
Abstract
Early cell biology reports demonstrated the presence of cells with stem-like properties in bone marrow, with both hematopoietic and mesenchymal lineages. Over the years, various investigations have purified and characterized mesenchymal stromal/stem cells (MSCs) from different human tissues as cells with multilineage differentiation potential under the appropriate conditions. Due to their appealing characteristics and versatile potentials, MSCs are leveraged in many applications in medicine such as oncology, bioprinting, and as recent as therapeutics discovery and innovation for COVID-19. To date, studies indicate that MSCs have varied differentiation capabilities into different cell types, and demonstrate immunomodulating and anti-inflammatory properties. Different microenvironments or niche for MSCs and their resulting heterogeneity may influence attendant cellular behavior and differentiation capacity. The potential clinical applications of MSCs and exosomes derived from these cells have led to an avalanche of research reports on their properties and hundreds of clinical trials being undertaken. There is ample reason to think, as discussed in this expert review that the future looks bright and promising for MSC research, with many clinical trials under way to ascertain their clinical utility. This review provides a synthesis of the latest advances and trends in MSC research to allow for broad and critically informed use of MSCs. Early observations of the presence of these cells in the bone marrow and their remarkable differentiation capabilities and immunomodulation are also presented.
Introduction
Mesenchymal stem/stromal cells: what is in a name?
Early reports indicated that the bone marrow contained both hematopoietic stem cells as well as mesenchymal stromal/stem cells (MSCs) (Friedenstein et al., 1968, 1970). Classic transplantation experiments by Friedensten et al. (1970) demonstrated that cells from the bone marrow can create a bone marrow microenvironment in the kidney as well as formation of bone tissue. This indicated that besides hematopoietic stem cells, there was an additional stem cell type, later identified as MSCs, a term coined by Caplan in the 1990s (Fig. 1) (Caplan, 1991; Friedenstein et al., 1968, 1970, 1974). MSCs have the ability to grow in vitro, display enhanced differentiation capabilities to form the connective tissues present in different organs compared with other cells, and release large amounts of biomolecules such as growth factors and cytokines (Caplan, 1991; Senthebane et al., 2017).

Historical background of mesenchymal stem cells. Cells isolated from the bone marrow were shown to form clonogenic colonies. Studies later showed that cells from the bone marrow can differentiate into the connective tissue.
MSCs that have been called by various names and experiments have shown that these cells demonstrate different degrees of stemness and differentiation capacities, capacities that appear to diminish with age (Bianco et al., 2008; De Schauwer et al., 2011; Pal and Das, 2017). In addition to mesenchymal stem cells, these cells can be referred to as mesenchymal stromal cells as well as multipotent stromal cells among many names (Dzobo et al., 2016a, 2016b, 2018). Recent efforts have been made to name MSCs based on their mechanism of action, which is mainly secretion of various biological molecules, including growth factors and cytokines (Caplan, 2017a; Sacchetti et al., 2016; Sipp et al., 2018). In this regard, MSCs can be seen as cells involved in influencing cellular signaling (Caplan, 2017a; Sipp et al., 2018).
It is important to note that MSCs differ in their origins, differentiation ability, functions, and consequently their therapeutic uses. As early as 2006, the International Society for Cellular Therapy (ISCT) defined MSCs as multipotent mesenchymal stromal cells and published a set of conditions needed to be fulfilled for cells to be called such. These conditions included the expression of several surface antigens such as cluster of differentiation (CD)90, CD105, as well as being able to grow in vitro as substrate-adhering cells (Dominici et al., 2006; Dzobo et al., 2016a; Horwitz et al., 2005).
As we demonstrated in our earlier report in addition to other studies in the literature, MSCs can differentiate into adipocytes, chondroblasts, and osteoblasts as well as lacking CD14, CD34, and CD45 expression (Dominici et al., 2006; Dzobo et al., 2016a). Additional markers also proved their utility in the isolation and identification of MSCs. For example, CD106, CD146, and stromal-1 antigen (STRO-1) are additional markers used to identify MSCs in vitro and are expressed by cells with differentiation abilities into multilineages (Gronthos et al., 2003; Sacchetti et al., 2007; Simmons and Torok-Storb, 1991).
The term “mesenchymal” present in many of the names given to MSCs comes from the word “mesenchyme,” which is used to describe cells from the mesoderm of a developing embryo. These cells migrate and are distributed throughout the body of the developing embryo (Caplan, 1991). In adults, the mesenchymal cells form the connective tissue, including cartilage, muscle, and bone marrow (Muschler and Midura, 2002). These cells are characterized based on their expression of markers, such as fibronectin, laminin β1, and vimentin (Dennis and Charbord, 2002; Pittenger et al., 1999). It is important to note that the origin of MSCs has not been proven since their discovery. Several reports have shown that MSCs can also be derived from cells of the ectoderm (Andrzejewska et al., 2019; Isern et al., 2014; Morikawa et al., 2009; Takashima et al., 2007).
Different MSC cells have been identified based on expression of markers such as Nestin as well as cell proliferation in bone marrows of developing embryos, with some cells being derived from the mesoderm, whereas others are derived from the neural crest (Isern et al., 2014). Thus, the origin of MSCs is broad based and is linked to cells of different germ layers. In summary, the germ layers from which MSCs are derived determine their final function in the adult body.
Sources and Niche of Adult MSCs
Well-known sources of MSCs include the bone marrow, umbilical cord tissue, adipose tissue as well the placenta (Fig. 2) (Haynesworth et al., 1992). In most cases, some of these tissues do not raise ethical issues regarding harvesting of cells as they can be considered medical waste for example during liposuction and after childbirth (He et al., 2017; In ‘T Anker et al., 2004; Romanov et al., 2003; Zuk et al., 2001). Several factors such as the final use of the isolated MSCs, the differentiation potential of the MSCs, and the total number of cells needed for the application can influence the source of the cells (Beauséjour, 2007; Farkhad et al., 2021; Stolzing et al., 2008).
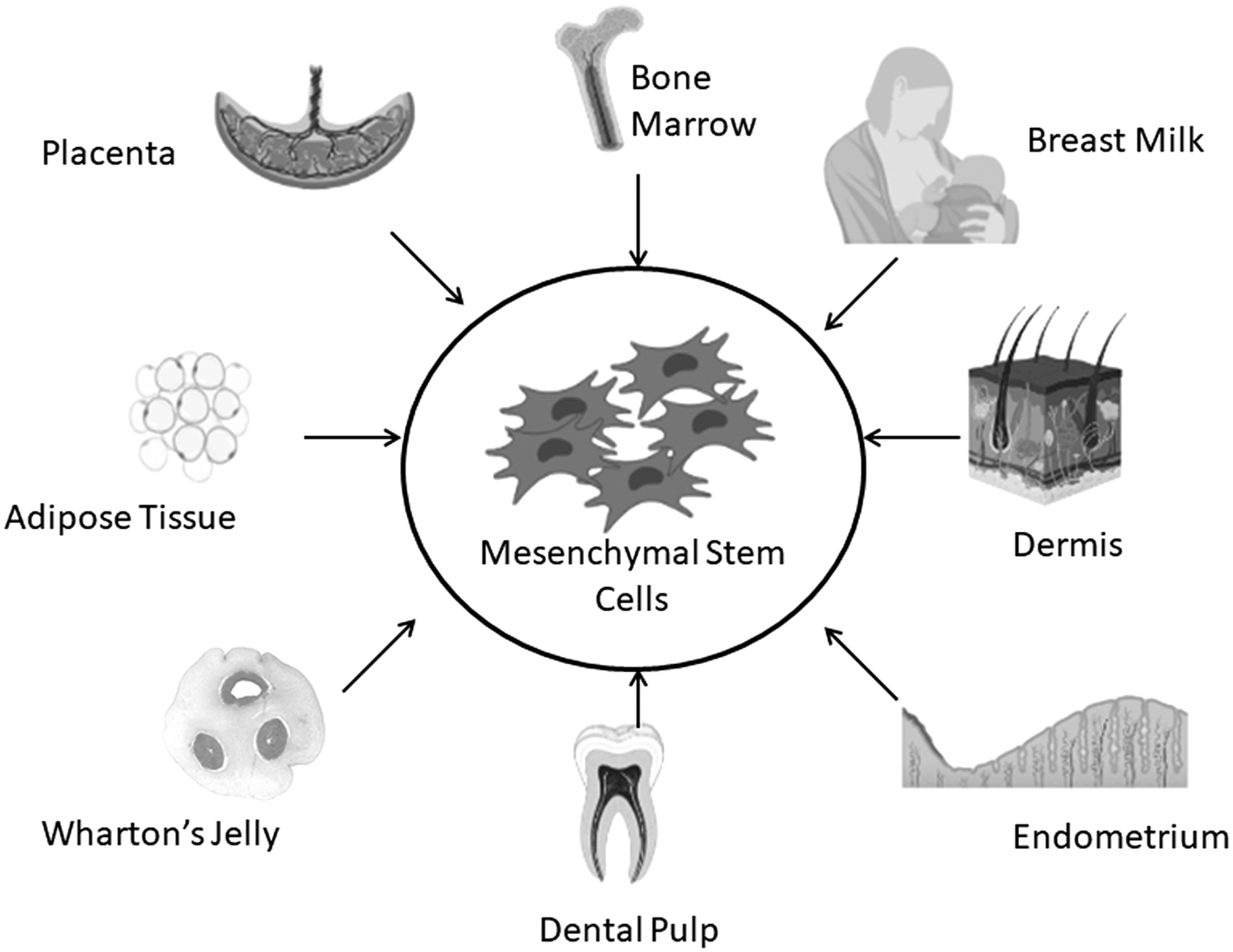
Sources of mesenchymal stem cells.
MSCs niche is the microenvironment within which MSCs exist and can maintain their “stem cell” status (Schofield, 1978; Zhang et al., 2003). In the bone marrow, MSCs released growth factors and cytokines required for the proliferation and maturation of hematopoietic cells (Anthony and Link, 2014; Méndez-Ferrer et al., 2010). Méndez-Ferrer et al. (2010) demonstrated using MSCs expressing Nestin that they are part of the hematopoietic niche in the bone marrow. In their study, the authors showed that MSCs expressing Nestin can form colonies with fibroblastic activity and can grow in suspension with self-renewing and expansion capabilities (Méndez-Ferrer et al., 2010). Several other studies have also shown that MSCs can be isolated from tissues derived from the mesoderm during embryonic development (Isern et al., 2014; Slukvin and Vodyanik, 2011; Vodyanik et al., 2010).
Several other studies have demonstrated that MSCs can be derived from different sources and therefore can exist in different niche. For example, it was shown that cells with MSC-like properties can be isolated from perivascular space of blood vessels from many tissues, including pancreas and adipose tissue (Chen et al., 2009; Crisan et al., 2008; Da Silva Meirelles et al., 2008; Pittenger et al., 1999). The isolated cells were able to express markers, such as CD146, PDGF-Rβ, as well as alkaline phosphatase (ALP) (Crisan et al., 2008). The osteoprogenitor cells or MSCs within sinusoidal blood vessels are able to produce a hematopoietic microenvironment after transplantation (Ding et al., 2012; Sacchetti et al., 2007). However, several other studies, including one by Feng et al. (2011) demonstrated that cells that do not express CD146, but can differentiate into pericytes for example, are able to meet the minimum criteria for MSCs (Covas et al., 2008; Da Silva Meirelles et al., 2015).
Several studies have shown that MSCs and pericytes are functionally different although they express similar genes and have almost identical differentiation capabilities (Blocki et al., 2013; Caplan, 2008; Da Silva Meirelles et al., 2008; Díaz-Flores et al., 2009; Farrington-Rock et al., 2004). Only pericytes are able to form fully functional blood vessels in contrast to MSCs isolated from the bone marrow (Blocki et al., 2013; Caplan, 2017b; De Souza et al., 2016; Guimarães-Camboa et al., 2017).
Further studies are needed to fully explain and delineate the relationship between MSCs and pericytes. In vitro assays performed by Blocki et al. (2013) demonstrated that not all MSCs are pericytes. A study by Loibl et al. (2014) demonstrated that MSCs can be induced into a pericyte-like phenotype through coculture with endothelial progenitor cells. In addition, flow cytometric purification of pericytes can result in pericytes that are similar to MSCs in many growth, morphology, and differentiation characteristics (Harrell et al., 2018; Kostallari et al., 2015; Mao et al., 2017; Rozycka et al., 2017; Wong et al., 2015). Overall, the interconnectivity between MSCs and pericytes require further investigations.
Cell–cell interactions are important within MSCs' in vivo niche for the maintenance of stem cell properties, and cadherins such as N-cadherin have been shown to play key roles (Abdal Dayem et al., 2018; Votteler et al., 2010). Migration of MSCs from their niche results in new environments characterized by increased extracellular matrix (ECM) molecules, including collagens and proteoglycans (Castillo and Jacobs, 2010; Cosgrove et al., 2016; Tatullo et al., 2016). To be able to interact with ECM molecules, MSCs express integrins and are able to form focal adhesions (Steward and Kelly, 2015; Wang and Chen, 2013). Culture of MSCs in vitro induce cellular changes and influence differentiation into specific lineages.
For example, culture on rigid surfaces can induce osteogenic and adipogenic differentiation (Dzobo et al., 2016a; Engler et al., 2006; Pek et al., 2010). Soft tissues and surfaces induce MSCs toward the myogenic and neurogenic lineages (Kao et al., 2021; Su et al., 2021; Sun et al., 2020; Zonderland and Moroni, 2021). Thus, the stiffness of a surface or microenvironment can influence the expression of markers and shape of MSCs both in vivo and in vitro (Fig. 3) (Dupont et al., 2011; Fu et al., 2010; Yang et al., 2014). Generally, a stiff surface or environment tends to favor MSC differentiation into lineages associated with stiff tissues, such as bones (Engler et al., 2006; Fu et al., 2010).

Mesenchymal stem cells interact with different components of its microenvironment, including cells, extracellular matrix, and biomolecules.
Heterogeneity of Mesenchymal Stem Cells
MSCs display great heterogeneity in terms of their functionality and consequently application despite their sharing of several characteristics. Thus, while MSCs from different tissues meet the minimum criteria needed for classification as MSCs, those isolated from different tissues and developmental stages, display differences that impact on their use (Liu et al., 2015; Muhammad et al., 2015; Sensebe et al., 2010). Initially isolated from the bone marrow, MSCs can now be obtained from a variety of sources such as Wharton's Jelly, adipose, blood, placenta, and amniotic fluid (Fig. 2) (Agha-Hosseini et al., 2010; Chen et al., 2011; Kassis et al., 2006; Katz et al., 2005; Meng et al., 2007). MSCs from fetal tissues tend to proliferate at a higher rate and have longer telomeres than those from adult tissues (Guillot et al., 2007; O'Donoghue and Fisk, 2004).
In addition, several reports also indicate that fetal MSCs have a higher differentiation potential than adult tissue-derived MSCs (Christodoulou et al., 2013; Hass et al., 2011; Zhang et al., 2009). Furthermore, MSCs from fetal tissues can proliferate in vitro for a longer period of time before displaying characteristics of senescence (Hass et al., 2011). In contrast, MSCs from the bone marrow and adult adipose tissue display a higher level of stemness as demonstrated by the formation of more colony-forming units compared with fetal tissue-derived MSCs (Fafián-Labora et al., 2015; Ganguly et al., 2019; Mennan et al., 2019; Tawonsawatruk et al., 2012).
In a detailed analysis of MSCs from different sources, Heo et al. (2016) demonstrated that MSCs from bone marrow express DLX5 and Sox2 needed for brain neuron development. In addition, the authors showed that bone marrow- and adipose tissue-derived MSCs displayed similar differentiation capacities as well as gene expression. MSCs from different sources also display multilineage differentiation as well as immunomodulatory behaviors.
Kern et al. (2006) demonstrated that MSCs from the bone marrow and adipose tissue can easily be isolated compared with MSCs from the umbilical cord blood. In addition, the authors also demonstrated that umbilical cord blood MSCs showed no adipogenic differentiation abilities compared with those from the bone marrow and adipose tissue (Kern et al., 2006). Several studies, including our own have demonstrated that indeed, adipose-derived MSCs can undergo multilineage differentiation under different conditions (Dzobo et al., 2016a; Kirsch et al., 2020; Mathot et al., 2021; Voga et al., 2020).
MSCs from different donors and of different ages also display great heterogeneity. Phinney et al. (1999) demonstrated that bone marrow-derived MSCs from different donors show differences in differentiation capabilities as well as osteogenic potentials. The latter study showed that MSCs from different donors show different growth rates and ALP activity in culture (Phinney et al., 1999). In addition, the authors postulated that MSC's heterogeneity and differentiation potential were also influenced by the method of harvest used. For example, Pezzi et al. (2017) demonstrated that oxygen levels impact on MSC heterogeneity in vitro and this has huge influence on long-term culture of MSCs for therapy.
Other reports have postulated that MSCs may be a product of long-term in vitro culture (Lukomska et al., 2019; Okamoto et al., 2002; Pezzi et al., 2017). Furthermore, other reports demonstrate bone marrow-derived MSC heterogeneity due to age of donor, with more MSCs from older donors undergoing apoptosis and having a low proliferation rate than those from younger donors (Gala and Fabian, 2021; Massaro et al., 2020; Zhou et al., 2008). Naive MSCs have been shown to exhibit heterogeneity based on transcription data (Rohart et al., 2016).
If MSCs are to be used for therapy, there may be need to synchronize the cells through stimulation or growth in media with no growth factors (Kabat et al., 2020; Lukomska et al., 2019; Moll et al., 2019). “Priming” of MSCs through the use of a stimulus must be done with caution as some factors can induce undesirable effects such as apoptosis and senescence (Klinker et al., 2017; Le Blanc, 2003; Sivanathan et al., 2014).
Current data show that priming of MSCs can be done through the use of cytokines, drugs, growth factors, biomaterials, and the ECM as well as hypoxia. When cultured in vitro, MSCs display three main morphologies. Colter et al. (2000, 2001) as well as others demonstrated that besides the fibroblastic spindle-shaped cells and the large flat cells, a third type of cells characterized as small, round, and having enhanced self-renewal abilities also exists in vitro (Casciaro et al., 2021; Prockop et al., 2001). Studies now show that cells from a single colony are not all the same and can demonstrate multipotence, dipotence, or unipotence (Okamoto et al., 2002). Thus, a single MSC colony can give rise to different cell types with varied differentiation potentials.
With publication of different data sets, it has been theorized that even within MSC populations, cells exist in a hierarchical structure, with some remaining as unipotential or dipotential, whereas others become multipotential, a characteristic which they can eventually loose (Beyer Nardi and Da Silva Meirelles, 2006; Childs et al., 2020; Koide et al., 2007; Russell et al., 2010). Several lines of evidence suggest that the transformation of MSCs may be linked to a lower proliferation rate and decreased expression of markers such as CD146 (Durand et al., 2006; Muraglia et al., 2000; Zou et al., 2008).
Mesenchymal Stem Cells and the Immune System
Reports over the years have shown that MSCs have immunomodulatory properties. MSCs were shown to influence the behavior of cells involved in immune responses and to be able to impact a lot of cellular processes (Fig. 4) (Krampera et al., 2013). MSCs achieve their immunomodulating effects through prevention of apoptosis in native and activated neutrophils, preventing the action of peripheral blood mononuclear cell proliferation, preventing the recruitment of cells at wound sites, as well as preventing the interaction between neutrophils and vascular endothelial cells (Table 1) (Cassatella et al., 2011; Moll et al., 2011; Munir et al., 2015; Tu et al., 2010).

Interactions between mesenchymal stem cells and immune cells. Through the synthesis and secretion of various factors, including TGF-β, PGE2, interleukin-6, interleukin-10, and Gal-1, mesenchymal stem cells impact on the proliferation and activation of immune cells such as macrophages, dendritic cells, and natural killer cells. TGF-β, transforming growth factor beta.
Immunomodulatory Effects of Mesenchymal Stem Cells
IL, interleukin.
Several reports have shown that native or transformed MSCs release several cytokines, including transforming growth factor beta (TGF-β), which can impact other cells such as tumor cells and neutrophils (Brandau et al., 2014; Senthebane et al., 2017; Yang et al., 2021). Furthermore, MSCs have been shown to release various chemokines involved in recruiting macrophages and neutrophils to wound areas (Chen et al., 2008; Hu et al., 2018; Isakson et al., 2015; Rustad and Gurtner, 2012). Brown et al. (2011) demonstrated that MSCs prevent mast cell degranulation and limit cytokine secretion by these cells (Chehelcheraghi et al., 2020; Kim et al., 2015; Liu et al., 2016). In addition, MSCs can inhibit the proliferation of natural killer cells and have been shown to suppress natural killer cell cytotoxicity (Spaggiari et al., 2006, 2008).
Several MSC-secreted factors have been shown to prevent the differentiation and maturation of blood monocytes into dendritic cells as well as preventing dendritic cells from migrating to tissues (Jiang et al., 2005; Nauta et al., 2006; Su et al., 2011; Zhang et al., 2004). MSCs have been shown to induce transformation of M1 macrophages into M2 macrophages, through NF-κB and STAT3 pathways, with anti-inflammatory properties (Chen et al., 2014; Gao et al., 2014). The resulting M2 macrophages are anti-inflammatory and release cytokines such as interleukin 10 (IL-10) that can prevent T cell proliferation (Chen et al., 2014; Duffy et al., 2011; Soleymaninejadian et al., 2012). Glennie et al. (2005) demonstrated that bone marrow-derived MSCs can inhibit growth of activated T cells and lymphocytes.
Overall, MSCs induce a reduction in synthesis and release of cytokines that promote inflammation and increase the synthesis of anti-inflammatory cytokines by T lymphocytes (Yañez et al., 2006, 2010). MSCs have also been shown to prevent the differentiation of CD4-positive T lymphocytes into T helper cells, while inducing differentiation of T cells into CD4-positive regulatory T cells (Aggarwal and Pittenger, 2005; Ghannam et al., 2010). It has been shown that MSCs suppress chemokine expression by B lymphocytes and this impact the ability of the B lymphocytes to migrate (Corcione et al., 2006; Musso et al., 2011). Finally, MSCs have been shown to inhibit the synthesis of several immunoglobulins by activated B cells and in the process prevents formation of plasma cells (Fan et al., 2016; Jiang and Xu, 2020; Wang et al., 2018).
Paracrine Properties of Mesenchymal Stem Cells
Recent reports show that MSCs secrete several factors that act in both autocrine and paracrine fashion. Thus, one area of MSCs being investigated thoroughly is that of provision of factors rather than the differentiation of MSCs into different lineages. If MSCs can release factors necessary for immune modulation, tissue repair, and wound healing, then MSCs can be useful in various ways. One primary function of secreted factors is the regeneration of damaged or diseased tissues (Dzobo et al., 2018; González-González et al., 2020; Kupcova Skalnikova, 2013; Lee et al., 2011).
Factors secreted by MSCs include growth factors, enzymes, cytokines, chemokines, as well as ECM proteins (Chen et al., 2008; González-González et al., 2020). MSC-secreted factors act in a context-dependent manner. For example, secreted factors such as vascular endothelial growth factor (VEGF) and tissue inhibitor of metalloproteinase (TIMP)-1 and TIMP-2 are known to regulate angiogenesis in opposing ways, with VEGF promoting angiogenesis, whereas TIMP-1 and TIMP-2 inhibiting angiogenesis (Cantinieaux et al., 2013; Zanotti et al., 2016).
Additionally, VEGF secreted by MSCs is known to influence other processes such as fibrosis and apoptosis (Kwon et al., 2014; Pankajakshan and Agrawal, 2014; Shen et al., 2015). MSC-derived VEGF, IGF-1, IGF-2, and hepatocyte growth factor (HGF) have antiapoptotic effects (Harmey and Bouchier-Hayes, 2002; Kaga et al., 2006; Mehmet et al., 2006; Sadat et al., 2007; Yanai et al., 2006). Thus, an increase in the expression of one factor can have multiple effects on the tissue into which it is released. Cantinieaux et al. (2013) demonstrated that conditioned media from bone marrow-derived MSCs can protect neurons from apoptosis as well as activates macrophages. Menezes et al. (2020) demonstrated that MSCs can recruit pericytes and induce angiogenesis through release of factors such as VEGF during the repair of spinal cord injury in rats.
In classic coculture experiments of both esophageal and breast cancer cells with Wharton Jelly-derived MSCs, we demonstrated that MSCs reduced the effects of paclitaxel and cisplatin on cancer cells (Senthebane et al., 2017). One factor that was released in large quantities by both cancer cells and MSCs was TGF-β and was involved in the transformation of MSCs into cancer-associated fibroblasts (Dzobo, 2020; Senthebane et al., 2017). Thus, MSCs protected cancer cells from drug-induced apoptosis. Several other pieces of evidence substantiated our findings (Castells et al., 2012).
For example, Eliopoulos et al. (2010) demonstrated that bone marrow-derived MSCs also reduce the renotoxicity of cisplatin in mice. Bergfeld et al. (2014) showed that MSCs from the bone marrow enhances the metastatic abilities of tumor cells in addition to the promotion of drug resistance. Orimo et al. (2005) also demonstrated that stromal cells such as MSCs promote breast carcinoma cells as well as angiogenesis through the secretion of factors, including SDF-1. It appears that the source of the MSCs and the cancer under study is important in determining the overall effect of MSCs on the cancer cells. For example, Khakoo et al. (2006) demonstrated that MSCs can exert antitumorigenic effects in a model of Kaposi's sarcoma.
In yet another study, Ohlsson et al. (2003) showed that mesenchymal progenitor cells can inhibit tumor growth when grown in a gelatin matrix. Furthermore, Maestroni et al. (1999) demonstrated that factors secreted by bone marrow-derived stromal cells can inhibit the growth of Lewis lung carcinoma and B16 melanoma cells in mice. The contrasting data as presented above demonstrate the importance of accurate reporting and understanding the effects of MSCs and factors they release on cancer cells (Lazennec and Jorgensen, 2008; Vizoso et al., 2017). In addition, one of our studies showed that Wharton's Jelly-derived MSCs when cultured on an ECM activate apoptosis in a p21-dependent mechanism in esophageal and breast cancer cells (Dzobo et al., 2016b). It appears that the effect of MSCs on cancer cells is context dependent, may depend on the paracrine factors released and requires further investigations.
Several MSC-derived factors, including VEGF, HGF, epidermal growth factor (EGF), SDF-1, and TGF-α can suppress fibrosis in many tissues (Álvarez et al., 2015; Eom et al., 2015; Lan et al., 2015; Mahmoudi et al., 2020; Srour and Thébaud, 2015). Chemokines with opposing effects, including stromal cell-derived factor 1, CCL5, and monocyte chemoattractant protein 2 are also secreted by MSCs and impact on cell chemotaxis (Bai et al., 2016; Broxmeyer et al., 1999; Deshmane et al., 2009; Xing et al., 2014).
Some studies have shown that MSC-secreted factors can have antibacterial and antiviral activities (Meisel et al., 2011; Mezey and Nemeth, 2015). Indeed, several cytokines, including IL-6, IL-10, and TNF-α have been shown to fluctuate during bacterial infection (Masalova et al., 2020; Mei et al., 2019; Ou et al., 2016; Schlosser et al., 2019; Vasandan et al., 2016). Meisel et al. (2011) demonstrated that MSCs stimulated by inflammatory cytokines display antimicrobial activities against several bacteria, protozoal parasites, and viruses. In addition, Li et al. (2020) demonstrated that MSCs can prevent inflammation and severity of Haemophilus influenza infection by recruiting CXCR3+ Tregs.
Mesenchymal Stem Cell-Derived Extracellular Vesicles
MSCs have also been suggested to secrete extracellular vesicles (EVs), through which they can relay signals and cues to other cells. These EVs may include apoptotic bodies, microvesicles, and exosomes. The biomolecules and other factors that are carried by EVs, also called cargo, are largely dependent on the cell type from which they originate, although some reports indicate some processing can take place during transportation. EV cargo is composed mainly of lipids, nucleic acids, and proteins and thus mainly functions to regulate cellular processes, cellular functions, immune response, and also the maintenance of homeostasis (Margolis and Sadovsky, 2019; Raposo and Stoorvogel, 2013).
Recent reports indicate that EVs isolated from MSCs can function in the same way as the MSCs (Kahmini and Shahgaldi, 2021; Katsuda et al., 2013; Koniusz et al., 2016; Zheng et al., 2018). Isolated MSC supernatants or conditioned media can perform similar action compared with the actual MSCs, demonstrating that biomolecules and factors within the MSC-derived EVs are responsible for the effects of MSCs observed in many experiments (Bodart-Santos et al., 2019; Liau et al., 2020; Marino et al., 2019; Puig-Pijuan et al., 2020).
Patel et al. (2010) demonstrated that MSCs are protective to breast cancer cells through the release of several factors, including TGF-β. Tang et al. (2005) demonstrated that MSCs induce vascular regeneration in a model of myocardial infarction in rats through the release of various factors. Mao et al. (2021) showed that MSC-derived EVs can alleviate myocardial ischemia in mice. Ren et al. (2021) showed that MSC-derived EVs containing miR-29a-3p can protect against myocardial injury after severe acute pancreatitis.
Caution must be practiced, however, as the protection afforded to other cells by MSCs and their EVs can have a negative effect in cancer. For example, Li et al. (2021) showed that EVs from bone marrow-derived MSCs can enhance osteosarcoma cell proliferation, invasion, and migration in a mechanism involving the Wnt-β-catenin signaling. Furthermore, Guo et al. (2021) showed that MSC-derived EVs containing miR-130b-3p promotes the progression of lung cancer.
Mesenchymal Stem Cell Differentiation Potential
One of the criteria used to characterize MSCs is the ability to differentiate into multilineages (Fig. 5) (De Souza et al., 2016; Dennis and Charbord, 2002; Dzobo et al., 2016a, 2018; Liau et al., 2020). Several reports, including one by Dominici et al. (2006) spelt out the minimum criteria required for cells to be defined as MSCs as stipulated by the ISCT. As we showed in our early publication, this can be achieved through culturing the MSCs in differentiation media with specific supplements and then evaluating adipogenesis markers, osteogenesis markers, and chondrogenesis markers (Dzobo et al., 2016a). Adipogenic-inducing medium consisted of Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS), 1% 1 μM dexamethasone, 0.5 mM 3-isobutyl-methylxanthine, 200 μM indomethacin, and 10 μg/mL insulin.
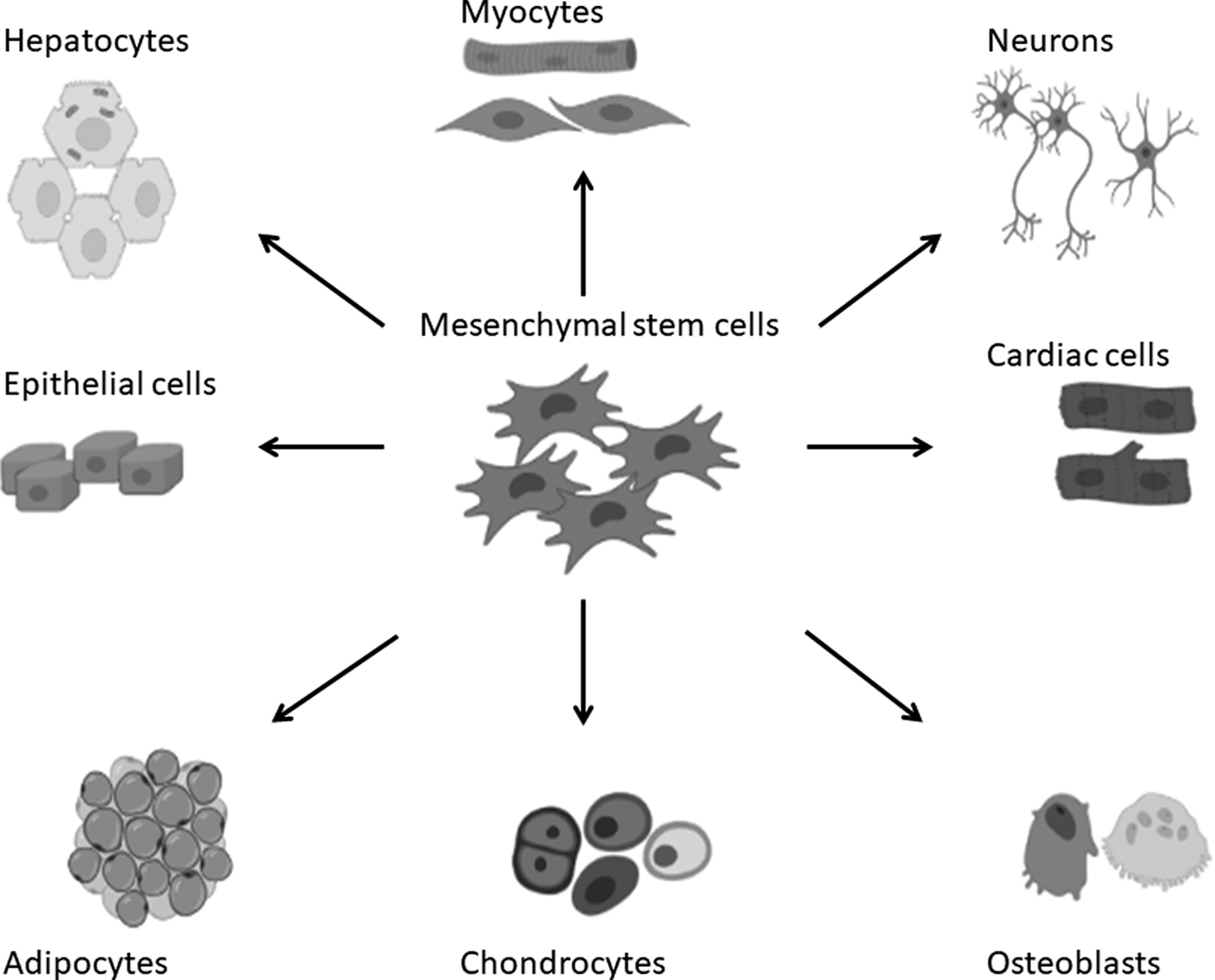
The differentiation potential of mesenchymal stem cells.
We utilized both real-time polymerase chain reaction (PCR) as well as staining for lineage-specific markers. For adipogenesis, we evaluated the levels of PPARG2 and lipoprotein lipase using real-time PCR as well as staining with Oil Red O for lipid droplets (Dzobo et al., 2016a). Osteogenic medium made up of DMEM with 10% FBS, 0.1 μM dexamethasone, 50 μM ascorbate-2-phosphate, and 10 mM β-glycerophosphate.
To evaluate osteogenesis markers, we used real-time PCR to measure the levels of CBFA1 and Osteocalcin as well as staining for mineralization with Alizarin Red S (Dzobo et al., 2016a). Chondrogenic medium made up of DMEM containing 0.1 μM dexamethasone, 50 μg/mL ascorbate-2-phosphate, 10 ng/mL TGF-β3, 40 μg/mL proline, 100 μg/mL pyruvate, and 1% insulin, human transferrin, and selenous acid premix universal culture supplement. To evaluate chondrogenesis markers, we used real-time PCR to measure the levels of type II collagen and Sox9 as well as staining with Toluidine Blue O for proteoglycans that are visible as purple (Dzobo et al., 2016a). These conditions have been utilized by several laboratories worldwide and are the stipulated conditions for such analysis (An et al., 2010; Mackay et al., 1998; Rosen and MacDougald, 2006).
Besides these differentiating conditions, MSCs are also characterized based on expression of several surface markers as we showed in our study. Specifically, our data show that our adipose-derived MSCs expressed markers such as CD73, CD90, and CD105, with no expression of CD34 and CD45 (Dzobo et al., 2016a).
In addition, we also utilized and characterized Wharton's Jelly-derived MSCs (Dzobo et al., 2016b). Flow cytometric analysis of Wharton's Jelly-derived MSCs using antibodies against CD44, CD45, CD73, CD90, and CD105, showed that our MSCs fulfilled the minimum requirements for MSCs as stipulated by the ISCT (Dzobo et al., 2016b). Cells characterized in our laboratory demonstrated the classical MSC phenotype of CD44+CD73+CD90+CD105+CD45− cell population (Dzobo et al., 2016b). Furthermore, lineage-specific differentiation capacity of Wharton's Jelly-derived MSCs fulfilled stipulated criteria by the ISCT.
In addition to having multilineage differentiation potential, MSCs have been shown to transform to other cell types, given the right cues or signal. As reviewed elsewhere, MSCs can be a source of fibroblasts, other stromal cells, and progenitors for cells making the body ligaments and connective tissue (Caplan and Dennis, 2006). Xu et al. (2004) demonstrated that bone marrow-derived MSCs can differentiate into cells with a cardiomyocyte phenotype. It is also possible to obtain liver cells, including hepatocytes as well as B cells of pancreatic islets from MSCs, provided the right supplements, such as EGF, nicotinamide, dexamethasone, insulin, transferrin, and oncostatin, are added to the culture media (Lee et al., 2004).
Indeed, MSCs from different sources have been shown to differentiate and to mature into pancreatic islets of B cells with the ability to produce insulin (Govindasamy et al., 2011; Phadnis et al., 2011). Naghdi et al. (2009) were able to differentiate MSCs from the bone marrow into cholinergic neurons in rats, clearly demonstrating the utility of MSCs for the treatment of spinal cord injury (Naghdi et al., 2009). Other cues necessary for the differentiation of MSCs into neurons include retinoic acid, valproic acid, EGF, hydrocortisol, and 5-azacytidine (Anghileri et al., 2008; Pavlova et al., 2012). The resulting neurons are able to express several markers of mature neurons such as neuron-specific enolase and Nestin.
Dzobo et al. (2016a) demonstrated that MSCs cultured on a 3D cell-derived matrix, to recapitulate in vivo physiological microenvironment, showed reduced expression of integrins α2 and β1 and subsequently differentiated toward the chondrogenic phenotype as evidenced by the loss of SOX2, and NANOG gene expression, as well as increase in markers such as Runx2, osteopontin, and p-TGF-β-RII. Furthermore, our study showed that the use of siRNA and a mutant Notch1 construct showed that the mechanism through which MSCs differentiated toward chondrogenic phenotype involved Notch1 and β-catenin signaling (Dzobo et al., 2016a). Overall, these and other studies indicate that Cs, given the right cues and signaling molecules, can differentiate into multilineage cells and form several tissues of the human body.
Challenges, Prospects, and Outlook
The discovery that the bone marrow contained more than just hematopoietic stem cells, initiated a frantic study of these cells, resulting in the realization that some cells within the bone marrow can form the connective tissue of the body. MSC research entered an exciting period mainly due to their appealing properties, including the easy accessibility and multilineage differentiation. Further studies revealed that MSCs have immunomodulatory properties and can be a source of many difficult-to-repair cells and tissues, astounding many scientists in the process. Ever since the discovery of MSCs, these cells have been receiving great attention in different fields of biology from regenerative medicine, cancer research, and even the treatment of infectious diseases (Dzobo et al., 2016b; Mendicino et al., 2014).
It is important that MSCs prepared for therapy are properly “synchronized” or “tuned” to provide optimum effect. Factors such as MSC isolation method, culture method, metabolic state, and doses used must be carefully considered during treatment. Thus, MSCs intended for different applications must be prepared differently to increase “therapeutic effect.” A knowledge of the “microenvironments” that MSCs are likely to encounter when in vivo, means that the MSCs can be subjected to the same treatments or conditions during preparation time.
To overcome challenges such as donor differences in MSCs characteristics, longitudinal culture analysis can be done together with genetic tagging. In addition, MSCs intended for tissue repair may benefit from “priming” for certain environments, can be grown on specific surfaces such as the ECM as we have done previously (Dzobo et al., 2016a). MSCs can be “tuned” for specific therapeutic use or they can be used to provide EVs with a specific biological factor(s) “package.”
Furthermore, cells do not perform tissue repair in isolation. If MSCs are intended for tissue repair or regenerative purposes, coculture or cotransplantation with other cells may increase “therapeutic effect” or engraftment and differentiation. Based on our studies and those by others, the use of decellularized ECM and other scaffolds may increase both MSC differentiation and engraftment (Dzobo et al., 2016a, 2016b, 2019b; Eliopoulos et al., 2010; Eom et al., 2015; Farrington-Rock et al., 2004).
Although MSCs can be isolated and purified effectively, they comprise a very small fraction of cells in tissues. Once isolated, MSCs will therefore require in vitro expansion, a process called Biobanking. In most cases, Good Manufacturing Practices are adopted and the isolated and expanded MSCs can be used in Clinical Trials or for patients' treatment in the clinic. Many clinical trials show that a specific number of cells are needed for effective stem cell therapies. Higher and lower cell numbers may not be optimal. To date, several MSC preparations have been approved by the Food and Drug Administration (FDA) or European Medicines Agency (EMA) for the treatment of various ailments.
Efforts are underway to optimize the in vitro expansion of MSCs for the best therapeutic effect in vivo. The process of preparing different MSCs for therapeutic use must be defined for each and every application for reproducibility purposes. One popular route for delivery of therapeutic MSCs is intravascular infusion as shown in many clinical trials. While studies show that infusion of bone marrow-derived MSC is safe, the same cannot be said about other MSC products from other sources. One major problem is the varied levels of procoagulant tissue factor, which can result in an inflammatory reaction.
To curtail the mushrooming of unapproved MSC treatment, new and strict regulatory practices are now in place. New rules require that any medical product preparation with living cells must first seek “investigational new drug” status before being used in clinical trials for example. The EMA has the Centralized Marketing Authorization, a series of steps involving quality checks, safety checks, and final approval before any MSC-containing product can be released to the market. Based on the current innovation trajectory, the MSC product market is poised for significant strides. Soon it appears many MSC products will likely be available in the clinic for the treatment of various conditions.
Many global health challenges, including diseases such as cancers, HIV/AIDS, tuberculosis, and degenerative diseases, still remain with modern medicine and drugs unable to provide adequate cures (Dzobo et al., 2019a; Dzobo, 2019; Thomford et al., 2018). Hence, the search for solutions continues and MSCs may offer a new and novel strategy to treat these conditions. Advances in MSC research in the public and industry sectors will transform medicine and provide innovative strategies for different medical conditions.
While many clinical trials are underway to evaluate the utility of MSCs in different applications, the mushrooming of scientifically unproven stem cell therapies calls for caution, and careful consideration of what these cells are able to do. As wound healing demonstrates that it is a complex process, MSCs should not be used carelessly or with little knowledge of their function and capabilities. Inappropriate use of stem cell therapies is actually a hindrance to proper stem cell research and can cause public distrust in science and the effectiveness of this therapy. Importantly, all stem cell products must be thoroughly characterized with traceable records on their preparation. Scientists researching on stem cell for therapeutic use must provide accurate records and evaluate each product at several stages of preparation.
Lastly, this article provides an update on the biology and therapeutic use of MSCs and their associated cells and new strategies being devised to utilize these cells in treating various human pathological conditions. While many clinical trials are underway, it is important that research into the various MSCs continue to enhance our understanding of these cells. MSCs are appealing by virtue of their differentiation potential, immunomodulatory activities, paracrine effects, as well as interactions with other cells. MSCs thus provide exciting and innovative potentials to treat the many diseases afflicting humans.
Footnotes
Author Disclosure Statement
The author declares there are no conflicting financial interests.
Funding Information
No funding was received for this article.