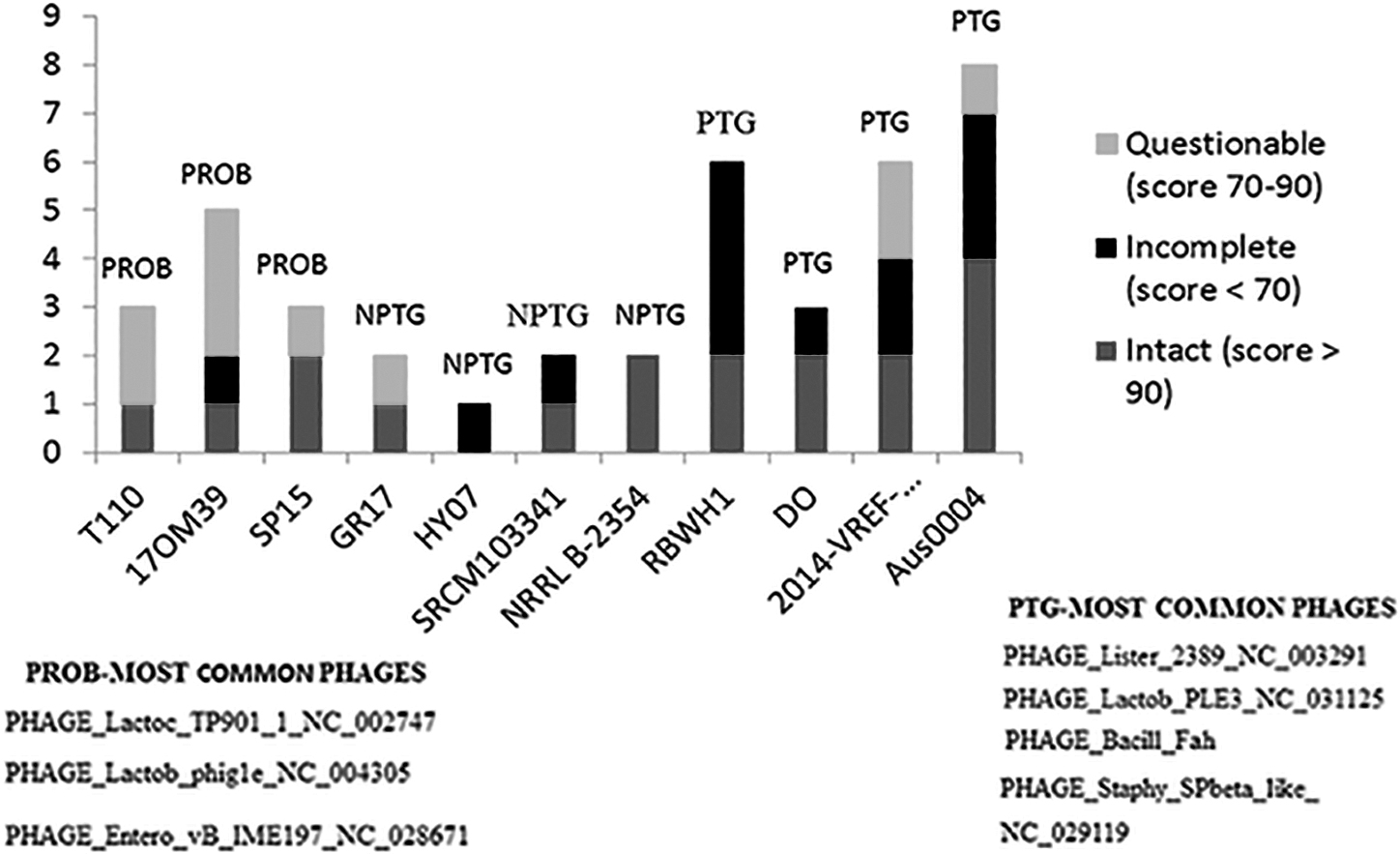
The presence of phages in strains of E. faecium.
Antibiotic resistance genes
All strains in the studies presented the gene AAC (6′)-II for antibiotic protection. The three probiotic strains also presented eatAv, a mutated form of the wild-type eatA ABC-F subfamily protein isolated from E. faecium, which confers resistance to lincosamides, streptogramins A, and pleuromutilins (LSaP phenotype). However, this gene was not present in the pathogenic strains. The efMA gene was present in 17OM39 probiotic, NRRL B-2354 nonpathogenic, and all pathogenic. The pathogenic strains RBW1 and AUS0004 presented genes of vancomycin resistance (vanRB, vanyB, vanHB, vanB, vanXB, and vanSB), and the strains RBW1 and 2014-VREF-238 presented the vanA of vancomycin resistance. The pathogenic strains RBW1, AUS0004, and 2014-VREF-238 also present tet(w/v/w) and for tetracycline resistance antibiotics (Tables 1 and 2). Most ARGs were present in the chromosome and not in the plasmid.
Antibiotic Resistance Genes in Enterococcus faecium Plasmid
● Presence; ◯ Absence.
NPTG, nonpathogenic; PROB, probiotic; PTG, pathogenic.
Antibiotic Resistance Genes in Enterococcus faecium Chromosome
● Presence; ◯ Absence.
Pan-genome
The genome of E. faecium has the largest accessory genome (data no shown). Thus, probiotic, pathogenic, and nonpathogenic bacteria presented unique genes to the group (Table 3). All strains analyzed in the PANX WebServer (53 strains) presented the gene of Bacteriocin production. Only, the probiotic and nonpathogenic strain presented the gene enterocin-P, for enterocin exportation. The probiotic strain also presented the fibronectin type III domain-containing protein and the ECF-type riboflavin transporter substrate-binding protein. The nonpathogenic strain presented the Class IIb bacteriocin, lactobin A/cerein 7B family. However, the pathogenic strain presented the conjugative transposon protein, thermostable direct hemolysin 2, and botulinum neurotoxin type C1.
Proteins Found in Specific Groups of Enterococcus faecium
The Strains of Enterococcus faecium with Complete Genomes Analyzed and Registered in GenBank
Multiple genome alignment
The alignment of multiple genomes in the Mauve software showed rearrangements in a conserved region (Figs. 3 and 4). There was variation in the size of the genetic clusters.
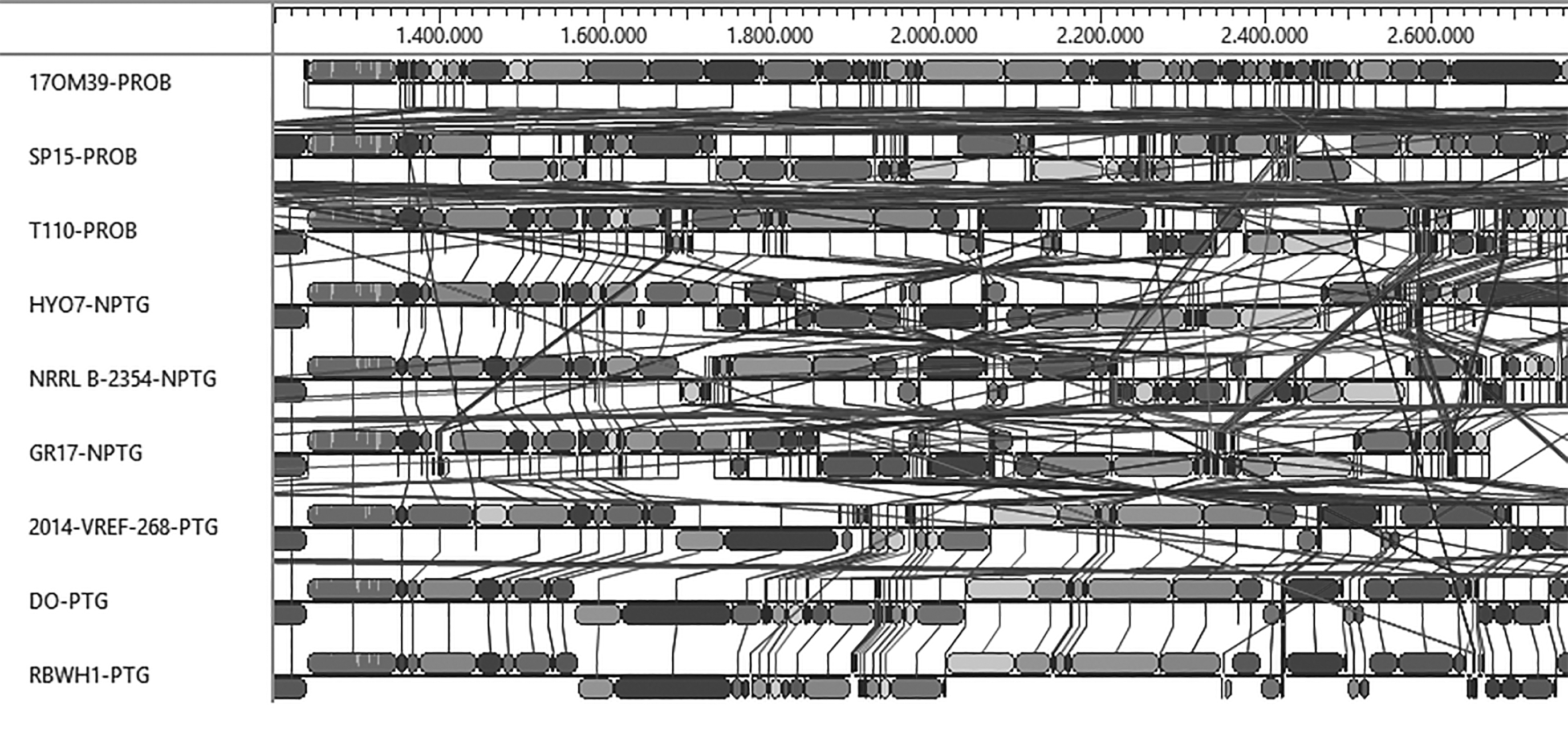
Synteny of E. faecium strains.

Synteny of E. faecium strains (overview).
Discussion
Differentiation of probiotic and pathogenic bacteria is an active area of research in nutrigenomics and food engineering fields in particular. Food safety can benefit from multiomics research as well (Bayram and Gökırmaklı, 2018). We found in the present study that the pathogenic strains presented a higher number of MGEs both in the chromosome and in the plasmid, and a low probiotic element (Fig. 1), as also observed by Zhong et al. (2019).
We note that the number of transposases in the genomes of probiotic strains should be continuously analyzed. As defined by Bourque et al. (2018) in their work on transposable elements, these enzymes favor genomic rearrangements and gene duplication by incorporating adjacent genes during the transposition or by genetic recombination because they are present in multiple copies and sometimes they create or reverse mutations.
Some transposable elements also contain integrons, genetic elements that can capture and express genes from other sources. These contain integrase, which can integrate gene cassettes (Domingues et al., 2012). E. faecium with a large number of transposases and integrases should be recommended with caution in probiotics or in fermented foods, as safety and efficacy are not guaranteed due to the plasticity of the genome.
Due to the accumulation of MGEs, according to Palmer et al. (2010), multidrug-resistant enterococci differ from nonpathogenic strains because, among other reasons, their genomes are frequently 25% larger than nonpathogenic strains. Zhong et al. (2019) showed that the E. faecium genome isolated from humans was larger than the isolates from dairy products. However, in our findings, despite the greater number of transposases in pathogenic E. faecium, the size of the genome remains constant compared with nonpathogenic strains, which may indicate the loss of genes related to probiotic characteristics in the pathogenic strains.
This should also be considered in the classification of probiotic E. faecium. The difference in phage infection between probiotic and pathogenic bacteria could explain the acquisition of genes, as probiotic bacteria are infected by a phage of a GRAS bacterium such as Lactobacillus sp. and pathogenic bacteria by pathogenic bacterial phages such as Listeria sp. Bacteriophages can package part of their host's genetic material, including ARGs, contributing to a rapid dissemination of resistances among bacteria. Antibiotic resistance is a major threat to public health and food security (Gómez et al., 2019). Therefore, phages in probiotic bacteria should be monitored.
The ARGs in probiotic E. faecium are present. However, for example, the chromosomal aac(6′)-II, encoding a 6′-N-aminoglycoside acetyltransferase, is gene specific for E. faecium. Costa et al. (1993) suggest that the aac(6′)-II gene is species specific and may be used to identify E. faecium, as with the eatAv, a mutated form of the wild-type eatA ABC-F subfamily protein. The ABC-F subfamily of ATP-binding cassette proteins mediates resistance to a broad array of clinically important antibiotic classes that target the ribosome of gram-positive pathogens (Sharkey et al., 2016).
The genes vanA and vanB were localized in the chromosome and/or plasmid of pathogenic strains (Tables 1 and 2). The vanA phenotype is characterized by the strains that exhibit high levels of resistance to vancomycin and teicoplanin (Arthur et al., 1992). The vanB operon induces several levels of vancomycin resistance but does not induce resistance to teicoplanin. This gene is located on the bacterial chromosome, but can be obtained from the plasmids of other organisms (Leme and Ferreira 2001). There were also the genes of tetracycline resistance (tet(w/v/w) and tetM) and genes of erythromycin resistance (ermA, ermB, and ermG) in pathogenic strains.
The analysis of the ARG in probiotic and food bacteria must be performed more than once, because depending on the niche, “good” bacteria can acquire ARGs. The continuous exposure to antibiotics may cause a mutation that confers bacteria resistance to certain antimicrobial compounds, enabling their survival, primarily in isolation from a hospital environment (Shepard and Gilmore 2002). Plasmids must be analyzed because of the transfer of ARGs. However, resistance genes are also present in the chromosome and can “jump.”
Pan-genomic analysis showed (Table 3) that probiotic and pathogenic bacteria have unique genes for their niche. Probiotic and alimentary E. faecium has genes for the production of bacteriocins, and the bacteriocin is active against selected strains of Listeria, Staphylococcus, Pediococcus, and Enterococcus (Abengózar et al., 2017), B9 vitamin, and B12 vitamin, which causes health and food benefits. Probiotic bacteria, members of the gut microbiota, can synthesize vitamin K and most of the water soluble B vitamins in humans (Hill, 1997). The probiotic has the fibronectin adhesion protein, which is responsible for adhesion to the intestinal epithelia. This protein is needed because to be considered a probiotic, the bacteria must temporarily adhere to the intestine (Monteagudo-Mera et al., 2019).
However, the pathogenic bacteria have conjugative transposons for horizontal gene transfer, mainly of antibiotic resistance and virulence (Johnson and Grossman, 2015). The gene botulinum neurotoxin type C1 was found in the genome of pathogenic E. faecium and Zhang et al. (2018) in their study showed that E. faecium carried out a horizontal gene transfer with Clostridium botulinum, which is representative of the potential of genes to exchange E. faecium with other pathogenic bacteria, this being a major problem for the health of the population. Thermostable direct hemolysin 2 (TRH) is related to the process of infection and lysis of blood cells in the host, which is highly cytotoxic to human gastrointestinal cells Vibrio parahaemolyticus (Raghunath, 2015).
E. faecium genes are in constant motion. Sometimes they are located in the chromosome and sometimes in the plasmid (Van Schaik et al., 2010). Rearrangements were found in the genomes of the three groups studied. However, there was no pattern of rearrangement to classify the bacteria as probiotic, pathogenic, and nonpathogenic in the genome alignment by Mauve, considering only this region, shown in Figure 3.
The difference is noticed when considering the complete genome (Fig. 4), mainly in pathogenic bacteria, which formed a characteristic pattern. However, the presence and absence of certain genes enabled probiotic, pathogenic, and nonpathogenic bacteria to be grouped.
Conclusions
Genetic mobility appears informative in differentiating among probiotic, pathogenic, and “nonprobiotic and nonpathogenic” strains and can be used to complement studies on probiotic bacteria and to verify pathogenicity. However, it is recommended that further studies be undertaken because a limited number of articles have been published to date and the number of probiotic strains available in the current database is small. Probiotic strains need to be kept under constant observation because of their mobility, exclusion, and/or duplication of genes. These can interfere with the product quality in food engineering, quality of the probiotic, in addition to the health risk of ARGs. E. faecium pathogenic and probiotic strains tend to follow different evolutionary mechanisms and so because of their duality, they may or may not be good for the host. While the present findings are not necessarily applicable to all probiotics, they offer novel molecular insights to guide future research in integrative biology, nutrigenomics, clinical medicine, and food engineering on new ways to differentiate pathogenic from probiotic bacteria.
Footnotes
Acknowledgments
The authors are grateful to the Capes (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasília—Brazil) and Cenapesq (research support center) and Biochemistry and Sequencing Laboratory Tânia Falcão-Genoma, both by University Federal Rural of Pernambuco—Brazil, for the infrastructure assistance.
Author Disclosure Statement
The authors declare they have no competing financial interests.
Funding Information
No funding was received in support of the present study.