
Abstract
Gastrointestinal cancers pose a significant global health challenge. N-glycosylation modulates various cellular processes, including key cancer-related mechanisms. Elucidating its involvement in the onset and advancement of these cancers can offer critical insights for enhancing diagnostic and therapeutic approaches. This review outlines the core process of protein N-glycosylation and highlights its contribution to the progression of gastrointestinal cancers, encompassing cell proliferation, survival, invasion, metastasis, and immune evasion, mainly through its impact on critical signaling pathways. Notably, aberrant N-glycosylation patterns have emerged as crucial biomarkers for the diagnosis and prognosis of various gastrointestinal cancers, providing the foundation for more personalized therapeutic approaches. Therapeutic strategies targeting N-glycosylation, such as glycosyltransferase inhibitors and glycoengineering, show significant promise in mitigating tumor aggressiveness and enhancing immune recognition. However, the clinical implementation of N-glycosylation biomarkers requires the standardization of glycosylation analysis techniques and solutions to challenges in sample processing and data interpretation. Future research efforts should concentrate on overcoming these obstacles to unlock the full potential of N-glycosylation in enhancing cancer management and advancing patient outcomes.
Introduction
Gastrointestinal cancers, including colorectal, gastric, liver, pancreatic, and esophageal cancers, represent a significant global health challenge due to their high incidence (24% of new cancer cases) and mortality (33.3% of deaths) among the 36 identified types of cancer (Bray et al., 2024). In 2022, colorectal cancer was identified as the third most prevalent cancer worldwide and the second leading cause of cancer-related deaths. Similarly, gastric cancer was ranked as the fifth most common and the fourth leading cause of cancer-related deaths, particularly in East Asia. Additionally, liver cancer, although ranked sixth in incidence, held the third position in terms of mortality, reflecting its poor prognosis. Pancreatic cancer, also known for its poor prognosis, was identified as the sixth leading cause of cancer-related deaths, accounting for nearly 5% of all cancer fatalities worldwide. Lastly, esophageal cancer, ranked as the 11th most frequently diagnosed cancer, was the 7th leading cause of cancer-related mortality (Bray et al., 2024).
The management of these cancers largely relies on early detection through screening techniques such as colonoscopy, endoscopy, and ultrasound and imaging methods such as computed tomography and magnetic resonance imaging. Treatment strategies include surgery, chemotherapy, radiation therapy, and targeted therapies (Anwanwan et al., 2020; Biller and Schrag, 2021; Dekker et al., 2019; Fu and Wang, 2018; Goggins et al., 2020; Haque et al., 2022; Liu et al., 2023; Obermannová et al., 2022; Tanaka et al., 2017). Recent studies investigating the mechanisms underlying tumorigenesis have significantly advanced our understanding of gastrointestinal cancers (Huang et al., 2023; Lei et al., 2022; Liu et al., 2020; Nguyen et al., 2020; Rebouissou and Nault, 2020). However, accurately predicting tumorigenesis and prognosis using multiomics data remains a challenge.
In this context, the concepts of the “sugar code” and the “paracentral dogma” (Özdemir et al., 2020; Wang, 2023b) may provide a promising framework for systematically interpreting the occurrence, progression, prognosis, and therapeutic strategies in gastrointestinal cancers. This review seeks to elucidate the role of protein N-glycosylation in the biology of these cancers and discuss the potential of N-glycosylation as a biomarker and therapeutic target while addressing challenges in its clinical implementation.
Glycosylation
Glycosylation is a critical post-translational modification (PTM) that involves the addition of carbohydrate chains to proteins or lipids, significantly influencing their function, stability, localization, and interactions (Haltiwanger and Lowe, 2004; Varki, 2017; Wang, 2023b). This process initiates with the attachment of glycans to nascent proteins in the endoplasmic reticulum (ER), followed by further modifications in the Golgi apparatus, where various glycosyltransferases and glycosidases act to refine these structures (Varki, 1998; Varki, 2011; Zhang and Wang, 2016). Glycosylation can be categorized into several types based on the attachment of glycans to proteins, including N-glycosylation, O-glycosylation, C-glycosylation, glypiation, and phosphoglycosylation, with N-glycosylation and O-glycosylation being the most prevalent (Haltiwanger and Lowe, 2004; Reily et al., 2019; Stanley, 2011). These glycosylation types collectively contribute to the diverse and complex roles of proteins in biological systems.
Unlike DNA synthesis, which follows a template-driven mechanism, glycosylation is a nontemplate-driven process, resulting in a wide diversity and heterogeneity of glycan structures (Lin and Lubman, 2024; Wang, 2023a). This structural diversity becomes particularly pronounced under pathological conditions, where aberrant glycosylation patterns can emerge (He et al., 2024; Lin and Lubman, 2024; Reily et al., 2019).
N-Glycosylation
N-glycosylation is a crucial PTM that is ubiquitously observed in eukaryotic cells. N-glycosylation occurs on the nitrogen atom of asparagine (Asn) residues, forming complex glycan structures that are critical for protein folding, stability, and membrane interactions (Lin and Lubman, 2024). This modification is also crucial in regulating the immune system’s ability to recognize antibodies, thereby affecting immune responses and the progression of different diseases (Lin and Lubman, 2024).
N-glycosylation is initiated in the ER when the oligosaccharyltransferase complex transfers a lipid-linked oligosaccharide precursor, Glc3Man9GlcNAc2, to an Asn residue within a defined sequon (N-X-S/T) of the nascent polypeptide chain. The process begins with the synthesis of GlcNAc2Man5 on dolichol phosphate, located on the cytosolic side of the ER membrane. It is then translocated to the lumen, where additional mannose and glucose residues are added. Following this, glucose residues are sequentially removed by two α-glucosidases (α-Glu I and α-Glu II), and a mannose residue is trimmed by ER α-mannosidase (ER α-Man). Once the glycoprotein passes the ER’s quality-control mechanisms, it is transported to the Golgi apparatus, where further modifications occur, such as mannose trimming by α-Man I and II (Hirata and Kizuka, 2021; Reily et al., 2019). This is followed by additional glycosylation steps catalyzed by various glycosyltransferases, including GlcNAc-transferases I and IV, galactosyltransferases, and sialyltransferases (Stanley, 2011).
This intricate processing results in the production of diverse N-glycoforms, with the final structure shaped by glycosyltransferase expression levels, the accessibility of glycosylation sites, and the duration the glycoprotein spends in the ER and Golgi apparatus (Fig. 1) (Lin and Lubman, 2024; Reily et al., 2019). This process plays a pivotal role in various biological functions, including protein folding, stability, cell–cell communication, immune response, and signal transduction (He et al., 2024; Stanley, 2011; Zhang and Wang, 2016).

N-glycosylation process from ER initiation to Golgi maturation. N-glycosylation is initiated in the ER, where the OST complex transfers a lipid-linked oligosaccharide precursor, Glc3Man9GlcNAc2, onto an asparagine residue of the newly formed peptide chain. The process begins with the construction of GlcNAc2Man5 on dolichol phosphate at the cytosolic side of the ER, followed by its translocation to the luminal side for the addition of further mannose and glucose units. The glycoprotein then undergoes sequential trimming, with glucose residues being removed by two α-glucosidases and a mannose residue by ER α-Man. After passing a quality control checkpoint, the glycoprotein is transported to the Golgi apparatus, where it undergoes additional processing and is modified by various glycosyltransferases. α-Glc, α-glucosidases; α-Man, α-mannosidase; ER, endoplasmic reticulum; GlcNAc, N-acetylglucosamine; OST, oligosaccharyltransferase. This figure was created with BioRender.com.
N-glycosylation can be subdivided into three primary types based on the branching architecture of the glycan chains: high-mannose N-glycans, complex N-glycans, and hybrid N-glycans (Fig. 1). High-mannose N-glycans are formed through the extension of mannose residues, whereas complex N-glycans are further elongated in the Golgi apparatus by the addition of N-acetylglucosamine (GlcNAc); hybrid N-glycans are created by incorporating galactose or other core residues along with mannose in the Golgi apparatus (Lin and Lubman, 2024; Zhang and Wang, 2016). N-glycosylation modulates the three-dimensional structure and functional properties of proteins by influencing the glycosylation processes within the ER and Golgi apparatus. This regulation affects the expression and localization of glycosyltransferases and glycosidases, thereby impacting multiple cellular processes (He et al., 2024; Helenius and Aebi, 2004).
We conducted a comprehensive search in major databases, including PubMed and Web of Science. Our search utilized keywords such as “N-glycosylation,” “N-glycans,” or “N-glycome,” combined with terms related to gastrointestinal cancer to maximize comprehensiveness. According to the PRISMA 2020 statement (Page et al., 2021), the following criteria were used to include studies and publications in this review: (1) study design: all original research articles, such as clinical studies, in vitro experiments, animal studies, and systematic reviews/meta-analyses focusing on N-glycosylation in gastrointestinal cancers; (2) population: studies involving patients or models related to colorectal, gastric, liver, pancreatic, and esophageal cancers; (3) topic relevance: articles addressing the biological role, mechanisms, clinical implications, or therapeutic potential of N-glycosylation; (4) language: publications available in English; and (5) publication date: studies published from 2014 to 2024, except where older studies provide essential foundational knowledge.
We found that, in the past decade, numerous studies have expanded our understanding of the crucial role N-glycosylation plays in the development and progression of gastrointestinal cancers. Aberrant N-glycosylation patterns have been shown to significantly influence tumor growth, metastasis, and immune evasion across various cancer types. For example, increased core-fucosylation has been observed in gastric cancer and is associated with poor prognosis, whereas elevated levels of α2,6-sialylated N-glycans have been linked to lymph node metastasis in colorectal cancer (Holst et al., 2015). Additionally, recent research has identified specific glycosylation changes in hepatocellular carcinoma (HCC), whereas altered glycan structures contribute to immune suppression and chemoresistance (Lin et al., 2022). These findings suggest that N-glycosylation not only serves as a biomarker for early detection but also plays an active role in modulating tumor behavior. The integration of glycomics with advanced analytical techniques, such as mass spectrometry (MS) and glycoproteomics, has facilitated the identification of novel N-glycosylation alterations, offering insights into potential therapeutic targets and prognostic indicators for gastrointestinal cancers (Wang et al., 2021).
Mechanisms and Functions of N-Glycosylation in Gastrointestinal Cancers
Regulation of tumor proliferation and survival
N-glycosylation is essential for the proliferation and survival of gastrointestinal cancer cells by modulating various cellular signaling pathways and receptor functions (Pérez et al., 2020). The primary mechanisms involved are as follows.
Regulation of receptor tyrosine kinase activity
The N-glycosylation of receptor tyrosine kinases (RTKs) typically involves complex-type N-glycans, which include branched structures, including bi-antennary, tri-antennary, and tetra-antennary structures. These glycan structures influence the folding, localization, and function of RTKs such as the epidermal growth factor receptor (EGFR) and human epidermal growth factor receptor 2 (HER2), thereby enhancing their signaling capacity (Fig. 2; Fujitani et al., 2024). The persistent activation of these receptors promotes proliferative signaling while inhibiting apoptotic pathways, thus enabling tumor cells to survive in a hostile microenvironment. For instance, aberrant N-glycosylation of EGFR has been associated with its overactivation in various gastrointestinal cancers, correlating with increased tumor growth rates and resistance to therapies (Fernández-Ponce et al., 2021).
Promotion of PI3K/AKT and RAS/MAPK pathway activity
N-glycosylation enhances the activity of the phosphoinositide 3-kinase/protein kinase B (PI3K/AKT) and rat sarcoma/mitogen-activated protein kinase (RAS/MAPK) signaling pathways by modulating the function of RTKs (Fig. 2). These pathways are fundamental to regulating cell proliferation, survival, differentiation, and metabolism, with their continuous activation being strongly linked to the malignant progression of numerous cancers (Chandler et al., 2017).
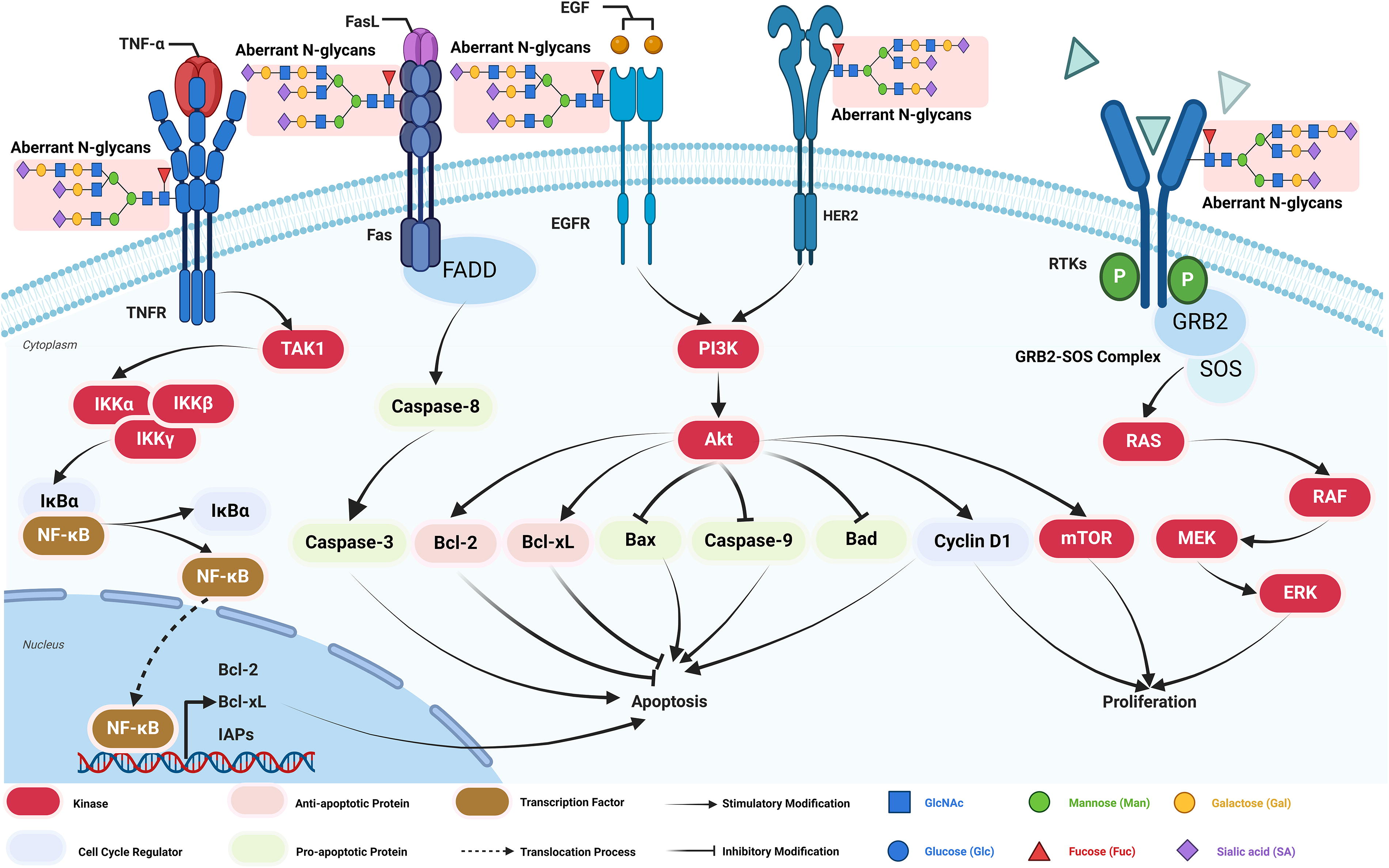
N-glycosylation of RTKs enhances their activity by facilitating more efficient ligand binding, receptor dimerization, and autophosphorylation, leading to the activation of the PI3K/AKT signaling pathway. This sustained activation of AKT promotes the expression of proliferative genes such as the mechanistic target of rapamycin and Cyclin D1 (Wu et al., 2024). This mechanism is particularly evident in gastrointestinal cancers, such as colorectal and gastric cancers. For instance, in colorectal cancer, N-glycosylation of EGFR amplified the PI3K/AKT pathway’s activity, contributing to enhanced cancer cell growth (Lau and Dennis, 2008).
N-glycosylation of RTKs facilitates the activation of RAS through the GRB2-SOS complex, subsequently triggering the RAF-MEK-extracellular signal-regulated kinase (ERK) signaling cascade (Rodrigues et al., 2018). Sustained ERK activity enhances cellular proliferation and migration. In gastrointestinal cancers, the overactivation of the RAS/MAPK pathway is closely linked to aberrant N-glycosylation. For example, in gastric cancer, aberrant N-glycosylation of HER2 amplified signaling through this pathway, promoting malignant behavior in cancer cells (Pinho and Reis, 2015).
Enhancement of antiapoptotic signaling
N-glycosylation can potentiate antiapoptotic signaling by modulating the function of death receptors or associated signaling molecules, thereby extending the survival of tumor cells. Specific N-glycosylation patterns may directly or indirectly inhibit the activation of apoptotic signaling pathways, contributing to tumor persistence (Lin and Lubman, 2024).
Proper glycosylation of the Fas receptor is essential for initiating apoptotic signaling; however, in cancer, aberrant N-glycosylation may prevent Fas receptor activation, thereby suppressing the initiation of apoptotic signals. This modulation of death receptor N-glycosylation allows tumor cells to evade Caspase-mediated apoptosis, granting them a survival advantage in malignant environments (Fig. 2). For example, in gastric cancer, changes in the N-glycosylation of the Fas receptor were believed to contribute to the inhibition of Fas-mediated apoptotic pathways, thereby enhancing the antiapoptotic capabilities of cancer cells (Pinho and Reis, 2015).
N-glycosylation significantly enhances the antiapoptotic PI3K/AKT pathway by promoting AKT activation. Glycosylated (RTKs, such as EGFR, increase PI3K activity, leading to the phosphorylation and activation of AKT. Activated AKT inhibits proapoptotic factors through various mechanisms, including the suppression of Caspase-9 and B-cell lymphoma 2 (Bcl-2)-associated death promoter (Bad) activity (Fig. 2; Itkonen and Mills, 2013). Furthermore, AKT signaling influences the expression of Bcl-2 family proteins by promoting the production of antiapoptotic proteins, such as Bcl-2 and B-cell lymphoma-extra large (Bcl-xL) while suppressing proapoptotic proteins like Bcl-2-associated X protein (Bax), thereby inhibiting apoptosis (Fuster and Esko, 2005; Lau and Dennis, 2008). For instance, in colorectal cancer, the N-glycosylation of EGFR has been associated with enhanced PI3K/AKT signaling, which significantly increased the antiapoptotic capabilities of cancer cells (Lau and Dennis, 2008).
N-glycosylation engages with the nuclear factor kappa B (NF-κB) signaling pathway by promoting NF-κB activation through improved functionality of cytokine receptors, including the tumor necrosis factor (TNF) receptor (Pan et al., 2014). NF-κB is a crucial transcription factor that bolsters antiapoptotic capabilities by upregulating various antiapoptotic genes, including Bcl-2, Bcl-xL, and inhibitor of apoptosis proteins (Fig. 2). NF-κB signaling activation is essential for sustaining an antiapoptotic environment in gastrointestinal cancers, allowing cancer cells to endure unfavorable conditions and resist apoptosis triggered by chemotherapeutic treatments (Fuster and Esko, 2005). For instance, in liver cancer, the N-glycosylation of the TNF receptor significantly enhanced the expression of antiapoptotic genes through NF-κB activation, thereby increasing the survival capacity of tumor cells (Lambrecht et al., 2023).
Regulation of tumor invasion and metastasis
N-glycosylation is pivotal in controlling tumor invasion and metastasis in gastrointestinal cancers, primarily by influencing the behavior of cell adhesion molecules, matrix metalloproteinases (MMPs), and the epithelial–mesenchymal transition (EMT) process (Zhang et al., 2020).
N-glycosylation influences the function of cell adhesion molecules, such as E-cadherin. Alterations in the N-glycosylation of E-cadherin can impair cell–cell adhesion, thereby facilitating the detachment and migration of tumor cells (Fig. 3; Lin and Lubman, 2024; Pinho and Reis, 2015). For example, in gastric cancer, reduced levels of N-glycosylation of E-cadherin led to decreased cell adhesion, thereby promoting tumor cell invasion and metastasis (Lau and Dennis, 2008).
N-glycosylation regulates the function of MMPs, essential enzymes responsible for breaking down the extracellular matrix (ECM), thereby promoting tumor invasion by allowing cancer cells to breach the basement membrane and infiltrate adjacent tissues (Fig. 3; Lin and Lubman, 2024; Pinho and Reis, 2015). For instance, in colorectal cancer, enhanced N-glycosylation of MMP-9 increased its enzymatic activity, thereby accelerating ECM degradation and promoting cancer cell metastasis (Pinho and Reis, 2015). Similarly, in esophageal cancer, aberrant N-glycosylation of MMP-9 was demonstrated to enhance its enzymatic activity, resulting in increased ECM degradation and facilitating tumor cell invasion (Pan et al., 2023).
N-glycosylation regulates key molecules involved in EMT, such as TGF-β receptors, thereby enhancing the migratory and invasive capabilities of tumor cells (Fig. 3). EMT is a process where epithelial cells transition into a mesenchymal state, gaining increased migratory properties that are vital for tumor metastasis (Fuster and Esko, 2005; Lin and Lubman, 2024). For example, in esophageal cancer, alterations in the N-glycosylation of TGF-β receptors promoted EMT, making tumor cells more prone to invasion and metastasis (Fuster and Esko, 2005).
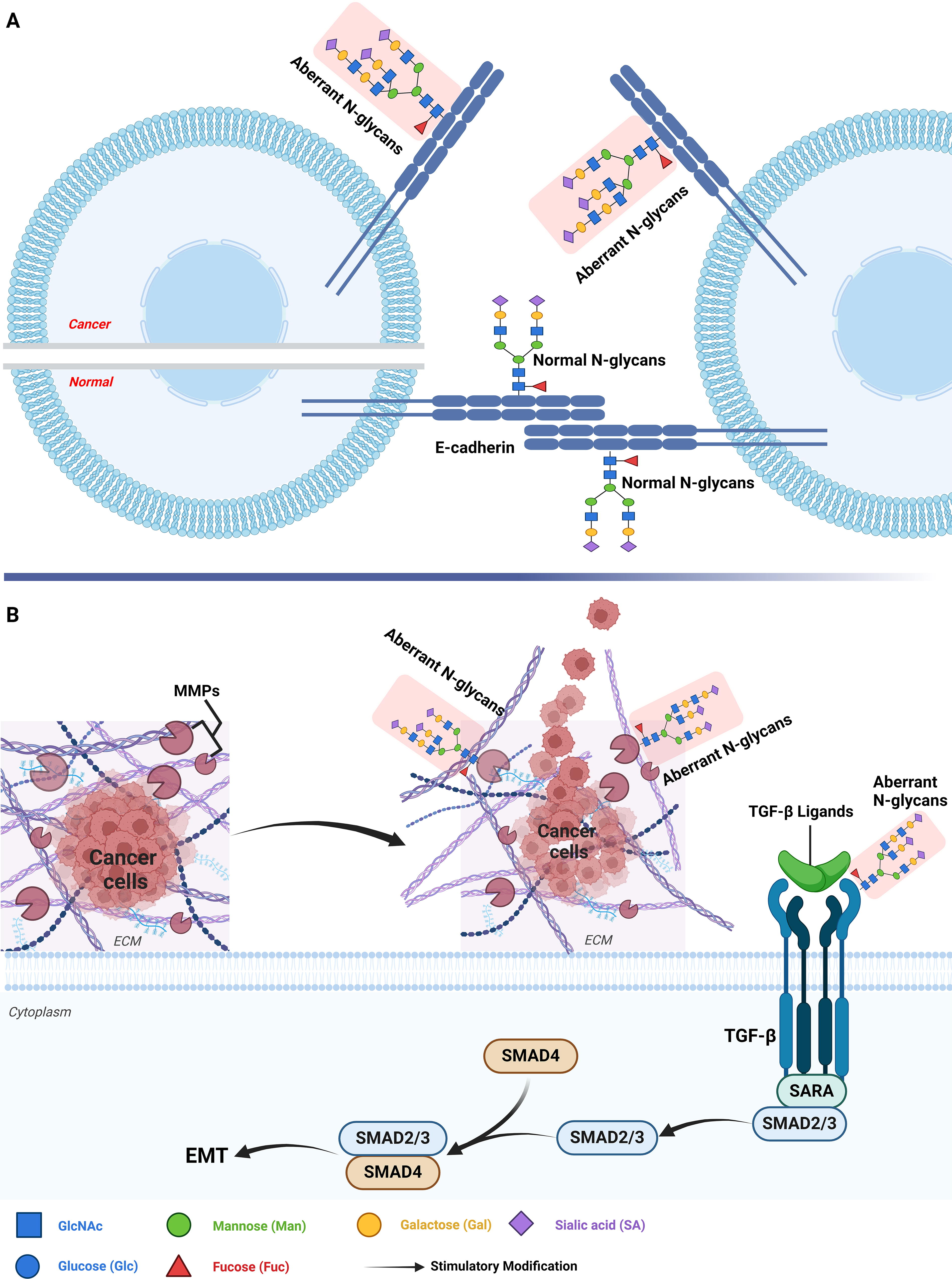
N-glycosylation in regulating invasion and metastasis in gastrointestinal cancers. (
Influence of tumor microenvironment
N-glycosylation is essential in shaping the tumor microenvironment in gastrointestinal cancers by influencing several mechanisms, such as altering cell–cell interactions, facilitating immune evasion, and remodeling the ECM.
N-glycosylation influences the function of cell surface molecules, such as integrins and selectins, thereby modifying interactions between endothelial cells, stromal cells, immune cells, and tumor cells (Lin and Lubman, 2024). These disrupted interactions aid in tumor progression by enabling immune evasion, allowing cancer cells to escape immune surveillance and destruction, and promoting ECM remodeling, which creates a conducive environment for tumor growth and invasion (Lau and Dennis, 2008; Pinho and Reis, 2015). For instance, in colorectal cancer, the N-glycosylation of integrins has been found to strengthen the adhesion between tumor cells and stromal cells, facilitating tumor invasion and metastasis (Pinho and Reis, 2015).
Influence of tumor immune evasion mechanisms
N-glycosylation significantly impacts immune evasion in gastrointestinal cancers through multiple mechanisms, including the modulation of immune checkpoint molecules, alteration of immune cell functions, and modifications in the composition of the tumor microenvironment.
Regulation of immune checkpoint molecule expression
N-glycosylation facilitates immune evasion by enhancing the stability and functionality of immune checkpoint molecules. For instance, N-glycosylation of programmed death-ligand 1 (PD-L1), an essential immune checkpoint molecule, enhances its binding affinity to the programmed cell death protein 1 receptor. This binding inhibits T-cell activation and impairs their immune function (Fig. 4) (Li et al., 2016; Pinho and Reis, 2015; Wang et al., 2020). In gastric cancer, elevated levels of N-glycosylation on PD-L1 increased its surface stability and binding capacity, thereby leading to the suppression of T cell-mediated immune responses (Wang et al., 2020).
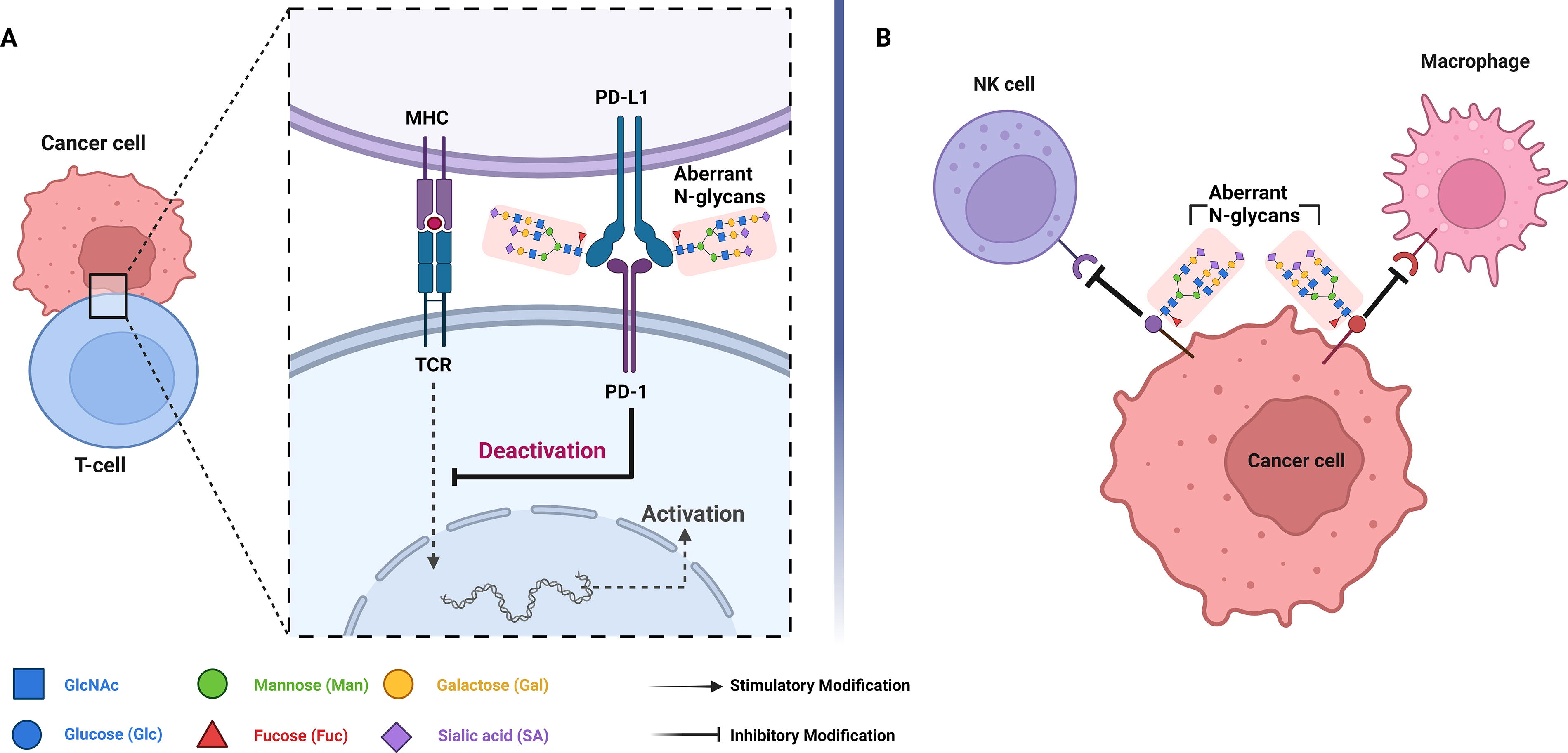
N-Glycosylation in influencing the gastrointestinal immune evasion. (
Impact on immune cell function
N-glycosylation modifies the structural conformation of tumor cell surface antigens, influencing their recognition and subsequent targeting by immune cells, such as macrophages and natural killer (NK) cells. Aberrant glycosylation patterns on tumor cells can inhibit NK-cell recognition, thereby promoting tumor growth and metastasis (Fig. 4; Boligan et al., 2015). For example, in colorectal cancer, abnormal N-glycosylation patterns enable tumor cells to evade detection by NK cells, further enhancing immune evasion (Zhang et al., 2024).
N-Glycans as Biomarkers in Gastrointestinal Cancers
N-Glycans as diagnostic biomarkers in gastrointestinal cancers
N-glycans are essential in the diagnosis and prognosis of gastrointestinal cancers. Abnormal glycosylation of cellular proteins leads to the generation of neoglycoforms, which can be released into the bloodstream and act as potential cancer biomarkers. Currently, specific glycoproteins, such as AFP and AFP-L3 (a1,6-fucosylated), are well-established as biomarkers in patients with HCC (Aoyagi et al., 1985; Sato et al., 1993), colon cancer (carbohydrate antigen 19-9 [CA19-9] and carcinoembryonic antigen), and pancreatic cancer (CA19-9).
However, their clinical application remains constrained due to several practical challenges. The validation of glycosylation biomarkers is hampered by the inherent variability in glycosylation profiles across different individuals (Reily et al., 2019). Factors such as genetic background, diet, and disease state can significantly influence glycan structures. This variability complicates the establishment of standardized glycan profiles that can reliably differentiate cancerous from noncancerous conditions. Additionally, sampling errors, such as variations in sample handling, storage, and processing, can lead to inconsistent glycosylation measurements, affecting the reproducibility of results (Clerc et al., 2016; Ma et al., 2023).
Interlaboratory consistency poses another significant challenge in translating glycosylation biomarkers into clinical practice. Differences in analytical methods, such as MS, HPLC, and lectin-based assays, can lead to variations in glycan detection and quantification (Huhn et al., 2009; Yang et al., 2013). Standardizing analytical techniques and establishing consensus protocols are critical for overcoming these limitations and achieving reliable biomarker validation.
Emerging data have shown distinct variations in serum glycoproteins between patients with cancer and healthy individuals, suggesting that new glycoproteins or glycopeptides may serve as promising biomarkers for gastrointestinal cancers (Lin et al., 2022). For instance, elevated levels of core-fucosylated glycans have been identified as potential diagnostic markers for early-stage gastric cancer and colorectal cancer (Holst et al., 2015; Pinho and Reis, 2015). Additionally, the upregulation of glycosyltransferases, such as N-acetylglucosaminyltransferase V (MGAT5), has been observed in colorectal cancer, indicating its potential as a diagnostic marker for early-stage disease (Ashkani and Naidoo, 2016).
N-Glycans as prognostic biomarkers in gastrointestinal cancers
The prognostic value of N-glycosylation patterns in gastrointestinal cancers is also well-recognized. Studies have shown that aberrant N-glycan profiles correlate with aggressive tumor behavior and patient survival rates. For example, increased expression of sialylated antigens, such as sialyl-Lewis a [sLe(a)] and sialyl-Lewis x [sLe(x)], is associated with poor prognosis across different cancer types, including colorectal cancer (Pinho and Reis, 2015). In particular, elevated sLe(x) antigen levels are linked to increased tumor invasiveness and metastatic potential (Pinho and Reis, 2015).
In colorectal cancer, the enzymatic activity of β-galactoside α2,3-sialyltransferase 1 (ST3Gal-I) and ST3Gal-II is significantly higher in cancer tissues than in normal mucosa (Schneider et al., 2001). Additionally, upregulated expression of N-acetylgalactosaminide α-2,6-sialyltransferase 2 (ST6GalNAc-II) in patients with lymph node metastasis suggests its potential as a prognostic marker (Schneider et al., 2001). Experimental studies have further demonstrated that transfecting ST6GalNAc-II into colon cancer cells increases the expression of the Thomsen–Friedenreich (TF) antigen, whereas transfection with ST3Gal-I or ST3Gal-II does not impact TF expression, underscoring the unique role of ST6GalNAc-II in colorectal cancer progression (Ishikawa et al., 2004; Schneider et al., 2001).
The translation of N-glycosylation biomarkers into clinical settings requires addressing the above challenges by implementing strategies to minimize interindividual variability and ensure consistency across laboratories. Developing robust quality control measures, establishing glycan reference standards, and harmonizing glycan analysis protocols will be key steps in enabling the widespread adoption of glycosylation biomarkers. Additionally, prospective studies with larger patient cohorts and standardized workflows are needed to validate these biomarkers’ clinical utility and establish reliable thresholds for diagnostic and prognostic applications.
N-Glycosylation as Potential Therapeutic Targets in Gastrointestinal Cancers
Therapeutic strategies targeting the N-glycosylation pathway in gastrointestinal malignancies have garnered increasing attention, demonstrating substantial potential for clinical application. N-glycosylation plays a crucial role in modulating cancer cell behaviors such as proliferation, invasion, immune evasion, and metastasis, making it an attractive target for therapeutic interventions. Two main approaches under investigation, glycosyltransferase inhibitors and glycoengineering strategies, offer promising avenues for modulating aberrant N-glycosylation in cancer cells, thereby potentially enhancing the efficacy of cancer treatments.
Glycosyltransferase inhibitors focus on disrupting the enzymes responsible for specific N-glycan modifications, which are often upregulated in tumor cells. These enzymes are critical for maintaining abnormal glycosylation patterns associated with cancer progression, and their inhibition can significantly impair tumor growth and spread. For example, inhibition of α-1,6-fucosyltransferase (FUT8), a key glycosyltransferase involved in core fucosylation, has been shown to markedly reduce tumor growth and metastasis in preclinical models of gastrointestinal cancer by affecting cell signaling and adhesion pathways (Bangarh et al., 2023; Thomas et al., 2021). Similarly, targeting other enzymes, such as MGAT5 and ST6Gal-I, which are associated with the biosynthesis of complex N-glycan structures, has been proposed as a strategy to diminish cancer cell invasiveness and promote sensitivity to chemotherapeutic agents (Chen et al., 2016).
Glycoengineering aims to modify or remodel the glycosylation patterns of tumor cells to alter their biological behavior and improve immune recognition. By reshaping the glycan structures presented on cancer cell surfaces, glycoengineering can enhance immune system recognition and clearance of tumor cells (Pereira et al., 2018). For instance, modifying N-glycosylation profiles to reduce sialylation and increase core fucosylation has been shown to enhance antibody-dependent cellular cytotoxicity in colorectal cancer, improving the therapeutic efficacy of monoclonal antibodies (Pereira et al., 2018). Another approach involves engineering glycosylation pathways to increase the presentation of immunogenic glycans on cancer cells, thereby promoting immune system activation and reducing tumor immune evasion (Itano, 2019; Pereira et al., 2018).
The clinical relevance of targeting N-glycosylation extends beyond enhancing the effectiveness of existing therapies. These strategies could potentially be used to overcome resistance to conventional treatments, such as chemotherapy and immune checkpoint inhibitors, by modifying the tumor microenvironment to be more conducive to immune infiltration and attack (Liu et al., 2022). Furthermore, the ability to selectively inhibit specific glycosyltransferases or engineer tumor glycosylation profiles opens avenues for developing personalized therapies tailored to individual patients’ glycomic signatures. However, the clinical application of these approaches is not without challenges. Key issues include the heterogeneity of glycosylation patterns across different tumor types and stages, variability in glycan structures among patients, and the need for standardized methods for detecting and quantifying glycosylation changes (Pinho and Reis, 2015; Reily et al., 2019). Additionally, off-target effects of glycosylation inhibitors must be carefully monitored, given the ubiquitous nature of glycosylation in normal physiological processes.
In summary, therapeutic targeting of the N-glycosylation pathway in gastrointestinal cancers, whether through glycosyltransferase inhibitors or glycoengineering strategies, represents a promising avenue for improving cancer therapies. By inhibiting key enzymes such as FUT8 or modifying glycosylation patterns, these approaches hold the potential to reduce tumor aggressiveness, improve immune system recognition, and provide more effective, personalized therapeutic options.
Optimization and Validation of N-Glycosylation Analysis
The precision and repeatability of glycosylation analysis are significantly influenced by the chosen analytical techniques, including microarray technologies, high-throughput platforms, MS, liquid chromatography–MS (LC-MS), and ultra-performance liquid chromatography (UPLC) (Reiding et al., 2014; Reusch et al., 2015; Selman et al., 2012; Wang, 2019; Zhang et al., 2018). To ensure consistent data interpretation, establishing standardized procedures across laboratories is essential. Variability in sample sourcing and processing, including differences in sample types (plasma, serum, and tissue) and pretreatment steps (e.g., protein purification and deglycosylation), can introduce biases that compromise data quality and comparability (Reusch et al., 2015; Selman et al., 2012). To address these challenges, it is imperative to develop standardized protocols for sample collection and handling to enhance data reliability. Additionally, the interpretation of glycosylation data must take into account influencing factors such as age, gender, and disease status, highlighting the need for large-scale multicenter studies with robust statistical analyses to mitigate potential biases and improve clinical applicability (Reiding et al., 2014; Selman et al., 2012).
Establishing the reliability and clinical utility of N-glycosylation analysis through stringent validation methods is essential for its incorporation into cancer management strategies. Validation efforts include the standardization of glycosylation analysis techniques such as MS, LC-MS, UPLC, and glycan microarray technology, enabling the precise characterization of glycosylation profiles and the identification of glycoforms associated with cancer phenotypes and treatment responses (Reiding et al., 2014; Reusch et al., 2015; Selman et al., 2012; Zhang et al., 2018). These studies aim to correlate glycosylation patterns with clinical outcomes such as prognosis, therapeutic response, and disease progression, with large-scale cohort studies across various cancer types providing the necessary statistical power.
Longitudinal studies are critical for evaluating the stability of glycosylation profiles over time and their sensitivity to changes in disease status or treatment. Although validated glycosylation biomarkers offer great potential for personalized cancer care, there are still challenges in standardizing sample collection, interpreting data, and incorporating these biomarkers into regular clinical practices (Reiding et al., 2014; Reusch et al., 2015). Future research should focus on expanding validation efforts across diverse patient populations, refining analytical techniques, and exploring novel glycomics technologies to enhance the reliability and clinical applicability of N-glycosylation biomarkers, thereby optimizing cancer diagnostics and therapeutic strategies.
Conclusion and Future Perspectives
Recent glycomics research has shown that N-glycosylation is crucial in the onset, advancement, and prognosis of gastrointestinal cancers, underscoring its significant potential as both a biomarker and a therapeutic target. The analysis of glycosylation patterns offers the potential to tailor treatment approaches, enhancing both specificity and efficacy. However, integrating these biomarkers into clinical practice presents several challenges, including the need for standardization of analytical techniques, consistency in validation methods, development of standardized sample handling protocols, and addressing data interpretation complexities.
Future research should prioritize specific glycosylation profiles known to be associated with cancer progression and treatment response, such as core-fucosylated N-glycans, bisecting GlcNAc structures, and sialylated glycoforms, particularly sLe(a) and sLe(x) antigens. These profiles have shown potential as diagnostic and prognostic biomarkers in gastrointestinal cancers and warrant further investigation. Expanding validation studies across various gastrointestinal cancer types, including colorectal, gastric, pancreatic, and hepatocellular cancers, should be a key focus. Additionally, computational modeling and machine learning algorithms should be employed to integrate glycosylation data with other omics and clinical parameters, improving the clinical applicability and predictive capability of N-glycosylation biomarkers. These efforts aim to ensure the robustness of glycosylation-based biomarkers in gastrointestinal oncology, optimize personalized treatment strategies, and ultimately improve patient prognosis and survival outcomes.
Footnotes
Acknowledgments
The authors thank SUMC-ECU collaborative PhD project for providing financial support.
Authors’ Contributions
R.X., L.B., G.C., and M.S. contributed to the discussion and conception of the original idea. R.X. wrote the article. L.B., G.C., and M.S. reviewed and edited it. All authors contributed to the drafts and approved the final article.
Author Disclosure Statement
The authors declare that there are no conflicting financial interests.
Funding Information
This work was supported by Doctoral Program of Guangdong Science and Technology Special Fund (