
Abstract
Introduction
Irradiation of cells with red or infrared laser light can stimulate gene expression and release of growth factors from cells in culture. 9 Experiments in which supernatants of macrophage-like cells irradiated with a 820 nm laser diode were transferred to fibroblasts also growing in culture demonstrated for the first time laser-induced secretion of growth factors resulting in increased fibroblast proliferation. 10 The laser photostimulation of wound healing in splinted wounds of diabetic mice described previously 11,12 could be the result of restoring the levels of certain growth factors in these wounds. It would be helpful to know which deficiencies of growth factors in diabetic wounds might be improved by laser irradiation, and also to identify those that are not likely to be increased by laser therapy. Application of the latter to wounds in combination with laser irradiation may provide a way of maximizing the healing of diabetic wounds.
The expression of growth factors and cytokines in the wounds of nondiabetic and diabetic animals reflects in part the cellular elements present in the wounds at different stages of healing. One approach to designing a protocol of combination therapy for diabetic wounds is to first identify those growth factors that are reduced in the wounds of diabetic animals. This could be the result of a delayed recruitment or inhibition of various cell types in the wounds. The gene expression and release of certain growth factors and cytokines by specific cell types involved in wound healing in response to laser irradiation have been described previously. 9 By this means, it may be possible to identify those growth factors or cytokines whose expression is lowered in wounds of diabetic animals and which might not be increased by laser irradiation. Such growth factors could be applied to diabetic wounds in combination with laser irradiation to further enhance healing.
It seemed pertinent to review studies examining the gene expression and release of growth factors during cutaneous wound healing of nondiabetic and diabetic animals to identify deficiencies in diabetic animals. Important processes that occur in wound healing are: (1) formation of a platelet- and fibrin-containing plug at the wound site; (2) invasion of the wound by neutrophils and monocytes/macrophages; (3) proliferation and migration of keratinocytes from the wound edge to begin re-epithelization of the wound; (4) proliferation of fibroblasts at the dermal edge; (5) formation of granulation tissue comprising fibroblasts, macrophages, lymphocytes, plasma cells, and an extracellular matrix containing collagen fibers and macromolecular components such as glycoproteins, and the formation of new blood vessels; and (6) maturation of granulation tissue and collagen fibers, and vascularization of wound tissue. The processes involving inflammation, re-epithelization, granulation tissue and extracellular matrix formation, angiogenesis, and tissue remodeling are controlled by growth factors and cytokines produced by the cellular elements present in the wounds and also contained in the blood. Cytokines regulate inflammatory cells during the early stages of wound healing and also influence the gene expression and release of growth factors that are important in the proliferation and differentiation of cells and the synthesis of an extracellular matrix. The presence of receptors or binding proteins for specific growth factors on cells is also important in determining cellular responses.
This article critically reviews those articles published up to September 2011. It focuses on experimental studies that have investigated the gene expression and release of growth factors in the cutaneous wounds of nondiabetic and diabetic animals. These studies have been performed mostly in mice and rats, and only studies with these two animals have been included. The aim of this article was to identify deficiencies of growth factors in the wounds of diabetic animals, and restoration of the levels of some which might be achieved by laser irradiation. Those growth factors whose levels might not be restored by laser irradiation at the appropriate stage of healing could be applied to the wound as part of a combined therapy with laser light.
Methods
Original research articles investigating growth factor expression in skin wounds of nondiabetic and diabetic mice and rats and published up to September 2011 were retrieved and used for this review. Relevant articles were sought and obtained from library sources and the online database PubMed using EndNote XV (Thomson Reuters, Carlsbad). Search terms were
We included studies that met the following criteria: 1. Studies involving skin wounds 2. Studies performed in nondiabetic (normal) and/or diabetic mice or rats 3. Studies examining gene expression and/or release of growth factors or cytokines from wounds 4. At least one growth factor or cytokine identified as the dependent variable
Studies excluded from this study were: 1. Studies performed on animals other than mouse or rat 2. Studies involving wounds in tissues other than skin (e.g., cornea, gingiva) 3. Reviews and meta-analyses 4. Studies for which only an abstract was available 5. Studies reported in languages for which no English language translation was available
Results
In vivo studies
Results from the literature search are summarized in Fig. 1. There were 23 searched studies on growth factors in nondiabetic and diabetic wounds (n=15, mouse; n=8, rat). 13 –35 A variety of acute wound models was used in these studies (burn ulcer, incisional, excisional, subcutaneous PVA sponge, and stainless steel mesh chamber). Further details regarding these studies are given in Tables 1 –5. One of the studies also examined the effect of serum growth factors on VEGF mRNA expression by human keratinocytes in vitro ,31 and this has been included in Table 5.
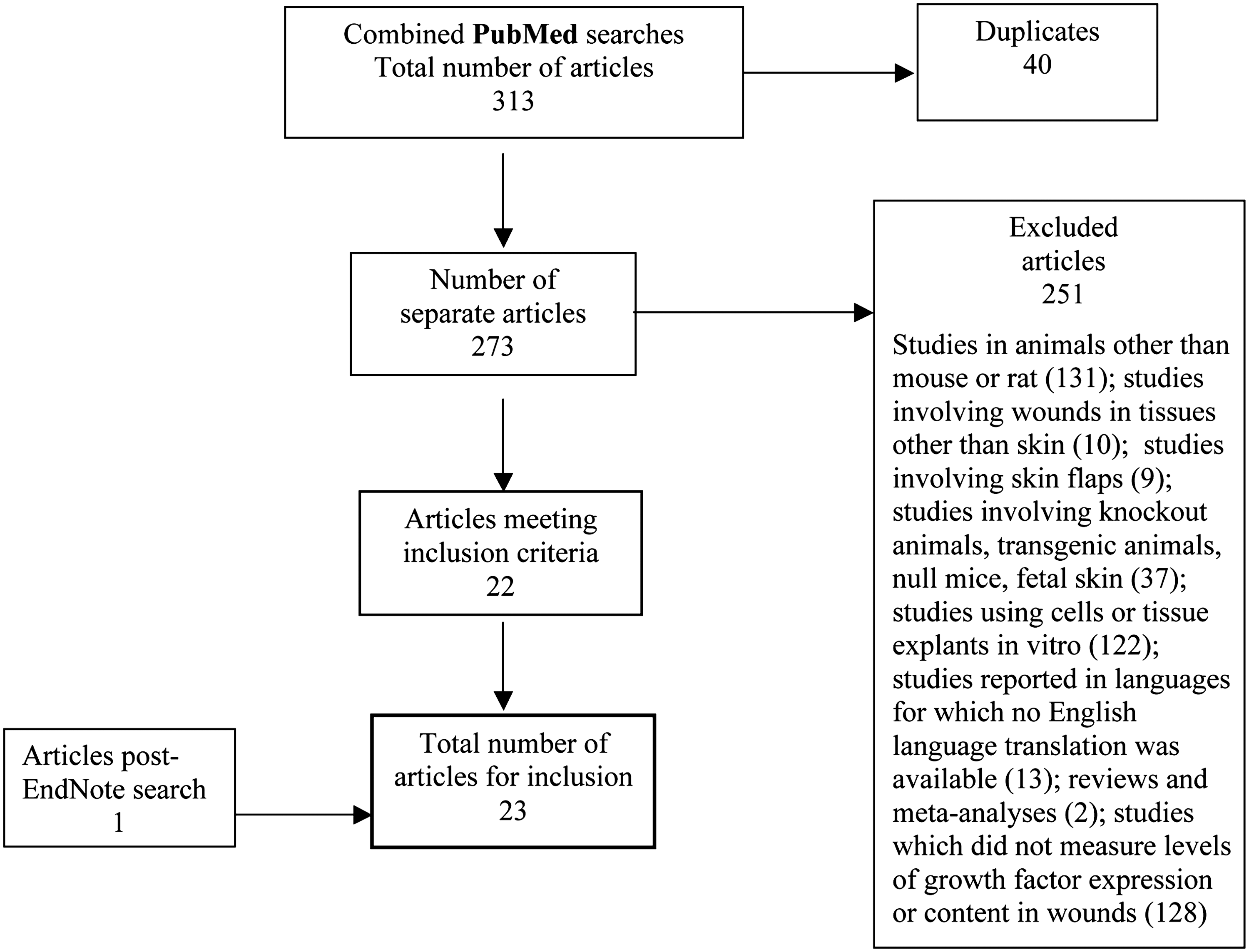
Flowchart of literature search: studies of gene expression of growth factors in skin wounds of nondiabetic and diabetic mice and rats.
STZ, streptozotocin; FGF, fibroblast growth factor; TGF-β, transforming growth factor beta.
IL, interleukin; TNF-α, tumor necrosis factor-α; TGF-β, transforming growth factor beta; VEGF, vascular endothelial growth factor; PCR, polymerase chain reaction; IGF, insulin-like growth factor.
HIF, hypoxia-inducible factor; PlGF, placental growth factor; PDGF, platelet-derived growth factor; VEGF, vascular endothelial growth factor; PCR, polymerase chain reaction; STZ, streptozotocin; FGF, fibroblast growth factor; TGF, transforming growth factor; HGF, hepatocyte growth factor; NGF, nerve growth factor; PBS, phosphate-buffered saline; EIA, enzyme immunoassay; IGF, insulin-like growth factor; KGF, keratinocyte growth factor.
STZ, streptozotocin; IGF, insulin-like growth factor; TGF, transforming growth factor.
STZ, streptozotocin; FGF, fibroblast growth factor; TGF, transforming growth factor; HIF, hypoxia-inducible factor; PlGF, placental growth factor; PDGF, platelet-derived growth factor; VEGF, vascular endothelial growth factor ; PCR, ppolymerase chain reaction; PBS, phosphate-buffered saline; NGF, nerve growth factor; CTGF, connective tissue growth factor; EIA, enzyme immunoassay; DMEM, Dulbecco's Modified Eagle Medium; FBS, fetal bovine serum; EGF, epidermal growth factor; KGF, keratinocyte growth factor; PMN, polymorphonuclear leukocytes; TNF-α, tumor necrosis factor α; IL, interleukin.
Burn ulcer wound model (Table 1)
In the rat, one study used nondiabetic and diabetic animals, which served as an experimental model of type 1 diabetes. Expression amounts of TGF-β cell proliferation proteins in diabetic animals on day 7 after establishing the ulcer were significantly reduced compared with nondiabetic animals. Expression amounts of TGF-β proliferation proteins were increased for diabetic animals on day 14 compared with day 7, whereas for nondiabetic animals the amounts were decreased on day 14 compared with day 7.
Incisional wound model (Table 2)
In the mouse, all five studies used nondiabetic and diabetic animals, which served as an experimental model of type 2 diabetes. Two full-thickness longitudinal incisions were made in the dorsum and closed with surgical sutures or skin clips. These wounds would have healed by first intention. A marked induction of VEGF mRNA was found at days 3 and 6 in the wounds of nondiabetic animals compared with basal values, and thereafter declined at day 12 after wounding. VEGF mRNA was markedly lower in the wounds of diabetic animals at days 3 and 6 compared with nondiabetic animals, and low levels were still detectable at day 12. VEGF content in the wounds of nondiabetic animals was raised on day 6 after wounding. VEGF content was markedly lower in the wounds of diabetic animals at days 3, 6, and 12 compared with nondiabetic animals. Therefore, the expression of mature VEGF protein in skin wounds of diabetic animals was significantly less than for nondiabetic animals, and did not show the maximum increase on day 6 that had been found for nondiabetic animals.
In the rat, all three studies used nondiabetic animals only. In two of these studies, a full-thickness single linear incision was made in the dorsum, and in one study the wounds were not sutured or covered, whereas in the other study the wounds were closed with surgical clips. In the former study, the wounds healed by second intention, which is the process by which excisional wounds heal, and VEGF mRNA expression in the wounds was significantly increased 1 day after wounding compared with 3 and 7 days after wounding. VEGF protein content in the wounds was significantly higher at 1 and 3 days after wounding compared with the unwounded skin of nondiabetic animals. Therefore, mRNA expression and protein content of VEGF were high at day 1 for wounds that were not sutured.
In the latter study, the wounds healed by first intention. IGF-1 mRNA expression was low in unwounded skin and at 0.5 day after wounding. The level in the wound started to increase at 1 day and peaked at 10 days after wounding. The expression remained at a high level through to 21 days after wounding. IGF-2 mRNA expression was low in unwounded skin and at 0.5 day after wounding. The level in the wound increased during the early stages of wound healing and reached a peak at 10 days after wounding. The expression then decreased, although the levels at 15 and 21 days were still significantly higher than for unwounded skin and at 0.5 day after wounding. Therefore, mRNA expression for both IGF-1 and IGF-2 was maximal at 10 days for wounds that were sutured.
Excisional wound model (Table 3)
In the mouse, all except one of the nine studies used nondiabetic and diabetic animals. Four of the studies used an experimental model of type 1 diabetes, and four used an experimental model of type 2 diabetes. In the study with nondiabetic animals only, four full-thickness wounds were made on the dorsum and not covered, and healing would have occurred by second intention. Expression of mRNA for FGF-7 (KGF-1), FGF-10 (KGF-2) and TGF-β was increased on days 2 and 15 as compared with the levels on day 0.
For the eight studies using nondiabetic and diabetic animals, two studies made a single full-thickness wound on the dorsum and in one of these studies the wounds were covered. In the study in which a single circular wound was made, mRNA induction of placental growth factor (PlGF) in wounds of nondiabetic animals began on day 1 and reached a maximum on day 3 after wounding. In wounds of diabetic animals, PlGF mRNA induction was significantly reduced compared with nondiabetic animals on days 1, 3, and 5 after wounding. PlGF protein induction in wounds of nondiabetic animals began to increase on day 1, reached a maximum on day 3, and remained upregulated until day 9 after wounding. In wounds of diabetic animals, PlGF protein induction was significantly reduced compared with nondiabetic animals from days 1 to 7 after wounding. VEGF mRNA expression was induced in wounds of nondiabetic animals and peaked on day 3 after wounding. In wounds of diabetic animals, VEGF mRNA expression was reduced compared with nondiabetic animals, with a marked lowering on days 3 and 5 after wounding. Induction of PDGF-B mRNA, FGF-2 mRNA, and TGF-β1 mRNA was strongly decreased in wounds of diabetic animals compared with nondiabetic animals. In the study in which a single square wound was made and covered with a transparent dressing, IGF-1 mRNA was induced in wounds of nondiabetic animals at 12 h and at 3 days after wounding there was a 50-fold increase compared with the level at 12 h, with a gradual decrease occurring to 28 days after wounding In wounds of diabetic animals, IGF-1 mRNA levels rose minimally at 5 days and did not reach a maximum until 14 days after wounding, with the levels being twofold lower than those of the peak at 3 days in the nondiabetic animals. A sharp increase in IGF-1 protein concentration occurred at 5 days in the nondiabetic wounds, which declined slowly thereafter to 28 days after wounding. IGF-1 protein levels in the diabetic wounds remained low until 21 days after wounding when they rose to nondiabetic levels. IGF-2 mRNA in wounds of nondiabetic animals peaked at 3 days after wounding, whereas in wounds of diabetic animals peak levels occurred at 10 days after wounding and were fourfold greater than in nondiabetic wounds. In the other six studies, multiple wounds were made on the dorsum of each animal. In the study in which two circular wounds were made in the interscapular region, nerve growth factor (NGF) in wounds of diabetic animals was significantly lower than in wounds of nondiabetic animals on day 3 after wounding. Expression of VEGF-A mRNA was significantly higher in wounds of diabetic animals than in wounds of nondiabetic animals on day 3 after wounding. In the other studies four or six circular wounds were made on the dorsum. One of these studies showed a lack of induction of VEGF mRNA in wounds of diabetic animals within the first few hours of wounding, and expression was markedly lower and barely detectable at day 5 compared with wounds of nondiabetic animals, for which highest levels occurred between days 3 and 7 after wounding. Another study found a delay in KGF induction in wounds of diabetic animals, with maximal levels of KGF expression only obtained between 3 and 5 days after wounding, whereas expression of KGF mRNA in wounds of nondiabetic animals reached a peak at 24 h after wounding. In addition, induction of both FGF-1 mRNA and FGF-2 mRNA occurred earlier in wounds of diabetic animals compared with nondiabetic animals, and expression of both FGF-1 mRNA and FGF-2 mRNA returned to basal levels within 3 days after wounding. In contrast, enhanced expression of FGF-1 mRNA and FGF-2 mRNA in wounds of nondiabetic animals occurred during the first 7 days after wounding. FGF-2 mRNA expression in wounds of diabetic animals was induced 3.5-fold and peaked within 24 h after wounding. In wounds of nondiabetic animals, FGF-2 mRNA expression was induced fourfold with the peak being reached at day 5 after wounding.
In the rat, both studies used nondiabetic and diabetic animals, which served as an experimental model of type 1 diabetes. In one study, a single square wound was made on the dorsum, and the level of NGF in wounds of diabetic animals was maximal on day 1 and levels were lower than for wounds of nondiabetic animals on days 1, 4, and 7 after wounding. A higher level of NGF in wounds of diabetic animals than for nondiabetic animals occurred on day 15 after wounding. In the other study, two square wounds were made on the dorsum and covered with an occlusive dressing. FGF-1 content in wounds of diabetic animals was significantly lower than in wounds of nondiabetic animals on day 7 after wounding, but was higher than in nondiabetic wounds on day 30 after wounding. FGF-2 content in wounds of diabetic animals was significantly lower than in wounds of nondiabetic animals on days 7 and 15 after wounding. Active TGF-β content in wounds of diabetic animals was significantly lower than in wounds of nondiabetic animals on days 7 and 30 after wounding.
Subcutaneous PVA sponge and stainless steel mesh chamber wound model (Table 4)
In the rat, one study used nondiabetic animals only, whereas the two other studies used both nondiabetic and diabetic animals which served as an experimental model of type 1 diabetes. In the two latter studies the levels of IGF-1 mRNA expression in sponges and of IGF-1 and TGF-β in wound fluid of diabetic rats were significantly reduced compared to nondiabetic animals at 7 days after wounding.
Influence of added growth factors in vivo
Burn ulcer wound model (Table 5)
Application of FGF-1 to cover the wound surface of burn ulcer in diabetic rats, which served as an experimental model of type 1 diabetes, on days 3 and 7 after establishment of ulcer, caused an increase in expression amounts of TGF-β compared with saline-treated diabetic rats on day 7, and a decrease in amounts on day 14.
Excisional skin wound model (Table 5)
Intradermal injection of a plasmid encoding for hypoxia-inducible factor (HIF)-1α increased the expression of VEGF, PlGF, and PDGF in wounds of diabetic mice used as an experimental model of type 2 diabetes. Treatment of wounds in diabetic mice as an experimental model of type 1 diabetes with a vector encoding for PlGF increased the expression of VEGF, FGF-2, and PDGF. Treatment with NGF of wounds in diabetic mice as an experimental model of type 1 diabetes increased the expression of VEGF-A mRNA. Recombinant VEGF applied to wounds of diabetic mice as an experimental model of type 2 diabetes increased the expression of FGF-2 and PDGF.
Treatment with platelet releasate containing PDGF and TGF-β of wounds in diabetic rats as an experimental model of type 1 diabetes decreased FGF-1 content on days 15 and 30 after wounding, but increased FGF-2 content on days 3, 7, 15, and 30 and active TGF-β content on day 7 after wounding.
In vitro studies
Influence of added growth factors
The addition of 10% fetal bovine serum to quiescent human keratinocytes caused a large induction of VEGF mRNA expression within 90 min, and elevated levels were still observed 8 h after serum addition. Moreover, the addition of serum growth factors such as EGF and KGF caused a large increase in VEGF mRNA expression that was maintained for at least 8 h after addition of the growth factor. TGF-β1 also stimulated VEGF mRNA expression, but expression levels had significantly declined by 5 h after addition of the growth factor. The cytokine TNF-α caused a large increase in VEGF mRNA expression within 2 h, whereas IL-1β was much less efficient in increasing VEGF mRNA expression, and similar to TGF-β1, expression levels had significantly declined after 5 h (Table 5).
Discussion
Wound healing and metabolic effects of growth factors
Growth factors mediate the cellular interactions among a variety of cells that orchestrate the complex sequence of cell migration, proliferation, differentiation, and protein expression during wound healing. Growth factors are released at the wound site and considered to be necessary for wound healing. The families of growth factors expressed in varying levels by the cells involved with healing are EGF, KGF, FGF, VEGF, IGF, TGF, PDGF, ILs, and colony-stimulating factor (CSF). Many of these factors are responsible for metabolic responses that have a high influence on wound healing. For example, IGF-1 has a similar effect as insulin; both increase glucose uptake and oxidation, produce similar suppression of glucose production, and free fatty acid levels and fat oxidation rates. An absolute IGF-1 and relative insulin deficiency may contribute to adverse metabolic changes in middle age, 36 and may predispose to developing diabetes. IGF-1 has been used experimentally to treat both type 1 and type 2 diabetes. 37 EGF inhibits gonadal steroid production from both ovarian and testicular tissues, and granulosa cell EGF inhibits follicle-stimulating hormone (FSH) and cAMP stimulation of estrogen and progesterone production. 38 –40 Estrogen is a major regulator of wound repair. 41 EGF and FGF inhibit FSH-induction of luteinizing hormone (LH) receptor in granulosa cells, whereas PDGF stimulates FSH-induction of LH receptor. 42
Impaired healing of diabetic wounds
The impaired healing of diabetic wounds is characterized by delayed re-epithelization, decreased cellular infiltration and granulation tissue formation, reduced angiogenesis, and reduced and disorganized collagen formation. 1,43,44 Increased levels of free radicals in diabetes cause lipid peroxidation and also compromise the metabolism of keratinocytes, fibroblasts, and endothelial cells. 45 In addition, deficiencies have been reported in the levels of various growth factors in the wounds of diabetic animals. These could be caused by altered gene expression of the growth factors, degradation by proteinases in the wounds, and/or reduced or delayed cellular infiltration into the wounds.
Experimental models and growth factors in diabetic wounds
A variety of in vivo animal models was used in the studies, with a greater number of studies performed in mice than in rats. Burn ulcer, incisional, and excisional cutaneous wounds were studied, and experimental models of type 1 and type 2 diabetes were used.
This review has highlighted significant alterations in the temporal pattern and level of gene expression of growth factors in cutaneous wounds of diabetic mice and rats. Growth factors having a decreased mRNA expression pattern and level in the early phases of healing for diabetic wounds compared with nondiabetic wounds were VEGF, PlGF, KGF, FGF-1, FGF-2, IGF-1, IGF-2, TGF-β, and NGF. There was a delay in the expression of mRNAs for KGF, IGF-1, and IGF-2, whereas the expression of FGF-1 and FGF-2 occurred much earlier, in excisional wounds of diabetic mice compared with nondiabetic mice. In addition to the levels of mRNA expression for these growth factors being reduced, the levels of these proteins were decreased in the diabetic wounds compared with the nondiabetic wounds. VEGF and PlGF belong to the family of vascular endothelial growth factors and are responsible for vascularization of the healing wound. NGF is a neurotrophin that is proangiogenic and has been shown to increase the expression of VEGF by endothelial cells. 27 KGF (FGF-7), FGF-1 (acidic FGF), and FGF-2 (basic FGF) are members of the fibroblast-stimulating family of growth factors. IGF-1 and IGF-2 belong to a family of proteins involved in mediating the growth and proliferation of cells. IGF-1 has the potential to stimulate cellular proliferation and collagen production by fibroblasts. TGF-β is a protein that controls proliferation and cellular differentiation in most cells.
Possible combined laser photoirradiation and growth factor administration to treat diabetic wounds
The historical supernatant transfer experiments of Rajaratnam and coworkers demonstrated for the first time the secretion of growth factors by laser-irradiated cells. 10 Photoirradiation of diabetic wounds using laser light at visible red wavelengths stimulated wound healing, 11 and could potentially be combined with growth factor administration to maximize the healing of diabetic wounds. A recent review of the effects of laser photoirradiation on gene expression and release of growth factors by cells in culture indicated that growth factors such as FGF-2, IGF, NGF, and VEGF are increased by exposure to visible red laser light. 9 Possible growth factors that might be considered as part of a combined therapy with laser irradiation could be TGF-β, FGF-1, PlGF, and KGF, as these have not been shown to be increased by laser light treatment. To evaluate which of these might be most suitable, the searched studies were analyzed for the effects of these growth factors on healing of diabetic wounds and the levels of expression of growth factor mRNAs in diabetic wounds and cells in vitro. The latter are summarized in Table 5.
Influence of individual growth factors on healing of diabetic wounds
When applied individually, each of the growth factors TGF-β, FGF-1, PlGF, and KGF enhanced diabetic wound healing. A single intradermal injection of TGF-β1 plasmid DNA immediately after making an excisional wound in diabetic mice was found to stimulate wound closure and the formation of a denser and more organized extracellular matrix. 46 Treatment of skin ulcer wounds in diabetic rats with FGF-1 on days 3 and 7 accelerated wound closure and increased granulation tissue and collagen formation, neoangiogenesis, and re-epithelization. The expression amounts of TGF-β in diabetic rats on days 7 and 14 were modified such that they were no longer different from nondiabetic rats. 13 Application of PlGF as a vector 1 day after creating an excisional wound in diabetic mice stimulated wound closure, with increased formation and accelerated maturation of granulation tissue. The expression of VEGF, FGF-2, and PDGF was increased. In addition, the number of monocytes/macrophages in the granulation tissue was increased. 23 It has been found that the monocyte/macrophage density was significantly reduced in the granulation tissue of diabetic mice compared with nondiabetic mice. 23 A single intradermal injection of KGF-1 DNA plasmid after making an excisional wound in diabetic mice, followed by electroporation, accelerated wound closure, and improved the quality of healing. 47 In vitro studies with human keratinocytes showed that expression of VEGF was increased by KGF. 31
TGF-β is produced by neutrophils and macrophages that migrate into a provisional matrix at the site of the wound, and dermal fibroblasts infiltrating from the wound edge are stimulated by TGF-β and responsible for the synthesis of a rich collagen matrix to replace the provisional matrix. 46 FGF-1 is secreted by multiple cell types and stimulates the proliferation of cells of mesodermal, ectodermal, and endodermal origin. PlGF is expressed by migrating keratinocytes and endothelial cells of small blood vessels during the angiogenic phase of wound healing, and contributes to regulating endothelial and lymphatic vessel development. 48 KGF is produced by mesenchymal cells including fibroblasts and endothelial and smooth muscle cells, and stimulates re-epithelization of wounds. 49
Future studies of combination treatment with laser light and growth factors
Quantitative assessment of wound healing parameters allows the efficacy of various growth factors and cytokines in affecting distinct stages of the wound healing process to be screened. Even though specific growth factors can be an important treatment at certain stages of wound healing, they are insufficient to promote the entire longitudinal process of wound repair. 1 Combination treatments may therefore be the preferred approach to developing future therapies for wound healing. Combinations of two growth factors have been tested and shown to act synergistically to promote wound healing in diabetic animals. They include the combinations of FGF-2 with TGF-β, PDGF with TGF-α, PDGF with IGF-2, and FGF-2 with IGF-2. 50 –52 Since laser irradiation has been shown to positively influence various stages of the wound healing process, research should be implemented on testing a combination therapy of visible red laser irradiation and growth factor administration. The most suitable growth factor would be determined by trial, but in the first instance, TGF-β, FGF-1, PlGF, and KGF could be tested, as these had not been shown to be increased by exposure of cells in culture to visible red laser light. 9 Visible red laser irradiation with VEGF application might also be tested, as administration of VEGF protein increased angiogenesis and granulation tissue formation in diabetic mouse wounds. 35 Combination therapy may enable several critical healing stages to be promoted concomitantly, and reduce the effective dose of laser irradiation and growth factor administered, thereby minimizing adverse changes.
Conclusions
The reviewed studies showed that gene expression and content of certain growth factors are decreased in wounds of diabetic mice and rats. Administration of individual growth factors can enhance wound healing in diabetic animals. Possible combination therapy utilizing laser irradiation and application of specific growth factors at appropriate stages of wound healing may provide an effective means of accelerating the healing of diabetic wounds. Future studies should be designed to test the effect of laser irradiation alone and in combination with a single growth factor in an appropriate animal model of compromised wound healing. One such model is to use genetic diabetic mice in which a full-thickness skin excisional wound is splinted using an occlusive adhesive dressing to retard contraction, and for which healing occurs by re-epithelization and granulation tissue formation. 11
Footnotes
Author Disclosure Statement
No competing financial interests exist.