
Abstract
Introduction
Numerous inflammatory pathologies are associated with an overproduction of cytokines such as tumor necrosis factor (TNF)-α and interleukin (IL)-1β. 5 IL-6 is induced during acute-phase reactions and is usually expressed in response to or together with IL-1 and TNF-α. 6 However, contradictory effects have been reported 7 ; it suppresses TNF-α, IL-1, and IL-12. Its vital role in wound healing is its ability to cause cell differentiation and proliferation. Inflammatory cytokines such as IL-1β and TNF-α have been shown to be increased in non-healing wounds, as well as in diabetic patients. 8 TNF-α is the most critical accelerator of diabetes. 9 Nuclear factor – kappa B (NF-κB) can be activated by IL-1β and TNF-α. Synthesis of IL-1β, TNF-α, and IL-6 is mediated by NF-κB. This therefore renders it the most important regulator of pro-inflammatory gene expression. 10 NF-κB pre-exists in the cytoplasm in a dormant form bound to inhibitor protein κB (IκB). Following degradation of IκB, NF-κB is liberated and translocated to the nucleus, binds to the consensus sequence on the target gene, and then activates transcription of promoter genes containing κB sites. 11
Treatment of diabetic wounds includes debridement, mechanical load relief, topical antibiotics, and dressings, whereas newer developments include the use of bioengineered skin equivalents, growth factor therapy, and hyperbaric oxygen treatment. The therapeutic advantages of photobiomodulation for various diseases including diabetes mellitus and inflammation have been documented. A study conducted by Al-Watban et al. 12 utilized different diode wavelengths (532, 633, 810, and 980 nm) and dosages (5, 10, 20, and 30 J/cm2) to determine the effects of wound healing on diabetic rats. They discovered that the healing of diabetic wounds was accelerated when treated with an optimum fluence of 10 J/cm2 at a wavelength of 633 nm. Rabelo et al. 13 also found laser therapy (632.8 nm; 10 J/cm2) to be beneficial in healing wounds in both streptozotocin-induced diabetic and non-diabetic rats. There was a decrease in inflammatory processes in inflammatory cells, vessels, and fibroblasts. Aimbire et al. 14 showed a reduction in acute inflammation and TNF-α levels post-irradiation at 650 nm and 0.11 J.
The aim of this study was to determine the response of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6 expression and NF-κB translocation) after 1 or 24 h incubation post-irradiation at a wavelength of 636 nm with a fluence of 5 J/cm2 in normal, normal wounded, diabetic wounded, and hypoxic human skin fibroblasts.
Materials and Methods
All cell culture products were supplied by Scientific Group Adcock Ingram, Midrand, South Africa, unless otherwise stated.
Cell culture
Fibroblasts were of the primary cells involved in wound healing. Commercially purchased normal human skin fibroblasts (WS1, ATCC® CRL-1502) were grown in 75 cm2 cell culture flasks in Minimum Essential Media (MEM; Invitrogen, 32360026). Media was supplemented with 2 mM L-glutamine (Gibco, 25030), 0.1 mM Non-Essential Amino Acid (NEAA; Gibco, 11140), 1 mM sodium pyruvate (Gibco, 11360), 1% penicillin-streptomycin (Gibco, 15140), 1% Fungizone (Amphotericin-B antifungal; Gibco, 15240-062), and 10% Fetal Bovine Serum (FBS; Gibco, 306.00301). Cultures were incubated at 37°C in 5% CO2 with 85% humidity. Cells were detached using 1 ml/25 cm2 0.25% trypsin (Gibco, 15090) - 0.03% EDTA solution.
The study consisted of four groups: normal (N), normal wounded (NW), diabetic wounded (DW), and hypoxic (H). Hyperglycemia is the key metabolic abnormality in diabetes mellitus that is believed to play the most prominent role in the development of diabetic complications. 15 A number of studies have shown that exposure of cells to a hyperglycemic environment influences cellular metabolism. Previous studies have shown that continuous exposure of fibroblast cells to a glucose concentration of 22.6 mMol/L slowed cellular migration and that there was an increase in both cellular and DNA damage and apoptosis. 16 A diabetic in vitro model was based on McDermott et al. 16 , Houreld and Abrahamse, 17 and Vinck et al. 15 , whereby cells were continuously grown in complete MEM (basal glucose concentration of 5.56 mM) containing additional 17 mM D-glucose (Sigma Aldrich, Kempton Park, South Africa D9434). Approximately 6 X 105 cells in 3 ml complete media were transferred into 35-mm culture dishes (BD Biosciences, BD 353001) and allowed to attach overnight. An in vitro wound model was performed as previously described. 18 Briefly, a wound was induced 30 min prior to irradiation by scratching the cellular monolayer with a sterile 1-ml pipette, thereby producing a cell-free zone. 19 The classical scratch method allows wounding of up to 5–10% of the surface of a confluent monolayer of cultured cells grown on a culture dish. 20
Diabetic wounds are often susceptible to hypoxia. For hypoxic cultures, 6 × 105 cells were grown in complete MEM without FBS for 24 h. 21 –25 Cultures were then incubated in an anaerobic jar with an anaerobic gas pack (Davies Diagnostics, Randburg, South Africa; Mitsubishi Gas Chemical Co., 4371051) for 4 h 26 to induce a hypoxic insult. Incubation with the gas pack results in 0% O2 and 20% CO2 within 2 h. 27
Laser irradiation
All lasers were set up by the National Laser Centre (NLC). Laser parameters are shown in Table 1. Beam power was measured using the FieldMate (Coherent, Santa Clara, CA) with sensor. The sensor was held in a position such that the reading aperture was filled. This reading (in mW) was used to determine irradiation time. Cells were irradiated with a fluence of 5 J/cm2 calculated at 476 sec (7 min and 56 sec). Cells were irradiated in the dark in 1 ml fresh media, from above with the culture dish lid off. Each model had a non-irradiated control (0 J/cm2), which was treated in the same manner as the irradiated cells, barring irradiation. Post-irradiation cells were incubated for 1 or 24 h at 37°C in 5% CO2 with 85% humidity. Changes following laser irradiation were determined by measuring cellular morphology, viability, apoptosis, proliferation, and cytokine expression.
Cellular morphology
Changes were observed using an inverted microscope (Olympus, Wirsam, Johannesburg, South Africa, CKX41). Haptotaxis (change in cell growth direction), cellular migration, and the number of cells in the central scratch were evaluated in wounded cultures.
Cellular viability
The trypan blue exclusion test was used to determine cellular viability. Briefly an equal volume of cells suspended in Hanks Balanced Salt Solution (HBSS; Gibco, 14170) and 0.4% trypan blue (Sigma Aldrich, Kempton Park, South Africa, T8154) were mixed, left at room temperature for 5 min, and loaded onto a hemocytometer. Viable and non-viable cells were counted and percentage viability was calculated by dividing the viable cell count by the total cell count and multiplying by 100. 28
Caspase 3/7
TNF-α is able to initiate apoptosis by activating caspase 8, which in turn activates caspase 3 and 7. Apoptosis was determined with the Caspase-Glo 3/7 luminescent assay (Whitehead Scientific, Brackenfell, South Africa; Promega G8091). Fifty microliters of cells were cultured in a 96-well luminescent plate (Scientific Group Adcock Ingram, Midrand, South Africa BD354651) and left to attach for 3 h at 37°C. Fifty microliters of Caspase-Glo 3/7 reagent was added and the plate was then incubated at room temperature for 2 h. Luminescence was read using the Victor 3 (Perkin–Elmer, Separation Scientific, Johannesburg, South Africa).
Cellular proliferation
Cell proliferation was determined using the XTT colorimetric assay (Roche, Isando, South Africa, 11465015001). The test is based on the cleavage of the yellow tetrazolium salt XTT to form an orange formazan dye by metabolically active cells. One hundred microliters of cells were incubated in tissue culture treated 96-well plates for 24 h at 37°C. XTT labelling reagent (50 μl/sample) was mixed with electron coupling reagent (1 μl/sample), added to the cells, and incubated for 4 h. Absorbance was read at 450 nm (Victor 3, Perkin–Elmer, Separation Scientific, Johannesburg, South Africa).
Pro-inflammatory cytokines
Cytokine expression was analyzed using ELISA sets from BD Biosciences (Scientific Group Adcock Ingram, Midrand, South Africa; Human TNF-α, 555212; IL-1β, 557953; and IL-6, 555220) and BD Biosciences optEIATM Reagent Set B (Scientific Group Adcock Ingram, Midrand, South Africa, 550534). Pro-inflammatory cytokines released from cells were measured in culture media from both irradiated and non-irradiated samples. Wells were coated with 100 μl capture antibody diluted in coating buffer and incubated at 4 °C overnight. Wells were then blocked with 200 μl assay diluent to avoid nonspecific binding and incubated at room temperature for 1 h. One hundred microliters of sample or standard were added to the appropriate wells and incubated at room temperature for 2 h. A biotin conjugated streptavidin-specific secondary antibody was added (100 μl) and incubated for 1 h at room temperature. Tetramethylbenzidine (TMB) substrate was added (100 μl) and plates were incubated for 30 min at room temperature in the dark. The reaction was stopped by adding 50 μl stop solution, and absorbance was read within 30 min at 450 nm with correction at 570 nm (Victor 3, Perkin–Elmer, Separation Scientific, Johannesburg, South Africa).
NF-κB (p50) translocation
NF-κB translocation was determined by immunofluorescence staining. Briefly, sterile cover slips were inserted in 35-mm culture dishes (Scientific Group Adcock Ingram, Midrand, South Africa; BD Biosciences, BD 353001) and cells were allowed to grow as described previously. Cells were then rinsed with phosphate buffered saline (PBS) and fixed with 3.5% formalin and washed three times with PBS. Incubation with Triton X-100 for 15 min made the cell membrane permeable so that primary and secondary antibodies could cross to the intracellular cytokine. Cells were then blocked with 0.1% bovine serum albumin (BSA) with 0.01% azide buffer diluted in PBS. Cultures were incubated for 20 min at room temperature and then washed with PBS. Primary antibody (1:250, Southern Cross Biotechnology, Johannesburg, South Africa; Santa Cruz Biotechnology Sc-53744) was prepared in 1.5% normal blocking serum (NBS) diluted in PBS. Cells were incubated in 200 μl primary antibody for 1 h at room temperature. Cells were washed with PBS, and then incubated in the dark with goat anti-mouse IgG-FITC conjugated secondary antibody (1:250, Southern Cross Biotechnology, Johannesburg, South Africa; Santa Cruz Biotechnology Sc-2010) for 45 min at room temperature. Nuclei were stained with 1 μg/ml 4’6-diamidine-2-phenylindole (DAPI, Sigma Aldrich, Kempton Park, South Africa, D9564). The cover slip with cells was mounted on a microscope slide and fluorescence was visualized on the Carl Zeiss Axio Z1 cell observer (Göttingen, Germany).
Statistical analysis
Each experiment was performed six times (n = 6), and each test was performed in duplicate, the average of which was used. SigmaPlot version 8.0 (Systat Software, San Jose, CA) was used for statistical analysis. The Student t-test was used to determine statistical significance between non-irradiated controls and their respective treated (irradiated) experimental group, and the one-way analysis of variance (ANOVA) was used to analyze the differences between the different data groups at 1 and 24 h incubation. Statistical significance was accepted at the 0.05 level (95% confidence interval). The ± value, or error bars in the figures, indicate standard error of the mean (SEM). Statistical probability is represented as p < 0.05 (*), p < 0.01 (**), and p < 0.001 (***) as compared to non-irradiated control cells (0 J/cm2).
Results
Cellular morphology
Morphologically, there was no difference between non-irradiated normal cells and non-irradiated diabetic wounded cells (Fig. 1a, b, g, and h, respectively). On the other hand, non-irradiated hypoxic cells were less directional and had lost their characteristic long elongated spindle shape, cells appeared star shaped with gaps between them (Fig. 1j and k). Non-viable cells could also be seen floating in the media. Irradiated normal cells showed no significant change in morphology post-irradiation. Cells remained elongated (Fig. 1c). In both normal and diabetic wounded cultures, a clear ‘wound’ could be seen 30 min after wound induction (Fig. 1d and g, respectively); few cells were seen in the central scratch after incubation for 24 h (Fig. 1e and h, respectively). However, 24 h post-irradiation cells changed their direction of growth (haptotaxis) and had migrated into the central scratch at a higher rate than non-irradiated cells (Fig. 1f and i, respectively). More cells were seen in the central scratch in irradiated cultures than non-irradiated cultures. Twenty-four hours post-irradiation, hypoxic cells regained their typical morphology (Fig. 1l); spaces were still observed between cells.

Morphology of normal (N), normal wounded (NW), diabetic wounded (DW), and hypoxic (H) WS1 fibroblasts irradiated with 5 J/cm2 (636 nm) and left to incubate for 24 h. N, unstressed fibroblast cells showed no changes in cellular morphology after 24 h incubation (
Cellular viability
Cellular viability was determined by the trypan blue exclusion test (Table 2). A significant increase in percentage viability was seen at both 1 and 24 h in irradiated hypoxic cells (p < 0.01 and p < 0.001, respectively), compared to non-irradiated control cells. Diabetic wounded cells showed an increase at 24 h (p < 0.01). Comparison of incubation times revealed a significant increase in viability in irradiated normal wounded and hypoxic cells (p < 0.05 and p < 0.01, respectively) when incubated for 24 h, whereas non-irradiated cells showed significances in normal and normal wounded cells (p < 0.05 and p < 0.01, respectively).
Significant differences between irradiated and non-irradiated controls are shown as p < 0.01 (**) and p < 0.001 (***). DW cells showed a significant increase in viability 24 h post-irradiation, whereas H cells showed significant increases at both 1 and 24 h. NW and DW cells showed a decrease in caspase 3/7 activity 1 h post-irradiation, whereas NW and H cells showed a decrease at 24 h.
Standard error of the mean (SEM).
Caspase 3/7
Apoptosis was determined by measuring caspase 3/7 activity (Table 2). After 1 h incubation, irradiated normal wounded and diabetic wounded cells showed a significant decrease in caspase 3/7 activity compared to non-irradiated control cells (p < 0.01). When cells were left to incubate for 24 h, a significant decrease was seen in normal wounded and hypoxic cells (p < 0.01). With the exception of non-irradiated hypoxic cells, all cells showed a significant decrease in caspase 3/7 activity 24 h post-irradiation compared to the same cells incubated for 1 h (p < 0.01). All three of the stressed models, namely normal wounded, diabetic wounded, and hypoxic, showed a significant increase in apoptosis compared to normal (unstressed) cells. This was seen in both non-irradiated (p < 0.001) and irradiated cells (p < 0.01) at both incubation times.
Cellular proliferation
The XTT colorimetric assay was used to determine cellular proliferation in irradiated cells (Fig. 2). Compared to each non-irradiated control, normal wounded, diabetic wounded, and hypoxic cells showed a significant increase in proliferation when left to incubate for 1 h (p < 0.001) and 24 h (p < 0.01). Comparison of the groups with 1 h and 24 h incubation times revealed a significant decrease in proliferation at 24 h compared to 1 h in non-irradiated normal wounded cells (p < 0.05). Non-irradiated normal cells showed a significant increase in proliferation compared to non-irradiated normal wounded, diabetic wounded, and hypoxic cells at 1 h (p < 0.001). At 24 h this increase was only seen compared to diabetic wounded and hypoxic cells (p < 0.01 and p < 0.001, respectively). When cells were irradiated and incubated for 1 h, a significant increase in proliferation was seen in normal wounded cells compared to in irradiated normal cells, whereas in hypoxic cells this significance was decreased (p < 0.01). At 24 h the decrease was only significant in hypoxic cells (p < 0.05).
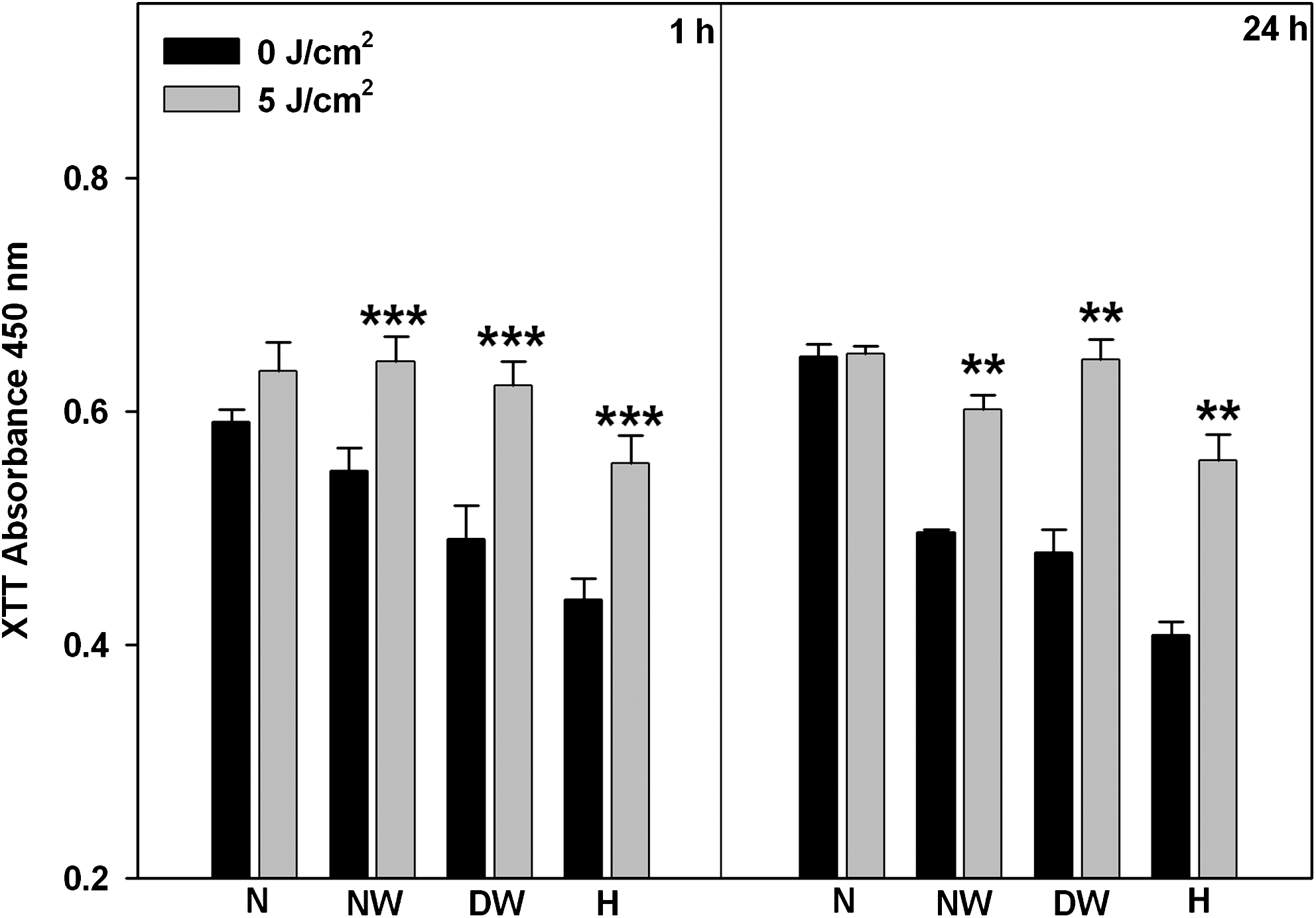
Proliferation in cells was determined 1 or 24 h post-irradiation using the XTT proliferation assay in normal (N), normal wounded (NW), diabetic wounded (DW) and hypoxic (H) WS1 cells irradiated with a diode laser (636 nm). Significant differences between irradiated and non-irradiated controls are shown as p < 0.01 (**) and p < 0.001 (***). NW, DW, and H cells showed a significant increase post-irradiation at both 1 and 24 h.
Pro-inflammatory cytokines
Pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6) were determined in cells 1 or 24 h post-irradiation by ELISA. TNF-α results are shown in Fig. 3. After 1 h incubation, normal wounded cells showed a significant decrease in TNF-α (p = 0.01), whereas hypoxic cells showed a decrease at 24 h (p < 0.05) as compared to their respective non-irradiated controls. Overall, there was a decrease in TNF-α levels when cells were incubated for 24 h compared to 1 h. Non-irradiated normal, normal wounded, and hypoxic cells showed a significant decrease at 24 h (p < 0.001, p < 0.05 and p < 0.001, respectively). The same decrease was seen in irradiated normal, normal wounded, and hypoxic cells (p < 0.01, p < 0.05, and p < 0.001, respectively). At 1 h, both non-irradiated and irradiated normal cells showed a significant decrease in TNF-α expression compared to non-irradiated and irradiated hypoxic cells respectively (p < 0.001 and p < 0.01, respectively).
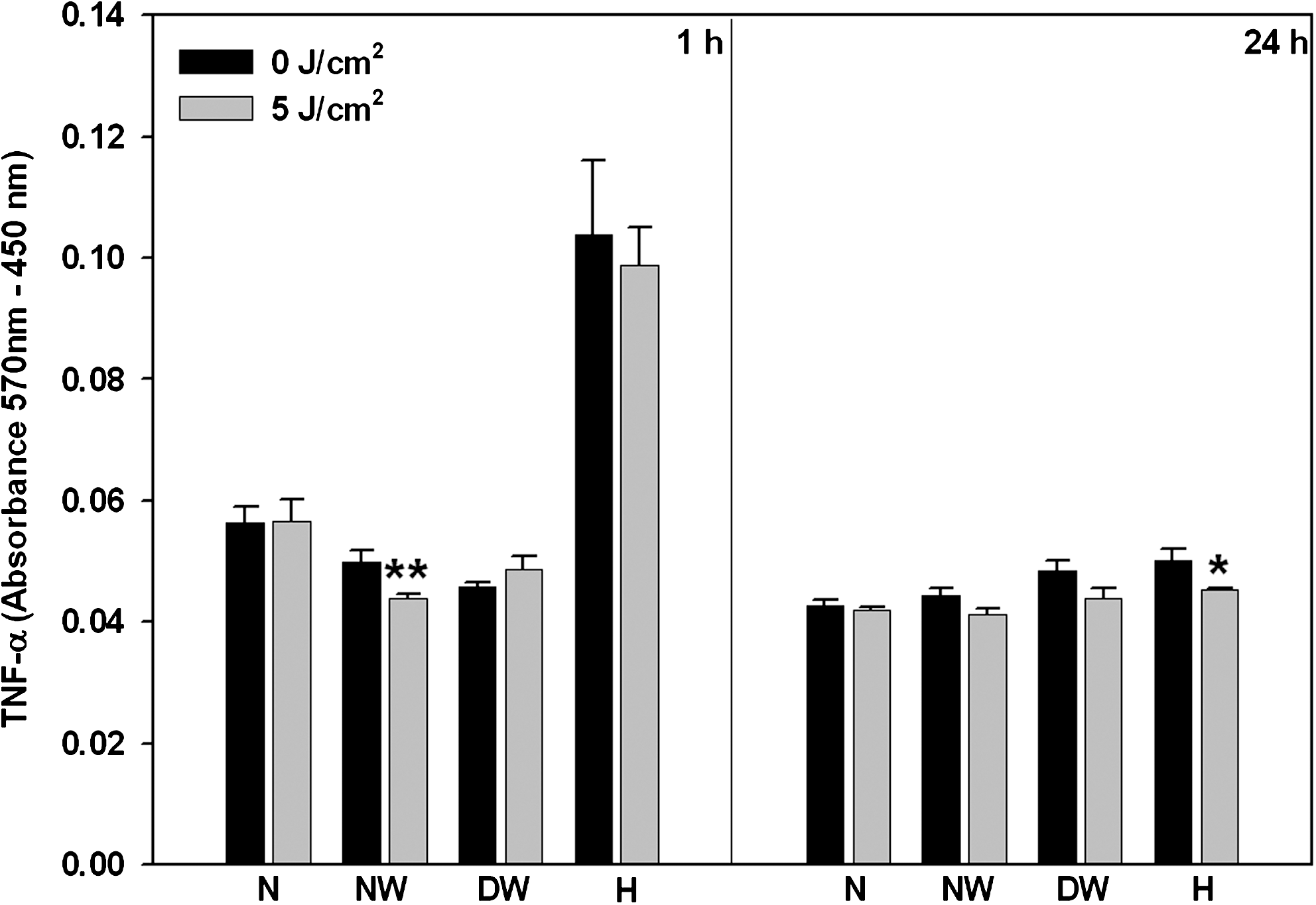
TNF-α was determined by ELISA after 1 or 24 h incubation in laser treated (636 nm) and untreated normal (N), normal wounded (NW), diabetic wounded (DW), and hypoxic (H) cells. Significant differences between irradiated and non-irradiated controls are shown as p < 0.05 (*) and p < 0.01 (**). After 1 h incubation, NW cells showed a significant decrease, whereas at 24 h, H cells showed a significant decrease.
One hour post-irradiation, IL-1β levels were significantly decreased in irradiated normal and diabetic wounded cells (p < 0.01 and p < 0.001, respectively) as compared to their non-irradiated controls (Fig. 4). When incubated for 24 h, there was a significant decrease in normal, normal wounded and diabetic wounded cells (p < 0.01). Comparison of the different times showed a significant decrease after 24 h incubation in non-irradiated normal, diabetic wounded, and hypoxic cells (p < 0.001) as compared to the same cells incubated for 1 h. At 1 h, both non-irradiated and irradiated normal cells showed a significant decrease in IL-1β compared to non-irradiated and irradiated hypoxic cells respectively (p < 0.01).

IL-1β was measured by ELISA after 1 or 24 h incubation post-irradiation (636 nm) in normal (N), normal wounded (NW), diabetic wounded (DW), and hypoxic (H) WS1 fibroblasts. Significant differences between irradiated and non-irradiated controls are shown as p < 0.01 (**) and p < 0.001 (***). N and DW cells showed a significant decrease at both 1 and 24 h, whereas NW cells showed a significant decrease at 24 h.
The only significant change seen in IL-6 compared to non-irradiated controls was seen in irradiated hypoxic cells incubated for 24 h (p < 0.001), (Fig. 5). Comparison of incubation times showed a significant decrease in IL-6 levels at 24 h in non-irradiated normal wounded and hypoxic cells (p < 0.01 and p < 0.05, respectively). At 1 and 24 h, both non-irradiated and irradiated normal cells showed a significant increase in IL-6 expression compared to non-irradiated and irradiated hypoxic cells respectively (p < 0.01).
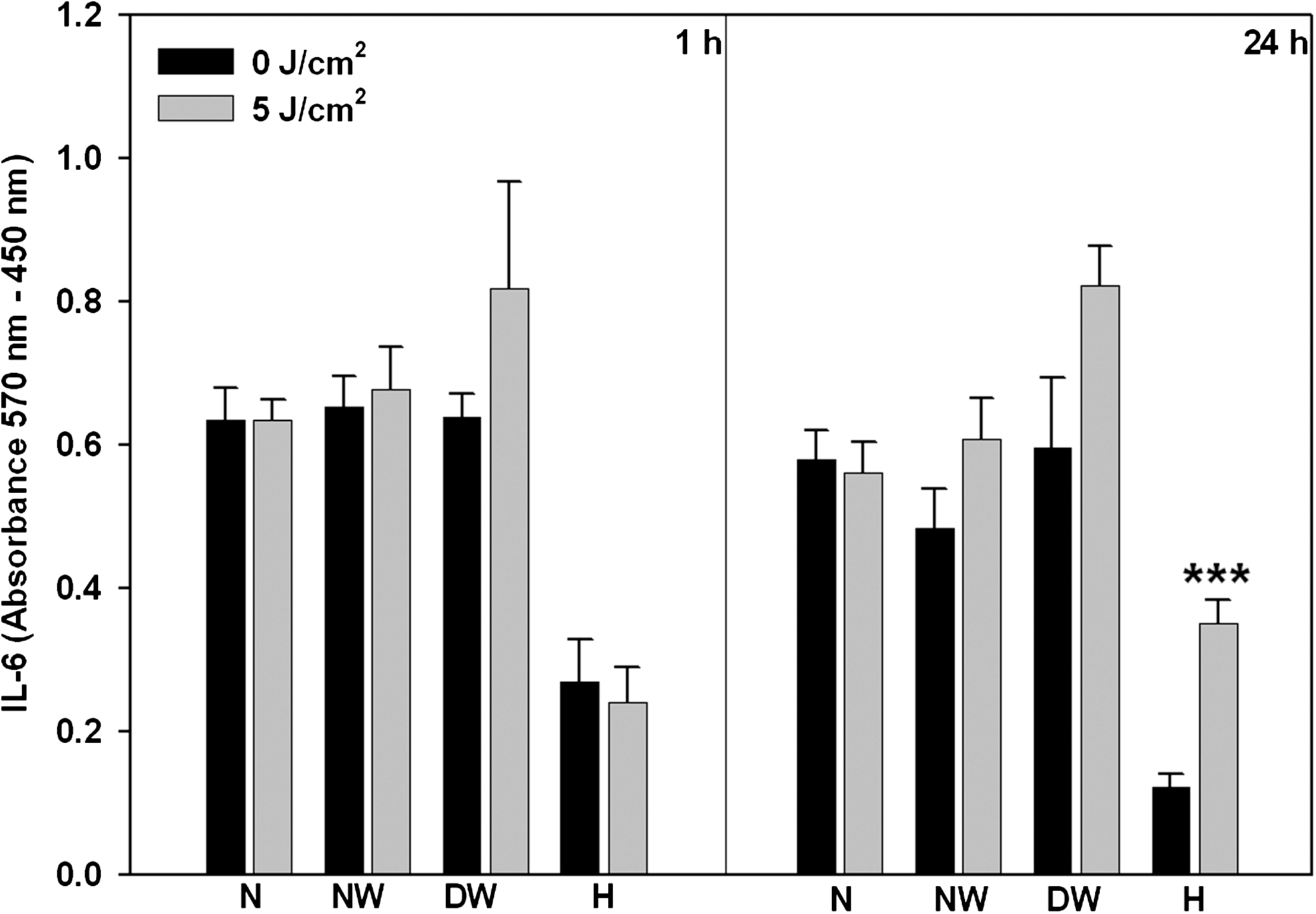
IL-6 expression was determined in normal (N), normal wounded (NW), diabetic wounded (DW), and hypoxic (H) cells at 1 and 24 h post-irradiation (636 nm) by ELISA. Significant differences between irradiated and non-irradiated controls are shown as p < 0.001 (***). Irradiated H cells showed a significant increase after 24 h incubation.
NF-κB (p50) translocation
After 1 h incubation, irrespective of irradiation, normal, normal wounded, and diabetic wounded cells showed translocation of NF-κB to the nuclei with sparse cytoplasmic fluorescence (Fig. 6a–c and e–g). Hypoxic cells showed NF-κB in the cytoplasmic and nuclear region, with abundance in the cytoplasm (Fig. 6d). However, after irradiation, less NF-κB was seen in the cytoplasm (Fig. 6h). After 24 h incubation, NF-κB was more evident. In both irradiated and non-irradiated normal, normal wounded, and diabetic wounded cells a similar pattern was seen, with little or no fluorescence in the cytoplasm (Fig. 7a–c and e–g). Hypoxic cells showed little NF-κB translocation into the nucleus, with some fluorescence seen in the cytoplasm (Fig. 7d). Irradiated hypoxic cells showed less cytoplasmic fluorescence than did non-irradiated hypoxic cells (Fig. 7h).

NF-κB (p50) translocation was determined in WS1 cells 1 h post-irradiation (5 J/cm2, 636 nm) by immunofluorescence. NF-κB can be seen predominantly in the nuclei of normal, normal wounded, and diabetic wounded non-irradiated and irradiated cells (

NF-κB (p50) translocation was determined in WS1 cells 24 h post-irradiation (5 J/cm2, 636 nm) by immunofluorescence. NF-κB can be seen in the nuclei in both non-irradiated and irradiated normal, normal wounded, and diabetic wounded cells (
Discussion
Wound healing is complex and more extensive in diabetic wounds, because of impaired cellular processes. Post-inflammation, cells such as fibroblasts begin to migrate to the wounded area with intent to form scar tissue by producing collagen and forming an extracellular matrix. In diabetes, there are various “mechanical” impairments that result in delayed healing. These include retardation of closure, delayed contraction, effects on granulocytes, defects in chemotaxis, and interference with collagen synthesis. 29 Diabetic patients require complex and continuous medical treatment, and every effort should be made to prevent or delay adverse outcomes and lower the incidence of amputation. Laser therapy has demonstrated an ability to increase proliferation and reduce inflammation, and it accelerates wound healing. 30 –33
This study used a wounded, diabetic wounded, and hypoxic cell model to determine the effect of laser irradiation (636 nm at 5 J/cm2) on proliferation, viability, and pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6). The models chosen for this study proved sufficient to study the effects of LILI in diabetic conditions in vitro and produced measurable differences between the models. Growing human skin fibroblast cells (WS1) in a hyperglycemic environment had no effect on cellular morphology; cells remained elongated and spindle shaped. There was also little effect on cellular migration. Cells were still able to migrate toward the central scratch, however, at a slower rate than normal wounded cultures. This is in contrast with the work conducted by Hamuro et al. 34 who found that 10 or 30 mM glucose inhibited cellular migration, whereas McDermott 16 found an increase in cells grown in 17.5 mM glucose. Hypoxic cells showed great variation in morphology as compared to normal cells. Cells appeared star shaped, and there were a number of gaps between cells, which was probably caused by a lack of nutrients and oxygen. This loss in normal morphology could be attributed to lactic acid production or accumulation and a decrease in ATP, which causes a switch from the oxidative to the glycolytic pathway leading to increased glucose utilization. 35 This hypothesis is supported by the change in color of the media from red to orange, suggesting a decrease in pH. Ricci et al. 36 showed that stressed endothelial cells (grown in serum-starved media) became star shaped, as opposed to cells grown in serum- supplemented media (10% FBS) which appeared round. Furthermore, stressed cells (normal wounded, diabetic wounded, and hypoxic) showed an increase in apoptosis and a decrease in proliferation compared to normal cells.
Irradiation of cells with a visible wavelength of 636 nm at a fluence of 5 J/cm2 improved migration of wounded cells, with almost complete closure or coverage of the central scratch 24 h post-irradiation. Because diabetic wounds have an impaired migration rate, fibronectin, and growth factor synthesis 37 , migration in diabetic wounded cells was not as rapid as in normal wounded cells after 1 h incubation. However, 24 h post-irradiation, diabetic wounded cells showed a marked reduction in wound area and hastened cell infiltration toward the central scratch, displaying both haptotaxis and migration. Both non-irradiated normal wounded and diabetic wounded controls showed a slower rate of wound closure and cell infiltration. This increase in migration in wounded models corresponds with Houreld and Abrahamse 17 , Hawkins and Abrahamse 18 , and Hopkins et al. 38 who also showed a time-dependent decrease in wound area. Irradiated hypoxic cells regained their characteristic fibroblast shape 24 h post-irradiation.
Hawkins and Abrahamse 18 deduced that laser irradiation stimulates mitochondrial activities that lead to normalization of cell function. This study showed a significant increase in viability and proliferation in hypoxic cells at both 1 and 24 h post-irradiation, and a decrease in caspase 3/7 activity at 24 h. Normal wounded cells also showed an increase in proliferation and decrease in apoptosis 1 and 24 h post-irradiation, whereas diabetic wounded cells showed an increase in viability at 24 h, an increase in proliferation at both 1 and 24 h, and a decrease in apoptosis 1 h post-irradiation. Therefore, LILI at a wavelength of 636 nm and 5 J/cm2 leads to normalization of cellular function and morphology, possibly leading to an increase in ATP levels and a decrease in lactic acid 39 , and also has a cytoprotective effect on cells. Carnevalli et al. 40 also found that irradiation of CHO K-1 cells with infrared light (830 nm; 10 mW; 2 J/cm2) prevented apoptosis.
In wound healing, pro-inflammatory cytokines play a role as mediators. IL-1β and TNF-α are potent inducers of inflammation. 35 TNF-α is associated with apoptosis via death domain receptor binding and activating caspase 8, which in turn activates caspase 3 and 7, cell survival mechanisms, 41 –43 and the pathogenesis and progression of gestational diabetes mellitus. Cytokines IL-1β, IL-6, and TNF-α have the potential of playing an advantageous or disadvantageous role in the cell depending upon how they are regulated or the pathway they follow. Synergism between IL-1β and TNF is a common occurrence, and they are both elevated in inflammatory states. 44 Hypoxia is a potent inducer of IL-1β. Yu and Lau 45 experienced a sharp increase in IL-1, IL-6, TNF-α, and IFN-γ from astrocytes exposed to mechanical and metabolic stress.
In the current study, non-irradiated hypoxic cells showed a significant increase in TNF-α and IL-1β levels 1 h after sham irradiation compared to normal cells. It is safe to assume that these cells are entering the apoptotic TNF-α mediated death pathway, as is supported by the decrease in viability and increase in apoptosis. Nonetheless, this significant increase is no longer evident at 24 h, and is likely caused by growing cells in a normoxic environment after sham irradiation. Irradiation of hypoxic cells produces a further decrease in TNF-α, as is shown by the significant decrease 24 h post-irradiation. This again leads to the fact that laser irradiation at the correct fluence and wavelength has the potential to shift cells from a death pathway into a cell survival pathway. This notion is further supported by the increase in viability and proliferation, and decrease in apoptosis in irradiated cells. Therefore, anti-inflammatory effects were initiated in hypoxic cultures. Safavi and colleagues 46 noted a decrease in inflammatory mediators (IL-1β, TNF-α, and IFN-γ) in wounded rats (7 mm incision on the gingival and mucosal tissues) measured 24 h post-irradiation (632.8 nm; 7.5 J/cm2). There was no significant change in TNF-α in diabetic cells. Perhaps the stress of growing cells in additional 17 mMol/l glucose was not enough to induce an increase in this pro-inflammatory cytokine. Irradiated diabetic wounded cells showed a significant decrease in IL-1β at both 1 and 24 h compared to non-irradiated control cells. Irradiated normal cells showed the same decrease in IL-1β. Irradiated normal wounded cells showed a significant decrease in TNF-α and IL-1β at 1 or 24 h, respectively.
IL-6 is a pleitropic cytokine that has both pro- and anti-inflammatory effects. 47 In skin it is produced by fibroblasts, endothelial cells, and other tissues or cells and is induced during acute phase reactions and usually expressed in response to or together with IL-1 and TNF-α. 48 In diabetic patients these cytokines are known to be present in high concentrations. 6,49 Care should be taken when interpreting IL-6, as this cytokine is also used as a measure of proliferation 17 , and should not be performed on its own, as an increase could mean an increase in proliferation, or an increase in inflammation. In this study, the decrease in IL-6 levels at 1 and 24 h in non-irradiated and irradiated hypoxic cells compared to normal cells is an indication of the decrease in proliferation, as is supported by the XTT results. The elevation of IL-6 in irradiated hypoxic cells is indicative of the anti-inflammatory effects induced by IL-6. This is supported by the decrease in apoptosis and TNF-α, and the increase in viability and proliferation.
In non-stimulated cells, NF-κB complexes are trapped in the cytoplasm bound to the inhibitory IκB proteins. External or internal stimulation, such as hypoxia, phosphorylates IκB proteins, which are then ubiquitinated and broken down in the proteasomes. Consequently, the NF-κB complex is translocated to the nucleus where it activates the transcription of numerous genes or responses, i.e., differentiation, apoptosis, oxidative stress, and inflammatory genes. 50 –52 NF-κB can be activated by many stimuli including IL-1 and TNF-α. 53
NF-κB in normal and normal wounded cells, despite irradiation and incubation time, was seen in the nuclear region. It could be expected that NF-κB will be seen abundantly in the cytoplasm, as cells were not stimulated. Nonetheless, as these cells exhibited high proliferation and viability as well as decreased caspase 3/7, IL-1β, and TNF-α, it can be assumed that translocation is caused by increased proliferation and viability, and that cells are not apoptotic. The same could be said for diabetic wounded cells, as they were not stressed compared to normal or normal wounded cells. Hypoxic cells were severely stressed, as was seen by elevated caspase 3/7, IL-1β, and TNF-α, especially after 1 h incubation. Therefore, less NF-κB fluorescence was seen in the nuclei in non-irradiated cells, suggesting that survival proteins were not yet activated. However, after irradiation, more fluorescence was noted in the nuclear region, which could have meant that cellular functions had been restored. This hypothesis is supported by the increase in viability and proliferation and the decease in apoptosis (caspase 3/7). When cells were left to incubate for 24 h, non-irradiated hypoxic cells showed less NF-κB in the cytoplasm than did the same cells incubated for 1 h. This could be because cells were no longer grown under stress conditions, i.e. hypoxia, but were instead grown under normoxic conditions.
Conclusions
This study showed that the models used were sufficient to study the effect of LILI in vitro, and they produced measurable differences. 54 The models also showed varying degrees of stress, with hypoxic cells showing the most stress and recovery post-irradiation. Irradiation of diabetic wounded cells to a wavelength of 636 nm at a fluence of 5 J/cm2 showed hastened migration of cells, an increase in viability and proliferation, and a decrease in caspase 3/7 activity (apoptosis) and IL-1β. Irradiated normal wounded cells showed hastened migration, increased proliferation, and a decrease in apoptosis and pro-inflammatory cytokines TNF-α and IL-1β. Irradiated hypoxic cells showed an improvement in morphology, an increase in viability, proliferation, IL-6, and a decrease in apoptosis and TNF-α.
Overall, this study showed that irradiation at a wavelength of 636 nm with 5 J/cm2 has a positive effect on cellular morphology and function, increases cellular proliferation, and has a cytoprotective effect on stressed diabetic wounded and hypoxic cells in vitro. It has an anti-inflammatory effect and is able to direct stressed cells into the cell survival pathway. The results show that phototherapy may be a useful modality for wound healing in diabetes. 54
Footnotes
Acknowledgments
This study was funded by the University of Johannesburg, National Research Foundation South Africa and Council for Scientific and Industrial Research. Lasers were supplied by the National Laser Center South Africa.
Author Disclosure Statement
No competing financial interests exist.